
Abstract
Numerous clinical and bioavailability studies addressed epigallocatechin gallate (EGCG) beneficial effects; however, our previous work revealed EGCG-induced nephrotoxicity in the presence of diabetes. In this study, the potential myocardial toxicity of EGCG preparation (100 mg/kg/day, IP; 4 days) in diabetic mice injected with streptozotocin (STZ; 150 mg/kg, IP) was investigated. Diabetic mice receiving EGCG preparation showed electrocardiographic changes in addition to elevation of both serum creatine kinase-MB and troponin-I levels accompanied by microscopic myocardial damage. Additionally, myocardial NADPH oxidase, lipid peroxides and nitrotyrosine were increased in the vicinity of decreases of nuclear factor erythroid 2-related factor 2, hemeoxygenase-1, reduced glutathione, total antioxidant capacity, glutathione peroxidase and reductase and heat shock protein 90. Moreover, in diabetic mice, EGCG preparation increased myocardial nuclear factor-kappa B and tumor necrosis factor-alpha in addition to pronounced overexpression of inducible nitric oxide synthase and active caspase-3. Therefore, this study substantiates that EGCG-mediated deterioration compromises diabetes-induced cardiotoxicity to solidify our previous report for its potential nephrotoxicity in the same experimental setting.
Similar content being viewed by others

Introduction
Diabetes mellitus presents from a constellation of heterogeneous malfunctions witnessed by episodes of hyperglycemia and glucose intolerance resulting from lack and/or defective insulin or insulin action. This type of metabolic syndrome arises from derangements in the regulatory systems for storage and mobilization of metabolic fuels1. Of note, diabetics have a 2 to 4 fold higher risk for cardiovascular events, where around 80% of diabetes-associated deaths are linked to cardiovascular disease (CVD)2. As such, diabetes is regarded as a coronary heart disease risk equivalent, i.e.; with a similar risk as an individual with a previous CVD event3. Hyperglycemia is known to aggravate intracellular reactive oxygen species (ROS) formation which induces inflammation and eventually apoptosis4. Most of the potential health benefits associated with green tea consumption have been linked to epigallocatechin-3-gallate (EGCG), a major catechin found in green tea5. EGCG exhibits anti-oxidant, anti-inflammatory and anti-apoptotic potentials6,7,8. Cardioprotective effects of EGCG were also reported in previous experimental studies of hypercholesterolemia and doxorubicin-induced injury9,10. On the other hand, several reports linked the consumption of EGCG containing supplements to acute hepatitis, and hence hepatotoxicity11,12,13. Drug-induced oxidative stress is implicated as a mechanism of toxicity in numerous tissues and organ systems, including liver, kidney, ear, and cardiovascular and nervous systems14. After being absorbed and passed onto the systemic circulation, EGCG undergoes oxidation in liver either enzymatically or non-ezymatically (e.g. oxidation from ROS leaked from the mitochondria) resulting in the formation of a reactive intermediate generating more ROS which react with cellular macromolecules resulting in hepatotoxicity15.
Experimentally, apart from its liver injurious effect when administered orally, high doses of EGCG were found to induce renal and intestinal damage when given in diet15,16. Furthermore, it was reported that natural products may act differently in health compromised affected subjects17. Noteworthy, in the presence of predisposing conditions, such as colitis, fever and diabetes, EGCG initiated paradoxical effects that were not obvious in healthy animals16,18,19. In fact, the administration of EGCG in the presence of lipopolysaccharide-induced fever or dyslipidemia in mice resulted in hepatotoxicity, while in colitic mice it induced nephrotoxicity16,18. These effects are linked to the pro-oxidant nature of EGCG which is attributed to its catechol structural moiety which has low half peak oxidation potential where high concentration of EGCG can undergo self-oxidization and function as prooxidant by producing superoxide anion, hydroxyl radicals, hydrogen peroxide, and quinonoid intermediates causing cytotoxicity20,21.
Based on the data of our previous work signifying nephrotoxic effect of EGCG preparation in mice mediated by oxidative stress, inflammation and apoptosis, the current study was undertaken to delineate the potential cardiotoxic effect of Teavigo® consisting of 94% EGCG in diabetic mice.
Materials and Methods
Animals
Male Swiss albino mice (25–30 g; Faculty of Pharmacy, Cairo University, Cairo, Egypt) were housed under controlled temperature (25 ± 2 °C) and 12 h light/dark cycles with free access to a standard rodent chow diet and water ad libitum. The study was approved by the Ethics Committee for Animal Experimentation (Faculty of Pharmacy, Cairo University; PT 1271) and complies with the Guide for Care and Use of Laboratory Animals (USA NIH, 2011).
Chemicals
The highly purified extract (94% EGCG) Teavigo® (DSM Nutritional Products, Kaiseraugst, Switzerland) and streptozotocin (Sigma–Aldrich, MO, USA) were used in this study.
Experimental design
Mice were randomly divided into 4 groups (n = 16–30 mice each), where saline was injected intraperitoneally in mice to serve as the normal control group. A single dose of STZ (150 mg/kg, IP) was injected in animals to serve as the diabetic control group22. In addition, EGCG preparation (100 mg/kg, IP; 4 days) was administered in normal or diabetic animals representing either EGCG or STZ + EGCG groups, respectively19. The used dose of EGCG preparation was selected based on a pilot study where different doses of EGCG preparation (50, 100, 150 and 200 mg/kg IP) were investigated to detect the highest safe dose in normal animals which caused no myocardial injury based on measurement of serum troponin-I level. Only EGCG150 showed significant high troponin-I level which indicates its cardiotoxic potential whereas EGCG 200 killed all injected animals which confirms its lethality. Accordingly, EGCG 100 was the highest dose which showed no myocardial damage and hence was selected to be used in the main study. Notably, EGCG preparation was injected 48 h after diabetes induction, where mice with blood glucose level ≥200 mg/dl (One Touch Select Glucometer, Life Scan, Pennsylvania,USA) were selected.
Twenty four hours after the last dose of EGCG administration, animals were weighed. Mice were then anesthetized with thiopental (60 mg/kg, i.p.) and kept warmed with a heating lamp to prevent the incidence of hypothermia. Subcutaneous peripheral limb electrodes were inserted for electrocardiographic recording (HPM 7100, Fukuda Denshi, Tokyo, Japan) to determine heart rate (HR) and corrected QT interval (QTc). At the end of experiment, blood was collected under anesthesia and sera were separated for the estimation of creatine kinase-MB (CK-MB)and troponin-I levels. Animals were then euthanized and hearts were rapidly excised, washed with ice-cold saline, dried and weighed. In the present study, 2 equal sets of experiments were conducted; one was used for weight assessment and ECG recording and the heart was homogenized in saline to prepare 5% homogenate for estimation of oxidative stress and inflammatory markers. The other set was used for blood sampling, western blot analysis and histopathological examination.
Biochemical examination
Serum creatine kinase-MB
Serum CK-MB was assessed using a commercially available kit (Stanbio, USA). Results were estimated kinetically at 340 nm using spectrophotometer (Thermo electron corporation, England) and expressed as U/l.
Serum troponin-I
Serum troponin was estimated using ELISA kit (Kamiya Biomedical Company, Washington, USA). The procedure of the used kit was performed according to the manufacturer’s instructions and the results were expressed as ng/ml.
Oxidative stress and inflammatory markers
ELISA kits were used for the assessments of heart NADPH oxidase and nuclear factor erythroid 2-related factor 2 (Nrf2; Cusabio, Wuhan, China), hemeoxygenase-1(HO-1; Enzo Life Sciences, New York, USA), heat shock protein 90 (HSP 90) and nuclear factor-kappa B (NF-κB; Eiaab,Wuhan, China), tumor necrosis factor-alpha (TNF-α; Raybio, Georgia, USA) and nitrotyrosine (MyBioSource, San Diego, USA). The procedure of the used kits was performed according to the manufacturer’s instructions and the results were expressed as ng/mg protein for NADPH oxidase, Nrf2, HSP90 and TNF-α and as μg/mg protein for HO-1 and NF-κB. Results of nitrotyrosine was also expressed as pmol/mg protein.
Moreover, reduced glutathione (GSH), glutathione peroxidase (GPx), glutathione reductase (GR) and thiobarbituric acid reactive substances (TBARS) were assessed using Biodiagnostic kits (Egypt). Procedures were performed according to manufacturer’s instructions and results were expressed as mg/g wet tissue for GSH, nmol/mg protein for TBARS and U/mg protein for GPx and GR. Total antioxidant capacity (TAC) was measured colorimetrically using Zen-Bio kit (North Carolina, USA) and results were expressed as mmol/mg protein. The protein content in tissue homogenates was determined according to the method of Lowry et al.23.
Western blot analysis of inducible nitric oxide synthase
Part of heart was homogenized in lysis buffer and quantified for protein levels using a Bicinchoninic acid protein assay (BCA) kit (Thermo Fisher Scientific Inc., USA). Protein expression was assessed as previously described24 using anti-iNOS antibodies (Stressgen Biotechnologies, Victoria, British Columbia, Canada). The amount of protein was quantified by densitometric analysis of the autoradiograms using a scanning laser densitometer (Biomed Instrument Inc., USA). Results were expressed as arbitrary units after normalization for beta-actin (β-actin) protein expression.
Histopathological examination
At the end of the experimental period, the apex of heart was isolated, rinsed in ice-cold saline and immediately fixed in 10% formalin. The specimens were processed for paraffin embedding, and 5μm sections were prepared and then stained with hematoxylin and eosin (H&E; x400).
Active caspase-3 immunohistochemical examination
Immunohistochemical examination of active caspase-3 was carried out on 5 μm thick deparaffinized sections incubated in a 0.01 M sodium citrate solution (pH 6) at 120 °C for 10 min, followed by a 2 h cool-down. The sections were then treated with the proteolytic enzyme proteinase K (Dako, Copenhagen, Denmark), washed in phosphate buffered saline (PBS) for 5 min and incubated with a primary antibody against caspase-3 (Novus Biologicals, Colorado, USA) for 60 min at 37 °C. After washing in PBS, a secondary antibody (Novus Biologicals, Colorado, USA) was applied for 60 min, visualized with 3,3′-diaminobenzidine (DAB) chromagen (Dako, Copenhagen, Denmark) and counterstained with hematoxylin (x400).
Statistical analysis
All data obtained were presented as mean ± SEM. Results were analyzed using one-way analysis of variance (ANOVA) followed by Tukey-Kramer multiple comparisons test. Statistical analysis was performed using GraphPad Prism© software (version 6.01; Graph Pad Software, California, USA). For all the statistical tests, the level of significance was fixed at P < 0.05.
Results
EGCG induced changes in heart to body weight ratio as well as perturbations in electrocardiographic measurements, myocardial biomarkers and histological examinations in diabetic mice
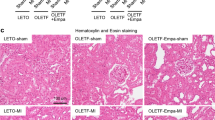
Myocardial alterations were assessed by measurements of heart to body weight ratio as well as HR and QTc. Serum CK-MB and troponin-I levels were also estimated as markers of myocardial damage. EGCG group showed no alteration in these parameters compared to normal group. STZ increased heart to body weight ratio and QTc whereas it decreased HR. Moreover, this group showed elevation of serum CK-MB and troponin-I levels. On the other hand, administration of EGCG to diabetic mice markedly deteriorated these parameters compared to diabetic group confirming that EGCG preparation potentiated diabetes-induced myocardial damage (Table 1). Moreover, EGCG administration to normal mice showed no histopathological alteration compared to the normal control mice. However, diabetic mice showed moderate congestion of myocardial blood vessels whereas EGCG administration to diabetic mice caused severe congestion of myocardial blood vessels and moderate intermuscular edema verifying that EGCG preparation potentiated diabetes-induced myocardial damage (Fig. 1).

Effect of EGCG on STZ- induced changes in histological examination (H&E x400). (A) Heart of a normal mouse showing normal cardiomyocytes. (B) Heart of a mouse from EGCG group showing normal cardiomyocytes. (C) Heart of a diabetic mouse showing moderate congestion of myocardial blood vessels. (D) Heart of a diabetic mouse receiving EGCG showing severe congestion of myocardial blood vessels. (E) Heart of a diabetic mouse receiving EGCG showing moderate intermuscular edema.
EGCG exaggerated oxidative stress in diabetic mice
In normal mice, EGCG significantly decreased NADPH oxidase, TBARS and nitrotyrosine contents with increases in TAC, GSH, GPx, Nrf2, HO-1, and HSP 90 contents indicating restrained ROS generation. On the other hand, diabetic mice receiving EGCG have shown increased NADPH oxidase, TBARS and nitrotyrosine associated by decreases in TAC, GSH, GPx, GR, Nrf2, HO-1 and HSP 90 cardiac contents. These effects were more profound than those in STZ animals, signifying the worsening of the oxidative stress status with a shifted balance towards a pro-oxidative milieu by STZ and EGCG administration (Figs 2 and 3).

Effect of EGCG on STZ-induced changes in (A) NADPH oxidase, (B) TBARS, (C) TAC, (D) GSH, (E) GPx and (F) GR. Each value represents the mean of 6–8 experiments ± S.E.M. *P < 0.05 vs. normal, @P < 0.05 vs. EGCG, #P < 0.05 vs. STZ.

Effect of EGCG on STZ-induced changes in (A) Nrf2, (B) HO-1 and (C) HSP 90. Each value represents the mean of 6–8 experiments ± S.E.M. *P < 0.05 vs. normal, @P < 0.05 vs. EGCG, #P < 0.05 vs. STZ.
EGCG augmented inflammation in diabetic mice
EGCG administered to normal mice did not affect NF-κB and TNF-α as well as iNOS protein expression compared to the normal control group. Induction of diabetes increased the myocardial NF-κB and TNF-α contents as well as iNOS protein expression, while administration of EGCG preparation to diabetic animals showed marked increment in the previously mentioned markers indicating a severe inflammatory status (Fig. 4).

Effect of EGCG on STZ-induced changes in (A) iNOS protein expressionas well as (b) nitrotyrosine, (C) NF-κB and (B) TNF-α contents. Each value represents the mean of 6–8 experiments ± S.E.M. *P < 0.05 vs. normal, @P < 0.05 vs. EGCG, #P < 0.05 vs. STZ.
EGCG induced apoptosis in diabetic mice
Normal mice treated with EGCG preparation showed no alteration from the normal control animals regarding active caspase-3 expression, whereas diabetic mice receiving EGCG preparation showed significant elevation of active caspase-3 expression compared to diabetic control mice. This signifies EGCG prominent apoptotic potential in the presence of diabetes (Fig. 5).

Effect of EGCG on STZ-induced changes in immunohistochemical expression of active caspase-3 expression (400). (A) Normal animals receiving saline showed normal myocardium with −ve immunostaining for active caspase-3, (B) normal animals receiving EGCG showed normal myocardium with −ve immunostaining for active caspase-3, (C) diabetic animals showed focal brown immunostaining for active caspase-3 in myocardial tissues and (D) diabetic animals receiving EGCG showed widely distributed brown immunostaining for active caspase-3 in myocardial tissues. Each value represents the mean of 3 experiments ± S.E.M. *P < 0.05 vs. normal, @P < 0.05 vs. EGCG, #P < 0.05 vs. STZ.
Discussion
In an earlier study, we documented the involvement of inflammation, oxidative stress and apoptosis in EGCG-induced nephrotoxicity in the presence of diabetes19. To the best of the authors’ knowledge, the current study is the first to provide evidence for the potential cardiotoxic effect of EGCG preparation in diabetic mice. Mouse maybe a better predictor of EGCG toxicity than rat due to higher bioavailability in both human and mouse compared to rat25,26. Moreover, i.p route of administration offers a higher bioavailability compared to oral route (poor bioavailability and extensive biotransformation) which allows better investigation of the potential biological activity and toxicity of EGCG as estimated using previous studies27,28. Moreover, higher EGCG bioavailability has been linked to higher unconjugated EGCG level which is linked to its related toxicity29.
Myocardial hemodynamic and biochemical alterations were clearly observable in the EGCG treated and non treated diabetic groups, which were lower with diabetes alone. Moreover, the microscopic appearances, featuring severe myocardial blood vessels congestion and moderate intermuscular edema in diabetic mice receiving EGCG outweighed the effect of diabetes alone.
The increase in heart to body weight ratio in EGCG treated and non treated diabetic mice indicated hypertrophy of myocardial tissues. Hypertrophy may contribute to impairment of electromotive forces30 as demonstrated by decrease in HR and increase in QTc in the present study. HR variability could be also related to metabolic alterations due to hyperglycemia and insulinopenia or downregulation of cardiac adrenergic receptors31. Short-term diabetes mellitus induced by STZ has previously demonstrated to modify autonomic control of HR and cause bradycardia which was reversible by insulin administration confirming that these cardiovascular changes were related to metabolic perturbations and not to the direct toxic effect of diabetogenic agent; STZ32.
Serum CK-MB is a useful early diagnostic index for myocardial injury and infarction, while troponin-I is a specific and more sensitive cardiac injury biomarker that predicts the risk of both cardiac cell death and subsequent infarction33. Therefore, probable reason for the rise in serum levels of CK-MB and troponin-I by EGCG could be cell death, namely apoptosis as will be elaborated later that may impose diabetic mice to the possible risk for myocardial injury. Noteworthy, CK-MB and troponin-I elevations are linked to increased cell permeability caused by inflammation and free radical damage34,35. Such effects can be clarified by the core findings of the present work as EGCG preparation, in diabetic mice, activated NFκ-B with the association of TNF-α and NADPH oxidase elevation as well as iNOS overexpression that surpassed diabetic animals. Of note, NADPH oxidase is a major source of ROS36 which cause oxidative damage, justified in the present study by the marked increase in oxidative stress markers (TBARS and nitrotyrosine) in addition to marked depletion of endogenous anti-oxidants namely, GSH, GPx, GRx and TAC. The aforementioned events support EGCG deleterious effects in diabetic kidney19 and reinforce its reported pro-oxidant effect which was previously linked to its nephro- and hepatotoxicity16,37. The elevation of inflammatory cytokines enhances the expression of iNOS contributing to the production of the highly reactive oxidant peroxynitrite in the presence of oxidative stress and resulting in reduced nitric oxide (NO) level and endothelial dysfunction38. Of note, the pro-inflammatory cytokine TNF-α35 and free radicals39 were documented to increase the transcriptional activity of NFκ-B, thus contributing to an endless feed forward cycle that propagates and potentiates both inflammation and oxidative stress leading hence to excessive CK-MB and troponin-I leakage as documented herein in mice receiving EGCG preparation and STZ.
Apart from the involvement of NF-κB in oxidative and inflammatory damage, the current more conspicuous reduced level of HO-1 in EGCG-treated diabetic animals plays also a role in the catechin-mediated myocardial toxicity. This enzyme catalyzes the degradation of heme which has both pro-oxidant and pro-inflammatory properties; hence in the vicinity of a sharp decrease of HO-1, free radicals and inflammation that lead to cellular injury are augmented40. The decrease in HO-1 might be clarified by the present inhibition of Nrf2 in EGCG-treated diabetic mice that was again more prominent than diabetic mice. These effects go in line with our previous report of its nephrotoxic effect in the presence of diabetes19. Moreover, it has been earlier reported that high doses of EGCG reduced the expression of HO-120,41. The transcription factor Nrf2 is an important machinery that upregulates the HO-1 gene. This enzyme initiates antioxidant and anti-inflammatory processes, hence Nrf2 down-regulation triggers and progresses various diabetic complications42,43. Moreover,it was reported that STZ-induced diabetes in Nrf2 knockout mice rapidly progressed to severe myocardial lesions associated by robust inflammatory response, oxidative stress, and apoptosis44. These data indicate that Nrf2 and its downstream target HO-1 serve as defense factors against cardiac diabetic complications; hence their excessive reduction by EGCG preparation in diabetic animals seen in this study can explain, in part, the polyphenol cardiotoxic effect. Besides the upregulation of HO-1, Nrf2 initiates the transcription of a number of anti-oxidative genes to reduce the pathological oxidative stress in the heart43. Such an effect might afford an additional explanation to the previously documented descent of TAC and GSH in addition to antioxidants enzymes controlling GSH levels (GPx and GR) by EGCG + STZ in the present work.
In response to various forms of stress, cells activate a highly conserved heat shock response in which a set of HSPs are induced to participate in cellular repair and protective mechanisms45,46. Several studies noticed that diabetes is associated with significant reduction of HSPs levels47,48,49. Moreover, in diabetic patients, cells are more vulnerable to damage if HSPs levels are decreased which results in organ failure50. Since in the present study EGCG preparation was shown to suppress HSP 90 in diabetic mice more persistently than diabetes alone, therefore, EGCG-induced cardiotoxicity could be attributed to its ability to decrease HSP 90, and hence maladapted stress defense responses. The present decrease in HSP 90 after EGCG administration in diabetic mice confirms our previous report in the kidney19 and is consistent with the previous finding which elucidated that this proteo-stress marker was markedly down-regulated in mice receiving high doses of green tea polyphenols in diet16. Furthermore, in response to excessive oxidative stress-induced by high doses of EGCG, HSP90 was downregulated in liver and kidney20.
Diabetic control animals have shown low incidence of apoptosis; however, active caspase-3 was prominently elevated in the STZ/EGCG group, solidifying its apoptotic potential in the diabetic kidney19. It has been anticipated that oxidative stress and the pro-inflammatory cytokine TNF-α could contribute to diabetic cardiomyopathy through the stimulation of the intrinsic and extrinsic pathways of programmed cell death, respectively51. Additionally, Kim et al.52. suggested that the decreased HSP 90 expression in type 2 diabetic kidneys may mediate apoptosis; hence affording an additional clarification for increased apoptosis associated by EGCG administration in STZ mice. Accordingly, in the present study, the excessive cardiac toxicity produced by the polyphenol under hyperglycemic conditions could been linked to the present exaggerated oxidative stress and inflammation responses, as well as the reduction of HSP 90 in the heart to jeopardize cardiomyocytes function leading to CK-MB and troponin-I elevation as previously anticipated.
Therefore, the current investigation elaborated that parentral application of EGCG preparation (Teavigo®) exhibited cardiotoxicity in the presence of diabetes although no toxicity was observed in normal animals receiving the same dose and route of administration. This deleterious effect could be attributed to EGCG-induced apoptosis triggered by oxidative stress, inflammation and HSP 90 suppression which eventually leads to cardiac damage evidenced by elevation of myocardial biomarkers.
Conclusion
In spite of its well-known favorable effects, EGCG administration to diabetic mice revealed increased cardiotoxicity. Considering the rise of global diabetes, EGCG administration may impact cardiovascular health accelerating the progression of the disease-associated complications. Mediation of oxidative stress, inflammation, and apoptosis in the cardiomyopathy-induced by EGCG preparation in diabetes can be considered especially for parentral application of EGCG which exhibits higher bioavailability and thus its related toxicity. Accordingly, diabetic patients should consume EGCG based supplements cautiously until their cardiotoxic effects in the presence of diabetes are clinically evaluated.
Limitations and recommendations for future research
Despite being 94% of teavigo content, further studies are required to investigate the toxicity of pure EGCG in presence of uncontrolled hyperglycemic conditions and complications of chronic untreated diabetes.
References
Piero, M. N., Nzaro, G. M. & Njagi, J. M. Diabetes mellitus-a devastating metabolic disorder. Asian Journal of Biomedical and Pharmaceutical Sciences 4(40), 1–7 (2015).
Winer, N. & Sowers, J. R. Epidemiology of diabetes. The Journal of Clinical Pharmacology 44(4), 397–405 (2004).
Whiteley, L., Padmanabhan, S., Hole, D. & Isles, C. Should diabetes be considered a coronary heart disease risk equivalent? Results from 25 years of follow-up in the Renfrew and Paisley survey. Diabetes Care 28(7), 1588–1593 (2005).
Peng, C. et al. High glucose induced oxidative stress and apoptosis in cardiac microvascular endothelial cells are regulated by FoxO3a. PLoS One 8(11), e79739 (2013).
Saito, K., Mori, S., Date, F. & Ono, M. Epigallocatechin gallate inhibits oxidative stress-induced DNA damage and apoptosis in MRL-Faslpr mice with autoimmune sialadenitis via upregulation of heme oxygenase-1 and Bcl-2. Autoimmunity 47(1), 13–22 (2014).
Donà, M. et al. Neutrophil restraint by green tea: inhibition of inflammation, associated angiogenesis, and pulmonary fibrosis. The Journal of Immunology 170(8), 4335–4341 (2003).
Mereles, D. & Hunstein, W. Epigallocatechin-3-gallate (EGCG) for clinical trials: more pitfalls than promises. International Journal of Molecular Sciences 12(9), 5592–5603 (2011).
Schroeder, E. K. et al. Green tea epigallocatechin 3-gallate accumulates in mitochondria and displays a selective antiapoptotic effect against inducers of mitochondrial oxidative stress in neurons. Antioxidants & Redox Signaling 11(3), 469–480 (2009).
Zhong, W., Huan, X. D., Cao, Q. & Yang, J. Cardioprotective effect of epigallocatechin-3-gallate against myocardial infarction in hypercholesterolemic rats. Experimental and Therapeutic Medicine 9(2), 405–410 (2015).
Khan, G. et al. Cardioprotective effect of green tea extract on doxorubicin-induced cardiotoxicity in rats. Acta Poloniae Pharmaceutica 71(5), 861–868 (2014).
Bonkovsky, H. L. Hepatotoxicity associated with supplements containing Chinese green tea (Camellia sinensis). Annals of Internal Medicine 144, 68–71 (2006).
Garcia-Moran, S., Saez-Royuela, F., Gento, E., Lopez, M. A. & Arias, L. Acute hepatitis associated with Camellia thea and Orthosiphonstamineus ingestion. Gastroenterology& Hepatology 27, 559–560 (2004).
Abu el Wafa, Y., Benavente Fernandez, A., Talavera Fabuel, A., Perez Ramos, M. A. & Ramos-Clemente, J. I. Acute hepatitis induced by Camellia sinensis(green tea). An Med Interna. 22(6), 298 (2005).
Deavall, D. G., Martin, E. A., Horner, J. M. & Roberts, R. Drug-induced oxidative stress and toxicity. Journal of Toxicology 2012, 645460 (2012).
Lambert, M. J., Kennett, S., Sang, K. R., Reuhl, J. J. & Yang, C. S. Hepatotoxicity of high oral dose (-)-epigallocatechin-3-gallate in mice. Food and Chemical Toxicology 48(1), 409–416 (2010).
Inoue, H. et al. High-dose green tea polyphenols induce nephrotoxicity in dextran sulfate sodium-induced colitis mice by down-regulation of antioxidant enzymes and heat-shock protein expressions. Cell Stress Chaperones 16(6), 653–62 (2011).
Risher, J. F., Todd, G. D., Meyer, D. & Zunker, C. L. The elderly as a sensitive population in environmental exposures: making the case. Reviews of Environmental Contamination and Toxicology 207, 95,col (2010).
Saleh, I. G. et al. Effect of green tea and its polyphenols on mouse liver. Fitoterapia 90, 151–159 (2013).
Rasheed, N. O. A., Ahmed, L. A., Abdallah, D. M. & El-Sayeh, B. M. Nephro-toxic effects of intraperitoneally injected EGCG in diabetic mice: involvement of oxidative stress, inflammation and apoptosis. Scientific Reports 7, 40617 (2017).
Murakami, A. Dose-dependent functionality and toxicity of green tea polyphenols in experimental rodents. Archives of Biochemistry and Biophysics 557, 3–10 (2014).
Elbling, L. et al. Green tea extract and (–)-epigallocatechin-3-gallate, the major tea catechin, exert oxidant but lack antioxidant activities. FASEB Journal 19(7), 807–809 (2005).
Aldahmash, B. A., El-Nagar, D. M., Ibrahim, K. E. & Metwaly, M. S. Biotin amelioration of nephrotoxicity in streptozotocin-induced diabetic mice. Saudi Journal of Biological Sciences 22(5), 564–569 (2015).
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193, 265–275 (1951).
Ahmed, L. A., Shehata, N. I., Abdelkader, N. F. & Khattab, M. M. Tempol, a superoxide dismutase mimetic agent, ameliorates cisplatin-induced nephrotoxicity through alleviation of mitochondrial dysfunction in mice. PLoS One 9, e108889 (2014).
Lambert, J. D. et al. Epigallocatechin-3- gallate is absorbed but extensively glucuronidated following oral administration to mice. Journal of Nutrition 133, 4172–4177 (2003).
Lu, H., Meng, X. & Yang, C. S. Enzymology of methylation of tea catechins and inhibition of catechol-Omethyltransferase by (−)-epigallocatechin gallate. Drug Metabolism & Disposition 31, 572–579 (2003).
Ramachandran, B., Jayavelu, S., Murhekar, K. & Rajkumar, T. Repeated dose studies with pure Epigallocatechin-3-gallate demonstrated dose and route dependant hepatotoxicity with associated dyslipidemia. Toxicology Reports 3, 336–345 (2016).
Sang, S. et al. Synthesis and structure identification of thiol conjugates of (−)-epigallocatechin gallate and their urinary levels in mice. Chemical Research in Toxicology 18(11), 1762–1769 (2005).
James, K. D., Forester, S. C. & Lamber, J. D. Dietary pretreatment with green tea polyphenol, (−)-epigallocatechin-3-gallate reduces the bioavailability and hepatotoxicity of subsequent oral bolus doses of (−)-epigallocatechin-3-gallate. Food and Chemical Toxicology 76, 103–108 (2015).
Henkens, I. R. et al. Early changes in rat hearts with developing pulmonary arterial hypertension can be detected with three dimensional electrocardiography. American Journal of Physiology-Heart and Circulatory Physiology 293, 1300–1307 (2007).
Williams, R. S., Schaible, T. F., Scheur, J. & Kennedy, R. Effects of experimental diabetes on adrenergic and cholinergic receptors of rat myocardium. Diabetes 32, 881–886 (1983).
Schaan, B. D. et al. Time course of changes in heart rate and blood pressure variability in streptozotocin-induced diabetic rats treated with insulin. Brazilian Journal of Medical and Biological Research 30(9), 1081–1086 (1997).
Priscilla, D. H. & Prince, P. S. M. Cardioprotective effect of gallic acid on cardiac troponin-T, cardiac marker enzymes, lipid peroxidation products and antioxidants in experimentally induced myocardial infarction in Wistar rats. Chemico-biological interactions 179(2), 118–124 (2009).
Apple, F. S. Tissue specificity of cardiac troponin I, cardiac troponin T and creatine kinase-MB. Clinica Chimica Acta 284(2), 151–159 (1999).
Mariappan, N. et al. NF-κB-induced oxidative stress contributes to mitochondrial and cardiac dysfunction in type II diabetes. Cardiovascular research 85(3), 473–483 (2009).
Li, W. G. et al. H2O2-induced O2 Production by a non-phagocytic NAD(P)H Oxidase Causes Oxidant Injury. Journal of Biological Chemistry 276(31), 29251–29256 (2001).
Galati, G., Lin, A., Sultan, A. M. & O’Brien, P. J. Cellular and in vivo hepatotoxicity caused by green tea phenolic acids and catechins. Free Radical Biology and Medicine 40(4), 570–580 (2006).
Gunnett, C. A., Lund, D. D., McDowell, A. K., Faraci, F. M. & Heistad, D. D. Mechanisms of inducible nitric oxide synthase-mediated vascular dysfunction. Arteriosclerosis, Thrombosis, and Vascular Biology 25(8), 1617–1622 (2005).
Rani, V. & Yadav, U. Free radicals in human health and disease, 1st edition. Springer, New Delhi, India (2014).
Abraham, N. G., Tsenovoy, P. L., McClung, J. & Drummond, G. S. Heme oxygenase: a target gene for anti-diabetic and obesity. Current pharmaceutical design 14(5), 412–421 (2008).
Kim, M., Murakami, A., Miyamoto, S., Tanaka, T. & Ohigashi, H. The modifying effects of green tea polyphenols on acute colitis and inflammation-associated colon carcinogenesis in male ICR mice. Biofactors 36, 43–51 (2010).
Velmurugan, G. V., Sundaresan, N. R., Gupta, M. P. & White, C. Defective Nrf2-dependent redox signalling contributes to microvascular dysfunction in type 2 diabetes. Cardiovascular research 100(1), 143–150 (2013).
Chen, J., Zhang, Z. & Cai, L. Diabetic cardiomyopathy and its prevention bynrf2: current status. Diabetes & Metabolism journal 38(5), 337–345 (2014).
He, X., & Ma, Q. Disruption of Nrf2 synergizes with high glucose to cause heightened myocardial oxidative stress and severe cardiomyopathy in diabetic mice. Journal of diabetes & metabolism Supp7:002 https://doi.org/10.4172/2155-6156.S7-002 (2012).
Morimoto, R. I. Proteotoxic stress and inducible chaperone networks in neurodegenerative disease and aging. Genes & development 22(11), 1427–1438 (2008).
Morimoto, R. I. The heat shock response: systems biology of proteotoxic stress in aging and disease. Cold Spring Harbor symposia on quantitative biology 76, 91–99 (2011).
Kurucz, I. et al. Decreased expression of heat shock protein 72 in skeletal muscle of patients with type 2 diabetes correlates with insulin resistance. Diabetes 51(4), 1102–1109 (2002).
Bruce, C. R., Carey, A. L., Hawley, J. A. & Febbraio, M. A. Intramuscular heat shock Protein 72 and heme oxygenase-1 mRNA are reduced in patients with type 2 diabetes evidence that insulin resistance is associated with a disturbed antioxidant defense mechanism. Diabetes 52(9), 2338–2345 (2003).
Kavanagh, K., Zhang, L. & Wagner, J. D. Tissue-specific regulation and expression of heat shock proteins in type 2 diabetic monkeys. Cell Stress and Chaperones 14(3), 291–299 (2009).
Hooper, P. L. & Hooper, J. J. Loss of defense against stress: diabetes and heat shock proteins. Diabetes technology & therapeutics 7(1), 204–208 (2005).
Zhang, B. et al. Myricitrin alleviates oxidative stress-induced inflammation and apoptosis and protects mice against diabetic cardiomyopathy. Scientific Reports 7, 44239 (2017).
Kim, Y. S. et al. Expression of heat shock protein 90 in the kidneys of diabetic db/db mice. European review for medical and pharmacological sciences 18(15), 2198–2204 (2014).
Acknowledgements
The authors are thankful to Dr. Hebat Allah Amin, surgical and forensic pathologist at Egyptian Forensic Medicine Authority (EFMA) for her efforts in the histological examinations. This research received no specific grant from any funding agency in the public or commercial.
Author information
Authors and Affiliations
Contributions
Statement: B.M.E.L.-Sayeh was responsible for the study design. N.O. AbdeL Rasheed implemented the research. N.O. AbdeL Rasheed, L.A. Ahmed, D.M. Abdallah and B.M. E.L.-Sayeh wrote the main manuscript text and prepared the figures. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rasheed, N.O.A., Ahmed, L.A., Abdallah, D.M. et al. Paradoxical cardiotoxicity of intraperitoneally-injected epigallocatechin gallate preparation in diabetic mice. Sci Rep 8, 7880 (2018). https://doi.org/10.1038/s41598-018-25901-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-25901-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
