
Abstract
Macrofauna is known to inhabit the top few 10s cm of marine sediments, with rare burrows up to two metres below the seabed. Here, we provide evidence from deep-water Permian strata for a previously unrecognised habitat up to at least 8 metres below the sediment-water interface. Infaunal organisms exploited networks of forcibly injected sand below the seabed, forming living traces and reworking sediment. This is the first record that shows sediment injections are responsible for hosting macrofaunal life metres below the contemporaneous seabed. In addition, given the widespread occurrence of thick sandy successions that accumulate in deep-water settings, macrofauna living in the deep biosphere are likely much more prevalent than considered previously. These findings should influence future sampling strategies to better constrain the depth range of infaunal animals living in modern deep-sea sands. One Sentence Summary: The living depth of infaunal macrofauna is shown to reach at least 8 metres in new habitats associated with sand injections.
Similar content being viewed by others

Introduction
Deep-sea infauna is one of the most elusive branches of life on Earth; little is known about modern deep seabed environments, and less about the ancient. The limits of the macrofaunal biosphere in the deep-sea, and factors controlling life at depth below the seabed, are generally unknown.
In the modern, it is technologically challenging to collect undisturbed samples, and burrowing animals are usually found in marine sediments down to 20 cm1, or occasionally as deep as 2 m2. In ancient successions, the primary archive of deep zones of macrofaunal life is the ichnological (trace fossil) record in sedimentary rocks, which is limited by preservation factors with poor constraint on the original depth. Metre-scale (1–2 metre long) post-depositional burrows have been recorded on the base of turbidite sandstones, where individual depositional events are documented, providing depth control on infauna3,4. Rare cases of fauna extending beyond this depth require an open conduit in firm ground to allow filtration of seawater (e.g.5,6,7).
We have studied exhumed networks of clastic intrusions (injectites) produced by the injection of overpressured sand into surrounding strata (e.g.8). These injected sand dykes (vertical to sub vertical) and sills (horizontal) show evidence for post-injection living traces of macrofauna along their surfaces. Previously, injectites have been identified as favourable sites for colonisation by microbial life because they are permeable and provide a large sand-to-mud interface allowing for readily available electron donors and nutrients9. Here, we show that macrofauna also lived in injectites deep below the contemporaneous seabed.
Geological background and dataset
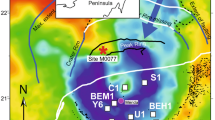
Three separate Permian outcrop sites from the SW Karoo Basin, South Africa (Fig. 1) exhibit sand injectites sourced from deep-marine turbidite sands in the Fort Brown Formation. Bioturbation is documented throughout this formation10 and ichnological assemblages include Thalassinoides and Planolites 11, commonly observed as hypichnia on sandstone bed bases (Fig. 2). At each site, the units have sharp and erosional bases (Fig. 2), and depositional architecture, and regional mapping demonstrates that they form stacked submarine lobe deposits12,13.

Location map of Laingsburg depocentre and outcrop sites 1–3, South Africa. Site 1 = Unit E, Geelbeck, Site 2 = Unit D, Slagtersfontein West, Site 3 = Unit D, Slagtersfontein East. Figure prepared in CorelDRAW × 8 (http://www.coreldraw.com/en/).

(A) A representative cross-section panel for outcrop sites with several stratigraphic logs taken from South to North. Here, Unit E at Geelbeck (Site 1) displays turbidite sandstones with underlying sandstone injections. Vertical to subvertical injectites are dykes, horizontal injectites are sills (see arrows). (B) Example of typical bioturbation seen on the base of Subunit E2 (see C). (C) Outcrop photograph demonstrating sharp and erosional contact between lobe sandstones (Subunit E2) and injectites below.
The injectites are sourced from fine sandstones, and are up to 0.5 m thick, sharp-sided, and propagated upwards and/or downwards and are discordant (dykes) or concordant (sills) to the stratigraphy (Fig. 2). A combination of 2 dimensional outcrops, a narrow grainsize-range and unvarying provenance ensures that specific source beds of the injectites cannot be discriminated. At sites 1, 2 and 3 (Fig. 1) injectites are found 8 m, 1.5 m and 3 m compacted depths respectively below overlying sandstones. The same trace fossils present on the base of sandstone beds throughout the Fort Brown Formation are observed on the margins of clastic injectites (Fig. 3), down to these lower limits.

Typical examples of bioturbation found on clastic injectite margins. (A) Unit E (Site 1): dewatering structures (Aristophycus) on margin of a subvertical injectite, overprinted by Thalassinoides bioturbation. (B) Unit E (Site 1): Planolites tube protruding in cross-section of sill, planform of tube is outlined on the top margin of the sill. (C) view of (B) from alternate orientation. (D) Unit E (Site 1): Cross-cutting Planolites on base of sill. (E) Unit D (Site 2): Dyke margin with several examples of bioturbation, largest Planolites are indicated. (F) Unit C (Site 3): Dyke margin with several, smaller Planolites.
Interpretation
The structures on injectite margins are interpreted as trace fossils, and not grooves or markings formed through the injection process because they include the branching structure of Thalassinoides and gently sinuous burrows of Planolites (Fig. 3A). Additionally, the structures show random orientations (Fig. 3D, F) fitting a bioturbation origin, whereas grooves would have a preferential direction caused by flow interacting with injectite-margins. It is clear that the bioturbation occurred after emplacement of the clastic injectites as it follows the sand-mud interface on both subvertical (dykes) and horizontal (sills) injectites (Fig. 3). If bioturbation on the injectites were casts of previously buried burrows then Planolites would be expected along sills only, parallel to stratal contacts. Traces in full relief are also observed on the top and bases of injectites, distinguishing them from seabed bioturbation, which will only have burrows in full relief on the lower side (Fig. 4B). In some cases, bioturbation overprints the dewatering structure Aristophycus (Fig. 3A), showing that clastic injection was followed by dewatering and then bioturbation. From these deductions, we can determine that Planolites and Thalassinoides formed after clastic injection. Forcible intrusion of sand into mud occurs in deep sea sediments in many sedimentary basins (e.g.14). Therefore, our discovery represents a widespread and unexplored macrofaunal environment. Previously, the organisms forming Planolites and Thalassinoides have been believed to have lived mainly in the top 20 cm of sediment, rarely reaching maximum depths of 1.5 m3,15. This limited depth range is due to the decline of oxygen and organic matter (food) in deeper levels of the sediment. Here, we document the presence of bioturbation on the margins of clastic injectites found at least 8 m below the seabed. In order to produce traces, organisms would need to survive long enough to burrow for hours-to-days. The size of the burrows (4–10 mm in diameter) suggests macro-infaunal invertebrates. We suggest 3 possible origins for the injected sand.

Different scenarios to explain presence of macrofaunal communities preservation in injectites. (A) Sandy lobe unit, with bioturbation along the sand-to-mud interface at the base (B), several metres below the seabed. Overpressure at the edge of the sandy lobes after rapid burial by muds causes unconsolidated sand to forcible intrude into underlying mud outwards from lobe centre. Macrofauna form new living traces on the sand-to-mud contacts that form the margins of the sand intrusions (B). Scenarios where a pre-existing injectite network either formed connection to the seabed through extrusion (C) or exhumation (D) to be exploited by macrofauna. Average lobe thickness in (A) based on26.
Discussion
In the first scenario, injectites propagate downwards, sourced from overlying sands. This requires overpressure within the source sands in order to inject mixed sand and water downwards into the substrate. For overpressure to occur sands must be sealed by muds and there must be sufficient overburden (at least as thick as injectites are deep to promote downward injection). Turbidity currents deposit event beds that can be muddy16, occurring over hours to days, which could provide the necessary seal over a lobe for pressure to build during burial (Fig. 4A). In this case, macrofauna would have been living at several metres depth post-burial before being injected down with the flow into the substrate, whereupon they burrowed in this new setting. The concept of animals surviving both considerable transportation, and then living in oxygen-depleted environments on the seabed is well known (doomed pioneers;17). Alternatively, the fauna migrated down along the new network of sand, post injection.
In the second scenario, if injectites were sourced from below, then the host mudrock provides the seal required for overpressure and injection. Injectites could establish a seabed connection through extrusion18 allowing burrowers to then penetrate downwards (Fig. 4C). In the third scenario, a preexisting injectite network in the host mudrock, with a parent sand from below or above, is exhumed through seascape degradation (e.g.19). Organisms could then burrow downwards to exploit the injectite network as it acts as a source of new organic matter (Fig. 4D). The presence of thin overlying siltstones in the three examples (see supplementary text for detail), suggests that the lobes were not buried rapidly by mud-rich flows. These overlying siltstones, and the erosional basal contacts of the lobes, instead support exploitation by macrofauna of preexisting injectite networks through extrusion, exhumation, or a combination of both mechanisms.
In the above cases, for bioturbation to take place in the injectite in situ, the organisms would have had to survive i) potentially limited oxygen and POM (particulate organic matter) supply, ii) overburden pressures associated with injection depths, and if transported during injection iii) initial high energy transport. Life in a deep-marine environment often has a slow metabolic rate to survive cold temperatures and energy deprivation20,21. In the latter two scenarios above, it is possible for oxygen and POM to permeate the injectite network through connection to the seafloor. Where the injectite network is sealed, oxygen and nutrients are not replaced. Here, we model the possible survival time of organisms, parameterised by data based on contemporary polychaete populations, within an injectite as oxygen is depleted by respiration. We have estimated a time-frame which the organisms could survive within the injectite without replenishment of oxygen to give a potential life-span in a closed system using equation 1.
where: t is time (days), [O2] is the dissolved oxygen at time t (ml/L), [O2i] is the initial (t = 0) dissolved oxygen concentration of the sediment (ml/L), SCOC is the Sediment Community Oxygen Consumption, which accounts for all the bacterial, meiofaunal and macrofaunal metabolic activity (mlO2/L/day), N is the abundance of polychaetes (number/L), Mr is the metabolic rate of polychaetes (mlO2/day), SX is the proportion of survival of injected polychaetes and Sb the proportional survival of the background community.
Parameterisation using contemporary analogues (see Supplementary Materials) shows that macrofaunal organisms with a slow metabolism rate could potentially survive for up to 270 days with no replenishment of O2 (Fig. S3). If the injected sand was sourced from above, this period is required to overpressure the sand body by depositing muddy flows, inject, and produce the traces we observe. If sand is sourced from below, 270 days is the minimum time for oxygen depletion if surface burrow connections cannot be maintained, providing ample time for fauna to burrow downwards and exploit injected sand networks (Fig. 4).
Our study highlights a mismatch between observations taken of ancient and modern environments. Modern deep-sea biological studies target clays and silts as these are simpler to sample. Standard sampling methodologies, such as piston coring, are typically unable to sample sandy sediments due to lack of cohesion of the grains. In contrast, rare examples of deep tier bioturbation from the rock record have been able to demonstrate that burrowing occurred in deep-marine sands at 1–2 metres below the contemporaneous seabed3,4,22. Here we demonstrate burrowing to much greater depths, up to an uncompacted depth of 8 metres. The high degree of compaction of fine-grained sediments at such depth is likely to make burrowing activity unlikely whereas the deep sand burrowing we document here suggests such activity may be common-place (but unsampled in the present day).
Whilst we have demonstrated in these examples that macrofauna were present in the deep biosphere, a key question is whether these are exceptional cases related to unique conditions, or whether the conditions conducive to burrowing of macrofauna to multi-metre depths are widespread. Sand-rich sedimentary environments, in the form of lobe deposits and channel-fills are present over very large portions of submarine fans (e.g.12,23,24,25,26), themselves the largest sedimentary deposits on Earth27. Macrofauna might be expected to burrow to depth in many of these sub-environments, however, the present examples show unique evidence that the depth reached is preserved. In most cases, the macrofauna will be preserved in the rock record but the information on their depth below the contemporaneous seabed is lost. More recently, there has been a rapidly growing recognition of sand injection in many deep-marine settings, from channels on the slope28 to lobes on the basin floor29,30. Given the ubiquity and thickness of sand-rich sedimentary successions in many deep-sea environments, and the widespread occurrence of sediment injection then it would seem probable that such macrofauna in the deep biosphere are more prevalent than currently recognised.
Conclusions
Our findings have several biological and geological implications, i) unusually, we can quantify a minimum depth below the seabed that organisms inhabited in ancient sediments, ii) show that the deepest organisms may be present in sandy sediments, rather than the clays and silts typically targeted in modern seabed investigations, iii) show that less organics are preserved due to carbon consumption during metabolic activity, which then also changes the sediment fabric at depth, with grains being processed and sorted into burrow structures, and iv) most importantly, we have shown that macrofaunal life survives for periods living at depths of up to 8 m below the seabed, giving an entirely new (lower) limit to the macrofaunal biosphere. This new evidence of bioturbation in Permian sandstones at many metres below the seabed suggests that we need to adapt sampling strategies when looking for macrofaunal life in the deep biosphere. Targeting sandy successions in deep-marine systems offers potential for observing the behaviour and diversity of organisms at greater depths than has so far been hitherto appreciated in modern deep-sea sediments.
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary information files)31,32,33,34,35,36,37,38,39,40,41.
References
Jumars, P. A. Spatial autocorrelation with RUM (Remote Underwater Manipulator): vertical and horizontal structure of a bathyal benthic community. Deep-Sea Res. 25, 589–604 (1978).
Weaver, P. P. E. & Schultheiss, P. J. Vertical open burrows in deep-sea sediments 2 m in length. Nature 301, 329–331 (1983).
Heard, T. & Pickering, K. T. Trace fossils as diagnostic indicators of deep-marine environments, Middle Eocene Ainsa – Jaca basin, Spanish Pyrenees. Sedimentology 55, 809–844 (2008).
Cummings, J. P. & Hodgson, D. M. Assessing controls on the distribution of ichnotaxa in submarine fan environments, the Basque Basin, Northern Spain. Sed. Geol. 239, 162–187 (2011).
Savrda, C. E., Browning, J. V., Krawinkel, H. & Hesselbo, S. P. Firmground ichnofabrics in deep-water sequence stratigraphy, Tertiary clinoform-toe deposits, New Jersey slope. Palaios 16, 294–305 (2001).
Hubbard, S. M. & Shultz, M. R. Deep burrows in submarine fan-channel deposits of the Cerro Toro Formation (Cretaceous), Chilean Patagonia: implications for firmground development and colonization in the deep sea. Palaios 23, 223–232 (2008).
Callow, R. H., McIlroy, D., Kneller, B. & Dykstra, M. Integrated ichnological and sedimentological analysis of a Late Cretaceous submarine channel-levee system: The Rosario Formation, Baja California, Mexico. Mar. Pet. Geol. 41, 277–294 (2013).
Jolly, R. J. & Lonergan, L. Mechanisms and controls on the formation of sand intrusions. J. Geol. Soc. London 159, 605–617 (2002).
Parnell, J., Boyce, A. J., Hurst, A., Davidheiser-Kroll, B. & Ponicka, J. Long term geological record of a global deep subsurface microbial habitat in sand injection complexes. Sci. Reports 3, 1828 (2013).
Oliveira, C. M., Hodgson, D. M. & Flint, S. S. Distribution of soft-sediment deformation structures in clinoform successions of the Permian Ecca Group, Karoo Basin, South Africa. Sed. Geol. 235, 314–330 (2011).
Morris, E. A. et al. Sedimentology, stratigraphic architecture, and depositional context of submarine frontal-lobe complexes. J. Sed. Res. 84, 763–780 (2014).
Van de Merwe, W. C., Hodgson, D. M., Brunt, R. L. & Flint, S. S. Depositional architecture of sand-attached and sand-detached channel-lobe transition zones on an exhumed stepped slope mapped over a 2500 km2 area. Geosphere 10, 1076–1093 (2014).
Spychala, Y. T., Hodgson, D. M., Flint, S. S. & Mountney, N. P. Constraining the sedimentology and stratigraphy of submarine intraslope lobe deposits using exhumed examples from the Karoo Basin, South Africa. Sed. Geol. 322, 67–81 (2015).
Huuse, M. et al. Subsurface sediment remobilization and fluid flow in sedimentary basins: an overview. Bas. Res. 22, 342–360 (2010).
Thomson, J. & Wilson, T. R. S. Burrow-like structures at depth in a Cape Basin red clay core. Deep Sea Research Part A. Oceanographic Research Papers 27, 197–202 (1980).
Talling, P. J., Masson, D. G., Sumner, E. J. & Malgesini, G. Subaqueous sediment density flows: depositional processes and deposit types. Sedimentology 59, 1937–2003 (2012).
Föllmi, K. B. & Grimm, K. A. Doomed pioneers: Gravity-flow deposition and bioturbation in marine oxygen-deficient environments. Geology 18, 1069–1072 (1990).
Ross, J. A., Peakall, J. & Keevil, G. M. Sub-aqueous sand extrusion dynamics. J. Geol. Soc. London 170, 593–602 (2013).
Hodgson, D. M., Di Celma, C. N., Brunt, R. L. & Flint, S. S. Submarine slope degradation and aggradation and the stratigraphic evolution of channel-levee systems. J. Geol. Soc. London 168, 625–628 (2011).
Mahaut, M., Sibuet, M. & Shirayama, Y. Weight-dependent respiration rates in deep-sea organisms. Deep-Sea Res. Pt I: Oceanographic Research Papers 42, 1575–1582 (1995).
McClain, C. R., Allen, A. P., Tittensor, D. P. & Rex, M. A. Energetics of life on the deep seafloor. PNAS 109, 15366–15371 (2012).
Heard, T. G., Pickering, K. T. & Clark, J. D. Ichnofabric characterization of a deep‐marine clastic system: a subsurface study of the Middle Eocene Ainsa System, Spanish Pyrenees. Sedimentology 61, 1298–1331 (2014).
Normark, W. R. Fan valleys, channels, and depositional lobes on modern submarine fans: characters for recognition of sandy turbidite environments. AAPG Bulletin 62, 912–931 (1978).
Richards, M., Bowman, M. & Reading, H. Submarine-fan systems I: characterization and stratigraphic prediction. Mar. Pet. Geol. 15, 689–717 (1998).
Piper, D. J. & Normark, W. R. Sandy fans–from Amazon to Hueneme and beyond. AAPG Bulletin 85, 1407–1438 (2001).
Prélat, A., Hodgson, D. M. & Flint, S. S. Evolution, architecture and hierarchy of distributary deep‐water deposits: a high‐resolution outcrop investigation from the Permian Karoo Basin, South Africa. Sedimentology 56, 2132–2154 (2009).
Curray, J. R., Emmel, F. J. & Moore, D. G. The Bengal Fan: morphology, geometry, stratigraphy, history and processes. Mar. Pet. Geol. 19, 1191–1223 (2002).
Hurst, A., Scott, A. & Vigorito, M. Physical characteristics of sand injectites. Earth-Sci. Rev. 106, 215–246 (2011).
Cobain, S. L., Peakall, J. & Hodgson, D. M. Indicators of propagation direction and relative depth in clastic injectites: Implications for laminar versus turbulent flow processes. Geol. Soc. Am. Bull. 127, 1816–1830 (2015).
Cobain, S. L., Hodgson, D. M., Peakall, J. & Shiers, M. N. An integrated model of clastic injectites and basin floor lobe complexes: Implications for stratigraphic trap plays. Bas. Res. 29, 816–835 (2017).
Flint, S. S. et al. Depositional architecture and sequence stratigraphy of the Karoo basin floor to shelf edge succession, Laingsburg depocenter, South Africa. Mar. Pet. Geol. 28, 658–674 (2011).
Di Celma, C. N., Brunt, R. L., Hodgson, D. M., Flint, S. S. & Kavanagh, J. P. Spatial and temporal evolution of a Permian submarine slope channel–levee system, Karoo Basin, South Africa. J. Sed. Res. 81, 579–599 (2011).
Figueiredo, J. J., Hodgson, D. M., Flint, S. S. & Kavanagh, J. P. Depositional environments and sequence stratigraphy of an exhumed Permian mudstone-dominated submarine slope succession, Karoo Basin, South Africa. J. Sed. Res. 80, 97–118 (2010).
Prélat, A. & Hodgson, D. M. The full range of turbidite bed thickness patterns in submarine lobes: controls and implications. J. Geol. Soc. London 170, 209–214 (2013).
Hodgson, D. M., Kane, I. A., Flint, S. S., Brunt, R. L. & Ortiz-Karpf, A. Time-transgressive confinement on the slope and the progradation of basin-floor fans: Implications for the sequence stratigraphy of deep-water deposits. J. Sed. Res. 86, 73–86 (2016).
Levin, L. A. & Gage, J. D. Relationships between oxygen, organic matter and the diversity of the bathyal macrofauna. Deep-Sea Res. II 45, 129–163 (1998).
Gage, J. D. & Tyler, P. A. Deep-sea biology: A natural history of organisms at the deep-sea floor. Cambridge, UK: Cambridge University Press (1991).
Glud, R. N., Gundersen, J. K., Jørgensen, B. B., Revsbech, N. P. & Schulz, H. D. Diffusive and total oxygen uptake of deep-sea sediments in the eastern South Atlantic Ocean: in situ and laboratory measurements. Deep-Sea Res. I 41, 1767–1788 (1994).
Piepenburg, D. et al. Partitioning of benthic community respiration in the Arctic (northwestern Barents Sea). Mar. Ecol. Prog. Ser. 118, 199–213 (1995).
Gage, J. D., Hughes, D. J. & Gonzalez Vecino, J. L. Sieve size influence in estimating biomass, abundance and diversity in samples of deep-sea macrobenthos. Mar. Ecol. Prog. Ser. 225, 97–107 (2002).
Levin, L. A., Huggett, C. L. & Wishner, K. F. Control of deep-sea benthic community structure by oxygen and organic-matter gradients in the eastern Pacific Ocean. J. Mar. Res. 49, 763–800 (1991).
Acknowledgements
This research was funded by a Natural Environment Research Council PhD. studentship (Ref. number NE/J016950/1) to S.L.C. with Statoil as a C.A.S.E. partner whilst at Leeds. The authors thank local landowners for their permissions to carry out fieldwork and Colleen Kurcinka for her assistance in the field. Additional background material and data are included in supplementary material.
Author information
Authors and Affiliations
Contributions
S.L. Cobain wrote the main manuscript text and prepared the figures, M.R.D. Cobain designed the model for oxygen consumption, all authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cobain, S.L., Hodgson, D.M., Peakall, J. et al. A new macrofaunal limit in the deep biosphere revealed by extreme burrow depths in ancient sediments. Sci Rep 8, 261 (2018). https://doi.org/10.1038/s41598-017-18481-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-18481-w
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
