
Abstract
Both biodiversity and biomass are important variables in forest ecosystems, and the relationship between them is critical for ecosystem functioning and management. The primary Pinus kesiya forest is increasingly threatened by human disturbance in Yunnan Province. We observed that species richness had a positive impact on aboveground biomass across all forest vegetation layers, and this relationship was strongest in the herb layer. The asymptotic relationship between cumulative species number and aboveground biomass suggested that individual of Pinus kesiya trees with relatively large diameters contributed the majority of the aboveground biomass in the tall tree strata due to their strong competitive advantage over other tree species. Although aboveground biomass increased with stand age in the tall tree strata, climate factors and the soil nutrient regime affected the magnitude of the diversity-productivity relationship. Stand age had no significant effect on species richness and aboveground biomass in the forest understory. The effect of the positive diversity-productivity relationship of the tall trees on the shrub layer was negligible; the diversity-productivity relationship in the forest understory was significantly affected by the tall tree aboveground biomass. The tall trees have increased the strength of the positive diversity-productivity relationship in the forest understory.
Similar content being viewed by others

Introduction
The relationship between biological diversity and ecosystem functioning has stimulated research on the relationship between terrestrial plant species richness and temporal variation in biomass production during the past two decades1,2. A clear forest biodiversity-biomass relationship has been shown in previous studies. Positive species diversity and biomass relationships are ubiquitous in most forest ecosystems around the world, and species loss in these ecosystems negatively impacts ecosystem functioning3,4. Most previous research has focused on the relationship between species diversity and productivity, not biomass, in forest ecosystems5,6. However, biomass is strongly correlated with productivity when the effects of stand age are taken into account4,7,8. An increasing number of studies have focused on the relationship between biodiversity and aboveground biomass, as well as the mechanisms resulting in variations in both species functional traits and environmental conditions in tropical, temperate and boreal forest ecosystems4,9.
Previous studies have shown that tree species in the overstory play important roles in mediating the effects of environmental conditions and disturbance on understory species richness10,11. Accordingly, studies on the relationships between species richness and biomass across forest strata have been carried out in boreal and subtropical forests9,11,12,13,14. Reich et al. confirmed that a positive species richness and aboveground biomass relationship across forest strata was found, and the understory species richness is mediated by the indirect effects of the dominant producers on resource availability and heterogeneity in a boreal forest13. Further evidence from Canada’s National Forest Inventory studies shows that positive relationships between species richness and aboveground biomass in understory vegetation may not affect overstory tree species12. Simultaneously, similar findings have also indicated that diversity-productivity relationships in the forest understory were not associated or were negatively associated with the aboveground biomass of the overstory layer7. In contrast, the species diversity of the overstory layer significantly increased the species diversity of the understory layer in a subtropical forest, while the effect of the aboveground biomass of the overstory on the diversity and aboveground biomass of the understory was negligible14. The occurrence of shade species from the canopy trees in the understory layer is a common phenomenon, and the overstory-understory species richness relationship suggests that different functional groups have specific growth strategies11. There is no doubt that the size of the overstory, the interaction between species richness and aboveground biomass in this layer, and its indirect effects on resource availability and heterogeneity mediate the relationship between species richness and aboveground biomass in the understory layers7,12,13.
The driving mechanisms of comprehensive causal relationships could be explained by both selection and complementarity effects15. A previous study demonstrated that the majority of productivity of the forest ecosystem occurred in the overstory, while understory plants have relatively low contributions to the total biomass stock16. The relationship between species richness and biomass is more complex in natural ecosystems because of species dominance and composition17. The selection effect suggests that species with the most productive traits will have greater opportunities to dominate the biomass of species-rich polycultures through interspecific competition, while the complementarity effect suggests that species can capture resources to maintain diversity as a result of niche partitioning or interspecific interactions when they occur together in plant communities15,18.
Although positive species diversity and aboveground biomass relationships are mostly found across forest strata that exhibit higher species richness and many trophic levels2,7, biotic and abiotic factors affect the magnitude and quality of such relationships, and these effects should be considered, especially by increasing the range of variables included in multivariate studies, such as competition intensity, resource heterogeneity and population dynamics13. Stand age is closely associated with the species richness-biomass relationship9. Simultaneously, the consideration of environmental conditions may be crucial for understanding the relationship between species richness and biomass along with two drivers4: First, higher resource availability supports more biomass accumulation under more favourable climate and soil nutrient conditions15. Second, the diversity-productivity relationship shows a unimodal association from harsh to favourable climate and soil nutrient conditions4. The extents of the influences of climate and local site conditions on plant species diversity differ significantly among vegetation strata19. Climate, soil nutrient conditions and stand age are recognized as directly or indirectly affecting the species richness-aboveground biomass relationship, but were rarely explicitly considered because of the instability in environmental conditions9,12.
Primary Pinus kesiya forests have their largest distribution throughout northern India, the Philippines, Myanmar, Vietnam, Laos, Thailand and Yunnan Province, China20,21. In Yunnan Province, China, primary Pinus kesiya forests play an important role in local forest ecosystem functioning, including carbon sequestration and biological conservation, in addition to timber production and resin tapping. Deforestation has decreased the area of forest over recent decades. Degradation of the remaining stands and climate change have affected the regeneration, growth and distribution of primary Pinus kesiya forests, increasing species loss and threatening stable ecosystem functioning22. Pinus kesiya usually invades monsoon evergreen broadleaf forest areas after the broadleaf species have been cleared23, causing the vertical structure of the community to experience obvious species stratification. There is a distinct difference between a primary Pinus kesiya forest and a boreal or subtropical evergreen broadleaf forest, as that almost all Pinus kesiya individuals are distributed in the overstory layer in the primary Pinus kesiya forest, while most of the broadleaf species occupy the understory layer. There is little information regarding the relationships between the species richness and aboveground biomass across forest vegetation strata in primary Pinus kesiya forests.
In this study, we used a multivariate model to evaluate path hypotheses to assess the relationship between the species richness and aboveground biomass of the tall tree, short tree, shrub, herb and liana layers in a primary Pinus kesiya forest. We also analysed the influences of the tall tree layer, stand age, soil nutrient regime and climate factors as mechanisms driving the relationship between species richness and aboveground biomass across forest vegetation strata based on existing theoretical frameworks7. Based on the existing study of Zhang et al.12 and our field data, we summarized theoretical frameworks to test the following paths: (1) the relationships between species richness and aboveground biomass across forest vegetation strata, including the tall tree, short tree, shrub, liana and herb layers; (2) the influences of the soil nutrient regime, stand age and climate factors on the species richness across forest vegetation strata and the effect of stand age on aboveground biomass; (3) the influences of species richness and aboveground biomass in the tall tree strata on the paths of (1) and (2) in the short tree, shrub, liana and herb layers; and (4) the influence of species richness in the short tree strata on aboveground biomass across all layers. The aim of this study was to disentangle the potential mechanisms that mediate the relationship between species richness and aboveground biomass across forest strata and to provide a reference for the management of natural subtropical coniferous forests under global climate change.
Results
Relationships between species richness and aboveground biomass across all forest vegetation strata
Few species contributed a great deal to the aboveground biomass in the primary Pinus kesiya forest. The cumulative proportion of aboveground biomass showed that there was a strong asymptotic relationship with the cumulative species number (Fig. 1a). One of 169 species (accounting for 83.25% of the species number in all plots) was found to have the contribution of more than 50% of the total aboveground biomass, up to 55.89% for Pinus kesiya, while most tree species contributed little. The majority of Pinus kesiya stems were in the tall tree layer, up to 85.43% (Fig. 1b).

Cumulative aboveground biomass was ranked against cumulative species number and DBH class distribution of Pinus kesiya at the community level. DBH, diameter at breast height; I, DBH < 9 cm; II, 9 ≤ DBH < 20 cm; III, 20 ≤ DBH < 30 cm; IV, 30 ≤ DBH < 40 cm; V, 40 ≤ DBH < 50 cm; VI, 50 ≤ DBH < 60 cm; VII, 60 ≤ DBH < 70 cm; VIII, 70 ≤ DBH < 80 cm.
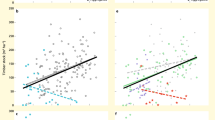
The aboveground biomass values of the tall tree, short tree, shrub, herb and liana layers were positively correlated with the species richness of the tall tree, short tree, shrub, herb and liana layers based on the cubic OLS regression, respectively (Fig. 2a–e). The aboveground biomass linearly increased with the species richness across the forest vegetation strata. The relationship between species richness and aboveground biomass for the tall tree layer was weaker than those for the shrub, herb and liana layers. The aboveground biomass and species richness in the herb layer showed a stronger positive correlation than those in the other forest vegetation strata (Fig. 2d).

Relationships between the tall tree species richness and aboveground biomass, short tree species richness and aboveground biomass, shrub species richness and aboveground biomass, herb species richness and aboveground biomass, and liana species richness and aboveground biomass. The red fitted lines are from the OLS polynomial regression, and the shaded areas show the 95% confidence intervals for the fit.
Effects of species richness, stand age and abiotic factors on aboveground biomass in the tall tree strata
The structural equation model linking the aboveground biomass and species richness of the tall tree layer showed a good fit (SRMR = 0.023, GFI = 0.998), accompanied by variations in the tall tree species richness and stand age of up to 57.3%. Our results suggested that the tall tree species richness and stand age directly affected the aboveground biomass of the tall tree strata (r = 0.096 and r = 0.727, respectively) (Fig. 3). Simultaneously, there were positive relationships among the soil nutrient regime, growing degree days, climate moisture index and tall tree species richness, of which the soil nutrient regime indirectly affected the aboveground biomass of the tall tree layer through the tall tree species richness (r = 0.027) (Fig. 3). Stand age had a stronger effect than species richness on the tall tree aboveground biomass based on the SEM results, while the tall tree species richness was not influenced by stand age (r = −0.024).

Structural equation models linking tall tree aboveground biomass (TTB) and tall tree species richness (TTR) with the influences of climate, soil, and stand age factors. Solid lines indicate significant paths (P < 0.05), and dashed lines indicate non-significant paths (P ≥ 0.05). TTB, tall tree biomass; STR, short tree richness; SNR, soil nutrient regime; GDD, growing degree days; CMI, climate moisture index; SA, stand age; SRMR, standardized root mean square residual; GFI, goodness-of-fit index.
Effects of the tall tree strata on the species richness and aboveground relationship in the forest understory layer
The structural equation models showed good fits with the data from the forest understory layer (Fig. 4a–d). The species richness of the forest understory layer had a direct positive effect on the corresponding aboveground biomass. Stand age had no significant relationship with species richness or aboveground biomass in the short tree, shrub and herb layers (Fig. 4a–c). The tall tree species richness had a positive direct effect on the short tree species richness and indirectly affected the short tree aboveground biomass through the short tree species richness along with the greater species richness in the tall tree layer and the greater short tree aboveground biomass. However, the tall tree species richness had a negative indirect effect on the short tree aboveground biomass through the tall tree aboveground biomass. Growing degree days and the climate moisture index affected the short tree aboveground biomass through the short tree species richness. The short tree aboveground biomass increased with greater growing degree days but decreased with climate moisture index.

Structural equation models linking aboveground biomass and species richness across forest strata, including the short tree (a), shrub (b), herb(c) and liana (d) layers. STR, SR, HR and LR represent the species richness of the short tree, shrub, herb and liana layers, respectively. STB, SB, HB and LB represent the aboveground biomass of the short tree, shrub, herb and liana layers, respectively. The other abbreviations are explained in Fig. 3.
The soil nutrient regime and growing degree days affected the shrub aboveground biomass through the shrub species richness (Fig. 4b). The soil nutrient regime, tall tree aboveground biomass and species richness affected the herb aboveground biomass through the herb species richness, but the soil nutrient regime had an indirect negative effect on the shrub aboveground biomass (Fig. 4c). The tall tree aboveground biomass showed a direct positive correlation with the aboveground biomass of the liana layer, while the tall tree species richness indirectly affected the liana aboveground biomass through the liana species richness and tall tree biomass. In addition, stand age affected the liana aboveground biomass through the liana species richness (Fig. 4d). The climate moisture index showed no influence on the aboveground biomass in the shrub, herb or liana layers, while growing degree days had no influence on the aboveground biomass in the herb and liana layers.
Discussion
Our findings show that the positive relationship between species richness and aboveground biomass is ubiquitous across forest strata, and widely exists in all forest vegetation strata in primary Pinus kesiya forests. Such relationships are consistent with previous studies that have found a positive association between species richness and aboveground biomass in most forest ecosystem5,7,12. Furthermore, we provide more evidence for this relationship across forest vegetation strata, especially in subtropical pine-oak forests. These results differ from those found in a study by Ali and Yan, which indicated that the tall tree layer had negligible effects on the short tree layer in terms of aboveground biomass14, while our results show that the tall tree aboveground biomass had a negative effect on the short tree aboveground biomass in the primary Pinus kesiya forest ecosystem, which covers a large area of the subtropical zone in Yunnan Province, SW China. The reason for these findings may be that distinguishing between the tall tree and short tree vegetation strata may be more complex and more difficult in evergreen broadleaf forests without the presence of productive pines along with the weakening of niche partitioning in the vertical spaces of the forest.
We observed that direct and indirect path processes mediated the relationships between species richness and aboveground biomass across the forest vegetation strata in a primary Pinus kesiya forest. As a rule, existing studies have demonstrated that the species richness of the tall tree strata was dependent on the macroclimate at a relatively large scale, which could increase the abundance of dominant species and enhance light utilization efficiency in the tall tree strata13,16. Pinus kesiya has been described as a species with relatively high production due to its prolonged photosynthetic activity and high nutrient uptake through the rapid turnover of nutrients in subtropical pine-oak forests21. This species is often considered to be the strongest competitor and has many large diameter individuals; it is therefore the main contributor to woody biomass production in these forests, which dominates the relationship between species richness and aboveground biomass in the tall tree layer. Larger trees of Pinus kesiya have a greater contribution to the aboveground biomass than smaller diameter trees in old-growth forests21. Although specific driving mechanisms were not analysed in our study, the role of Pinus kesiya could be potentially explained by the selection effect15,18. This species may have been the most productive species in the local species pools at the beginning of the establishment of the forest community and gradually became the most dominant species in the tall tree strata, which negatively affected the growth and regeneration of short tree species15. Our results show that the tall tree aboveground biomass had a negative effect on the aboveground biomass of the short tree and shrub layers, which could also be better explained by the amount of light-blocking tissue in the canopy of tall tree species.
The tall tree species richness significantly promoted the aboveground biomass of the short tree and liana layers. The short tree species are predominantly mountainous subtropical broadleaf species20, which could prevent strong interspecific competition and promote greater site resource utilization along with tall tree species according to the complementarity effect7. The coexistence of short tree species was beneficial in increasing the woody biomass production due to enhanced resource use efficiency by means of water and nutrient uptake facilitation, niche partitioning and the control of herbivores and disease24. Short tree species have specific environmental requirements and can maintain the species diversity in primary Pinus kesiya forests even if some species exhibit a lower woody biomass production which could also offset the negative effects through niche partitioning. Recent studies have suggested that coexisting species were more functionally similar or redundant in species-rich communities4. Our results support this idea that shade-tolerant and evergreen broadleaf species composed the short tree strata and demonstrated more niche overlap. The strength of the relationship between species richness and aboveground biomass in the short tree strata was greater than that in the tall tree layer. Because of the lack of highly productive species in the short tree strata, the short tree species also had a lower contribution to the total aboveground biomass across forest vegetation strata. In addition, the trees of larger diameter are important host trees in subtropical forest ecosystems and are beneficial to the establishment and growth of lianas. The larger trunks of most Pinus kesiya individuals and other canopy tree species facilitated climbing by liana species, and therefore, the species richness of the tall tree strata increased with the aboveground biomass production of lianas in accordance with stand age.
Stand age had a positive influence on the tall tree aboveground biomass accounting for 80.31% of the variance. It is widely believed that stand age is a critical driver of individual species dynamics, standing biomass and productivity9. Although the net primary productivity of old-growth stands is lower than that in younger stands of primary Pinus kesiya forests21, it was confirmed that the aboveground biomass increased with stand age in this primary Pinus kesiya forest20. Our results indicate that stand age had no impact on the aboveground biomass of the short tree, shrub, or herb layers. The magnitude of the positive diversity-productivity relationship in the forest short tree layer was affected by the shade of canopy trees and the environmental conditions7.
Despite the relatively low contribution of the shrub and herb layers to the total aboveground biomass, they contributed a substantial proportion of the annual litter fall which contributes to the total annual ecosystem nutrient uptake25. Litter fall and fine root decomposition change the supply of N and P and their availability in the soil, resulting in changes in plant growth and biomass production. A previous study showed that lower soil N:P tatios promoted the growth and biomass production of the shrub and herb layers26, which is in agreement with our results. However, the N:P ratio had a positive indirect influence on the tall tree aboveground biomass by enhancing plant photosynthesis as a result of greater N availability. In addition, climate factors also affected the species richness of the short tree strata, and environmental heterogeneity had an important effect on the short tree strata species diversity11. We observed that the short tree strata accounted for most of the vascular plant species diversity in the primary Pinus keisya forest, in contrast to the low tall tree species diversity. Short tree species have specific niches and are better able to adapt to different environmental conditions, which supports the idea that the climate moisture index, growing degree days and the soil nutrient regime affect the aboveground biomass across forest vegetation strata, especially in the tall tree layer7,12. Our results are consistent with those from a recent study on the relationship between species richness and biomass from boreal to subtropical forests in China4.
Conclusions
The positive relationship between species richness and aboveground biomass found in our study is similar to those described in previous studies, indicating that this relationship is ubiquitous across forest vegetation strata at large spatial scales. Moreover, our results also show that the relationship in the tall tree strata was weaker than that in the shrub, herb and liana layers. However, the tall tree species richness was positively correlated with the species richness and aboveground biomass of the short tree and liana layers. In contrast, the species richness of the tall tree strata had an indirect negative effect on the aboveground biomass of the herb layer. The species richness and aboveground biomass in the tall tree strata had a complex effect on the biodiversity-biomass relationship across forest vegetation strata in accordance with two important mechanisms of biological maintenance: the selection effect and the complementarity effect. The tall tree species had an obvious resource-filtering effect, which strengthened the intensity and magnitude of the positive biodiversity-biomass relationships in the forest understory strata. Furthermore, climate factors had an important influence on the positive biomass-richness relationship in the tall tree strata. The soil nutrient regime and shading by tall tree species directly or indirectly affected the species richness-biomass relationships in the short tree, shrub, and herb layers. Our study provides new insight into the ecology of the primary Pinus kesiya forest, and the results almost did not show an effect of species loss on biomass or productivity because of anthropogenic disturbance. For a better understanding of the biodiversity-biomass relationship, long-term observations will be essential in future studies.
Materials and Methods
Study area and site description
The study was conducted in primary Pinus kesiya forests that are mainly distributed in the southern and southwestern parts of Yunnan Province, China. This area has mountainous topography, with an altitude ranging from 850 m to 1850 m. This region has a typical monsoon climate with two distinct seasons: a rainy season (from May to October) and a dry season (from November to April). The annual mean temperature ranges from 17 °C to 18.5 °C. The annual precipitation ranges from 1100 mm to 1550 mm, with favourable hydrothermal conditions, and more than 80% of the annual precipitation falls during the rainy season23.
Pinus kesiya and Schima wallichii are the dominant canopy tree species in all the sampled stands. The subcanopy tree species in the forest are Castanopsis hystrix, Castanopsis echidnocarpa, Castanopsis calathiformis, Castanopsis fleuryi, Lithocarpus truncatus, Lithocarpus fenestratus, Machilus rufipes, Betula alnoides, Rhus chinensis, and Alnus cremastogyne. The shrub layer is dominated by Symplocos paniculata, Breynia fruticosa, Ficus hirta, Melastoma affine, Glochidion eriocarpum, and Inula cappa. The herb layer is dominated by Eupatorium adenophorum, Zingiber densissimum, Hedychium flavum, Imperata cylindrica var. major, Woodwardia japonica, Dicranopteris pedata, Scleria levis, and Dianella ensifolia. Mucuna macrocarpa, Craspedolobium schochii, Embelia ribes, Smilax spp., Gnetum montanum, and Celastrus monospermus are the dominant liana species in the primary Pinus keisya forest.
Forest inventory data
The data were from 170 field sampling plots located in nine counties, including Simao, Jinghong, Menghai, Jinggu, Zhenyuan, Jingdong, Yunxian, Changning and Lianghe in southern and southwestern Yunnan Province, China. The plots ranged from 22°11′ to 24°38′ N and from 22°11′ to 24°38′ E. The plots were 20 m × 20 m (400 m2) based on the requirement in Yunnan vegetation23. All plots were measured between 2012 and 2014 (the distribution of plots is shown in Fig. 5). Within each plot, the forest vegetation was classified as belonging to the tall tree, short tree, shrub, herb or liana layers. All trees, lianas and shrubs were measured for their diameter at breast height (DBH) if they had a DBH ≥ 1 cm. Five 2 m × 2 m subplots were used for the survey of the herb layer, including tree seedlings, shrubs and herbaceous plants. Plant species, number, height and environmental factors, including altitude, slope, aspect, and slope position, were recorded. We defined all stems in each plot with DBH ≥ 9 cm as part of the tall tree layer; the short tree layer included all trees with DBH < 9 cm and height ≥1.3 m as well as shrubs ≥1.3 m in height. The shrub layer included all shrubs and trees <1.3 m in height. The liana layer included lianas with stems of all different sizes in the plots. The herb layer included all non-woody vascular plants, such as ferns, graminoids and saprophytes12. We used species richness as a species diversity index for the tall tree, short tree, shrub, herb, and liana layers. We recorded a total of 203 trees, 44 shrubs, 102 herbs and 41 lianas in all plots. Stand age (SA) was determined as the mean age of the three largest Pinus kesiya individuals. We fitted an age-DBH equation for Pinus kesiya using the formula y = 3.326DBH0.733 (y is the tree age, R2 = 0.802, P < 0.001, F = 357.323, n = 90).

The locations of the plots in the study area. The map was generated using the software ArcGIS 10 (Environmental Systems Research Institute, Inc. Redlands, CA, USA; http://www.esri.com).
Estimation of aboveground biomass
Aboveground biomass (AGB, Mg·ha−1) across forest vegetation strata was estimated as the sum of the stems, branches, and leaves using allometric equations including trees with DBH ≥ 5 cm, shrubs and small trees with DBH < 5 cm, and lianas (DBH ≥ 2 cm) (Table 1). We classified species occurring in the forests into the following 9 plant functional groups and obtained their aboveground biomass using species-specific allometric equations based on DBH, height or length: (1) Pinus kesiya27, (2) Schima wallichii (the allometric equation was built by the authors), (3) Castanopsis hystrix and Fagaceae species with similar functional traits28, (4) Betula alnoides29, (5) Rhus chinensis30, (6) Alnus cremastogyne31, (7) other mixed tree species for less frequently occurring species32, (8) shrubs and small trees33, and (9) lianas34. We used a destructive harvesting method to measure the aboveground biomass of the herb layer, including herbs, shrubs and tree seedlings (DBH ≤ 1 cm)20.
Environmental factors
The soil nutrient regime (SNR), growing degree days (GDD, °C) and climate moisture index (CMI, mm, annual precipitation minus annual potential evapotranspiration) were used to account for the effect of environmental conditions and stand growth on aboveground biomass. We used the N:P ratio as a measure of the soil nutrient regime to represent local site conditions. Climate data were obtained from the Climate AP software and have been revised for use in the Asia-Pacific region35. The GDD and CMI values represented the overall energy available for plant growth and water availability for plants7.
Data analysis
We regressed the species richness on aboveground biomass across forest vegetation strata to assess the relationships between these two variables using ordinary least squares (OLS) regressions with a cubic smoothing spline. The cumulative value of aboveground biomass was plotted against the cumulative species number (these species represent 99.53% of the total biomass in the landscape) according to the magnitude of the aboveground biomass of each species in all plots, which could indicate the relative contribution of the species to the aboveground biomass17. We used a structural equation model (SEM) in our analyses to determine the effects of response variables, which were related to the tall tree and environmental conditions on the relationships between the species richness and aboveground biomass of different short tree layers.
In addition, we further fitted an SEM to interpret the casual effects and relative importance of stand age, soil nutrient regime, growing degree days, climate moisture index, species richness, and aboveground biomass of the tall tree layer on the species richness and aboveground biomass of the short tree, shrub, herb and liana layers (Table S1). The SEM in our study was based on the theoretical multivariate causes of plant diversity and ecosystem functioning in natural forests36. We explicitly defined two main paths: (1) the relationships between species richness and aboveground biomass within each layer, and (2) the effects of covariates, including GDD, SNR, CMI, SA and the tall tree layer, on the short tree layer. As recommended by Zhang et al.7, the GDD, SNR, SA, CMI and aboveground biomass across forest vegetation strata were transformed using the natural log scale. The standardized root mean square residual (SRMR) and goodness-of-fit index (GFI) were used to evaluate the goodness of fit of the SEMs with GFI values >0.95 and SRMR values <0.08. We report the standardized coefficient for each path from each component model. The total effects, as well as the direct and indirect standardized effects, were also calculated to strengthen the interpretation of our SEM results. The SEM analysis was implemented using the “lavaan” package in R 3.3.237 (R development Core Team 2016).
References
Gross, K. et al. Species richness and the temporal stability of biomass production: a new analysis of recent biodiversity experiments. Am Nat 183, 1–12 (2014).
Barnes, A. D. et al. Species richness and biomass explain spatial turnover in ecosystem functioning across tropical and temperate ecosystems. Philo T R Soc B 371, https://doi.org/10.1098/rstb.2015.0279 (2016).
Bunker, D. E. et al. Species loss and aboveground carbon storage in a tropical forest. Science 310, 1029–1031 (2005).
Wu, X. et al. The relationship between species richness and biomass changes from boreal to subtropical in China. Ecography 38, 602–613 (2014).
Liang, J. J. et al. Positive biodiversity-productivity relationship predominant in global forests. Science 63, 196–211 (2016).
Gamfeldt, L. et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat Commun 4, 1340 (2013).
Zhang, Y., Chen, H. Y. H. & Taylor, A. R. Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstorey trees. Funct Ecol 31, 1–8 (2016).
Jenkins, D. G. Estimating ecological production from biomass. Ecosphere 6, 1–31 (2015).
Zhang, Y. & Chen, H. Y. H. Individual size inequality links forest diversity and above-ground biomass. J Ecol 103, 1245–1252 (2015).
Laughlin, D. C. & Grace, J. B. A multivariate model of plant species richness in forested systems: old-growth montane forests with a long history of fire. Oikos 114, 60–70 (2006).
Evy, A. et al. Driving mechanisms of overstory-understory diversity relationship in European forests. Perspect Plant Ecol 19, 21–29 (2016).
Zhang, Y., Chen, H. Y. H. & Taylor, A. R. Aboveground biomass of understory vegetation has a negligible or negative association with overstory tree species diversity in natural forests. Global Ecol Biogeogr 2, 141–150 (2015).
Reich, P. B., Frelich, L. E., Voldseth, R. A., Bakken, P. & Adair, E. C. Understory diversity in southern boreal forests is regulated by productivity and its indirect impacts on resource availability and heterogeneity. J Ecol 100, 539–545 (2012).
Ali, A. & Yan, E. R. The forest strata-dependent relationship between biodiversity and aboveground biomass within a subtropical forest. Forest Ecol Manag 401, 125–134 (2017).
Loreau, M. & Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 412, 72–76 (2001).
Oberle, B., Grace, J. B. & Chase, J. M. Beneath the veil: plant growth form influences the strength of species richness-productivity relationships in forests. Global Ecol Biogeogr 18, 416–425 (2009).
Lohbeck, M., Bongers, F., Martinez-Ramos, M. & Poorter, L. The importance of biodiversity and dominance for multiple ecosystem functions in a human-modified tropical landscape. Ecology 97, 2772–2779 (2016).
Cardinale, B. J. et al. Impacts of plant diversity on biomass production increase through time because of species complementarity. P Natl Acad Sci USA 104, 18123–18128 (2007).
Zhang, Y., Chen, H. Y. H. & Taylor, A. Multiple dirvers of plant diversity in forest ecosystems. Global Ecol Biogeogr 23, 885–893 (2014).
Li, S. F. et al. Changes in biomass carbon and soil organic carbon stocks following the conversion from a secondary coniferous forest to a pine plantation. PLoS ONE 10, e0135946, https://doi.org/10.1371/journal.pone.0135946 (2015).
Baishya, R. & Barik, S. K. Estimation of tree biomass, carbon pool and net primary production of an old-growth Pinus kesiya Royle ex. Gordon forest in north-eastern India. Ann For Sci 68, 727–736 (2011).
Zonneveld, M. V., Koskela, J., Vinceti, B. & Jarvis, A. Impact of climate change on the distribution of tropical pines in southern Asia. Unasylva 60, 24–29 (2009).
Wu, Z. Y., Zhu, Y. C. & Jiang, H. Q. Yunnan vegetation. 398–416 (Beijing, Science Publish Press, 1987).
Aguiar, M. I. D. et al. Does biomass production depend on plant communtiy diversity? Agroforest Syst 87, 699–711 (2013).
Nilssion, M. C. & Wardle, D. A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Front Ecol Environ 3, 421–428 (2008).
Elser, J. J. et al. Global analysis of nitrogen and phosphorus limition of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10, 1135–1142 (2007).
Chen, Q. et al. Biomass and carbon storage of Pinus kesiya var. langbianensis in Puer’ Yunnan. J Yunnan Univ Nat Sci 36, 439–445 (2014).
He, Y. J. et al. Carbon storage capacity of monoculuture and mixed-species plantations in subtropical China. Forest Ecol Manag 295, 193–198 (2013).
Liang, N., Wang, W. B., Ni, J. B. & Tian, K. A study on biomass in sapling stage of pure Betula alnoides forest with Betula alnoides and Cinnamomum cassia mixed forest. Journal of West China Forestry. Science 36, 44–49 (2007).
Zhang, H. Q., Liu, Q. J., Lu, P. L., Yu, Q. & Zeng, H. Q. Biomass estimation of several common shrubs in Qianyanzhou experimental station. Forest Invent Plann 30(5), 43–49 (2005).
Li, G. X. et al. Characteristics of Almus cremastogyne plantation community and its biomass in central Yunnan Plateau. J Zhejiang Forest Univ 23, 362–366 (2006).
Zuo, S. D., Ren, Y., Weng, X., Ding, H. F. & Luo, Y. J. Biomass allometric equations of nine common tree species in an evergreen broadleaved forest of subtropical China. Chinese J Appl Ecol 26, 356–362 (2015).
Ali, A. et al. Allometric biomass equations for shrub and small tree species in subtropcial China. Silva Fenn 49, 1–10 (2015).
Zheng, Z., Feng, Z., Cao, M., Li, Z. & Zhang, J. Forest structure and biomass of a tropical seasonal rain forest in Xishuangbanna, southwest China. Biotropica 38, 318–327 (2006).
Wang, G. Y. et al. Adaptation of Asia-Pacific forests to climate change. J Forestry Res 27, 469–488 (2016).
Grace, J. B. et al. Intergrative modelling reveals mechanisms linking productivity and plant species richness. Science 529, 390–399 (2016).
Rosseel, Y. Lavaan: an R package for structural equation modeling. J Stat Softw 48, 1–36 (2012).
Acknowledgements
We thank the staff members of the forest bureaus and forest farms in Jinghong, Menghai, Lianghe, Changning, Yunxian, Jingdong, Simao, Zhenyuan and Jinggu Counties for the data collection. This work was supported by the Fundamental Research Funds of CAF (CAFYBB2017ZX002) and the Yunnan Provincial Science and Technology Department (2013RA004). We would also like to thank Alison Beamish at the British Columbia University and the Nature Research Editing Service for their assistance with the English language and grammatical editing of the manuscript.
Author information
Authors and Affiliations
Contributions
Shuaifeng Li conducted the field measurements and drafted the manuscript, Jianrong Su conceived the study and revised the manuscript, Xuedong Lang and Wande Liu conducted the field measurements, Guanglong Ou constructed the model between stand age and DBH.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, S., Su, J., Lang, X. et al. Positive relationship between species richness and aboveground biomass across forest strata in a primary Pinus kesiya forest. Sci Rep 8, 2227 (2018). https://doi.org/10.1038/s41598-018-20165-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-20165-y
This article is cited by
-
Monitoring carbon stock changes in Solan Forest Division of Indian Western Himalayas
Environment, Development and Sustainability (2023)
-
QTL mapping for growth-related traits by constructing the first genetic linkage map in Simao pine
BMC Plant Biology (2022)
-
Tree biomass and carbon stock in subtropical Sal forest of Central Himalaya, India
Vegetos (2022)
-
Stand structure and species diversity regulate biomass carbon stock under major Central Himalayan forest types of India
Ecological Processes (2021)
-
Interpreting forest diversity-productivity relationships: volume values, disturbance histories and alternative inferences
Forest Ecosystems (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
