
Abstract
Neuroticism is one of the “Big Five” personality factors and is characterized by a tendency to experience negative affect. We aimed to investigate how neuroticism influences the neural correlates for processing of emotional facial expressions. 68 healthy participants were presented with emotional dynamic facial stimuli, i.e. happy, neutral or angry, during functional MRI. Brain activations for the contrasts emotional vs. neutral, happy vs. neutral and angry vs. neutral were correlated with individuals’ neuroticism scores as obtained by the NEO Five Factor Inventory questionnaire and additionally investigated for gender differences. The bilateral medial temporal gyrus (MTG) was identified as key region in the processing of emotional faces and activations within this region correlated with individual neuroticism scores. Although female participants showed significantly stronger activation differences between emotional and neutral facial expressions in the left MTG, the correlation between activation and neuroticism scores did not show any significant gender differences. Our results offer for the first time a biological correlate within the face processing network for enhanced reactivity of neurotic individuals to emotional facial expressions which occurs similarly for both male and female participants.
Similar content being viewed by others


Introduction
According to the so-called “Big Five” personality model, the human personality can be described by five different personality traits: Neuroticism, Extraversion, Openness, Agreeableness, and Conscientiousness1. Of special interest in clinical neuroscience is the factor neuroticism due to its association with certain psychiatric diseases such as anxiety disorders, depression, and substance use disorders2,3. Neuroticism is characterized by the tendency to experience negative affect or dissatisfaction, to worry, and to experience anxiousness4,5,6. Several neuroimaging studies have been performed to investigate the neural correlates of neuroticism using different stimulus categories. Correlations between neuroticism scores and brain activation have been shown for the amygdala, the middle temporal gyrus (MTG), the middle frontal gyrus (MFG) and the dorsomedial prefrontal cortex (DMPFC) using emotional pictures as stimuli6,7,8,9,10 and for the amygdala, the MTG, temporal pole, left MFG, DMPFC, bilateral parietal cortex, visual cortex, and the fusiform gyrus using static emotional faces as stimuli11,12,13,14,15,16,17. Using a prisoner dilemma task, Feng, et al.18 were able to correlate neuroticism with the neural response to negative and positive social interaction in the anterior cingulate cortex/medial prefrontal cortex and the insula, respectively.
Correct interpretation of facial expressions is a prerequisite for effective social interaction. Therefore, face perception has been the subject of numerous neuroimaging studies in the past years, leading to the discovery of a distributed face network of which the occipital face area (OFA), the fusiform face area (FFA), and the posterior superior temporal sulcus (pSTS) are the core areas19,20,21,22. While the OFA and the FFA are hypothesized to be involved mainly in the processing of invariant facial aspects, such as identity or gender20,23, the cortex adjacent to the pSTS is thought to process changeable aspects like emotional facial expressions20,24,25.
Bridging the gap between basic functions underlying social interaction and individual personality traits, we aimed to identify the neural correlates of neuroticism with regards to the processing of emotional facial expressions using functional MRI in a large cohort of healthy participants. Subjects were presented with video clips of emotional face stimuli, i.e. happy or angry expressions, as such stimuli have a higher ecological validity than static images of faces. Furthermore, it has been shown that face-selective regions can be more robustly identified using dynamic compared to static stimuli26,27,28. To identify brain areas associated with certain personality traits, especially neuroticism, activations were correlated with participants’ personality scores as obtained by the NEO Five Factor Inventory questionnaire29. We hypothesized that activity of areas supporting processing of dynamic facial information, i.e. the cortical areas adjacent to the pSTS, is correlated with neuroticism. Finally, electrophysiological studies using electroencephalography30 or magnetencephalography31 as well as neuroimaging studies point to gender differences in face processing32,33. Therefore, we also examined whether relationships between neuroticism and activation to emotional faces are different or similar in male and female participants.
Results
Behavioral data
Intercorrelations between the five personality factors are presented in Table 1. The only intercorrelation which was significant after Bonferroni correction was a negative correlation between neuroticism and extraversion (r = 0.59, p < 0.001). Concerning gender differences, women scored significantly higher on neuroticism (20.7 ± 1.41 versus 16.7 ± 1.41, T(66) = −2.03, two-tailed p < 0.05) and conscientiousness (35.4 ± 0.96 versus 31.8 ± 1.32, T(66) = −2.23, two-tailed p < 0.05) than men. The task of the participants was to identify the gender of the presented face stimuli. They responded in 99.9% of the trials (mean reaction time = 1.98 ± 0.02 s) and correctly identified the gender of the presented actors in 96.0 ± 0.7% of the trials indicating good attention to the presented stimulus material throughout fMRI scanning. There were no significant differences in hit rates or reaction time between female and male participants (both T(66) < 0.70, both two-tailed p > 0.39).
fMRI activation
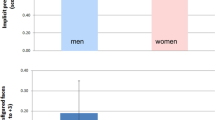
The first step of the fMRI data analysis aimed to identify brain areas involved in processing facial emotions. To this end, we compared activation to video clips displaying emotional versus neutral facial expressions which yielded significant differences (voxel-wise corrected height threshold: p < 0.05, see methods) in bilateral MTG (see Table 2 and Fig. 1). This effect was due to stronger activation to both happy and angry as compared to neutral facial expressions (see Fig. 1, upper panels). A direct gender comparison revealed that in the left MTG, female participants exhibited a larger difference for emotional versus neutral stimuli than male participants (T(66) = −2.74, two-tailed p < 0.01). In the right MTG, no significant difference between male and female participants was found (T(66) = −1.00, two-tailed p = 0.32).

Brain activation to video clips displaying significantly stronger activation (p < 0.05, FWE corrected at voxel level) to emotional versus neutral facial expressions are rendered on the surface of the standard SPM brain template as well as on a transversal slice (z = 6) of the mean normalized brain of the study participants. Activations (beta estimates) ± standard error to happy (positive), neutral, and angry (negative) facial expressions are shown in the upper panels. Correlation analyses between activation differences to emotional versus neutral facial expressions in right and left MTG and neuroticism are shown in the lower panels.
In the second step, we tested whether activation in the areas defined by the contrast emotional versus neutral facial expressions correlated with neuroticism scores. A significant correlation was found for both the cluster in right (r = 0.36, 95% confidence interval: 0.13 < r < 0.55, p = 0.003) and left (r = 0.42, 95% confidence interval: 0.20 < r < 0.60; p < 0.001) MTG (see Fig. 1, lower panels) which survived Bonferroni correction for ten statistical tests (two regions x five personality factors). No significant correlation was found for any of the other four personality factors (see Table 3).
In the third step, we investigated potential effects of the participants’ gender and valence of the stimuli (i.e. positive versus negative emotional information). Separate analysis revealed significant correlations in bilateral MTG both for female (right MTG: r = 0.30, 95% confidence interval: 0.02 < r < 0.55, p = 0.040; left MTG: r = 0.37, 95% confidence interval: 0.08< r < 0.59, p = 0.12) and male (right MTG: r = 0.47, 95% confidence interval: 0.06 < r < 0.74, p = 0.028; left MTG: r = 0.43, 95% confidence interval: 0.01 < r < 0.72, p = 0.046) participants with no significant differences between groups. Valence specific correlation analysis revealed significant correlations between neuroticism and activation of the right MTG to positive versus neutral stimuli (r = 0.26, 95% confidence interval: 0.02 < r < 0.47, p = 0.031) and negative versus neutral stimuli (r = 0.36, 95% confidence interval: 0.11 < r < 0.54, p = 0.003) without significant differences between conditions (p = 0.18). In the left MTG, a significant correlation was found for activation of negative versus neutral stimuli (r = 0.47, 95% confidence interval: 0.22 < r < 0.61, p < 0.001), which marginally failed to reach significance for positive versus neutral stimuli (r = 0.22, 95% confidence interval: −0.02 < r < 0.43, p = 0.07). The difference between the two conditions was significant (p = 0.02) for the left MTG.
In the fourth step, we examined whether removal of variance correlating with the other four personality factors by means of simple regression impacts the observed results obtained for neuroticism. Removal of variance correlating with extraversion reduced the correlation coefficients for the right (r = 0.25, 95% confidence interval: 0.01 < r < 0.46, p = 0.037) and left MTG (r = 0.29, 95% confidence interval: 0.05 < r < 0.49, p = 0.016). Openness, agreeableness, and conscientiousness had no considerable effect on correlation between activation to emotional versus neutral facial expressions and neuroticism which ranged between r = 0.36 and r = 0.37 in the right MTG and r = 0.42 and r = 0.44 in the left MTG (all p < 0.003). Similarly, the correlation coefficients remained significant after removing the conjoint effect of the other four personality factors by means of multiple regression analysis (right MTG: r = 0.38, p = 0.002, left MTG: r = 0.49, p < 0.001).
Finally, we conducted whole-brain correlation analyses between brain activation to emotional versus neutral facial expressions and the five personality factors. This exploratory approach was carried out on a less conservative threshold (voxel-wise uncorrected height threshold p < 0.001, correction for multiple comparison at cluster level, see methods) to make these results accessible to future meta-analyses on associations between personality factors and brain activity. This revealed a significant correlation between neuroticism scores and activation to emotional versus neutral facial expressions in bilateral temporoparietal junction (TPJ) including superior temporal gyrus and supramarginal gyrus as well as right MTG (see Table 4 and Fig. 2). In left MTG, this analysis failed to reach significance at cluster level (peak MNI coordinates: x = −42, y = −63, z = 6; Z = 3.99; k = 41 voxels). For agreeableness, a significant correlation with activation to emotional versus neutral facial expressions was found in left medial prefrontal cortex (MNI coordinates: x = −15, y = 46, z = 6; Z = 3.79; k = 96 voxels, p < 0.05, corrected). No significant correlations were found for any of the other three personality factors.

Correlation between activation to emotional versus neutral facial expressions and neuroticism scores revealed significant correlations in bilateral TPJ including superior temporal gyrus and supramarginal gyrus as well as right MTG (p < 0.05, FWE corrected at cluster level (k > 90 voxels)). Results are rendered on the surface of the standard SPM brain template as well as on a transversal slice (z = 18) of the mean normalized brain of the study participants.
Discussion
In our study, we investigated the impact of neuroticism on processing of dynamic emotional facial stimuli. In agreement with the face model of Haxby and colleagues20, bilateral MTG areas adjacent to the pSTS responded stronger to emotional than to neutral dynamic facial expressions. This enhanced responsiveness to facially displayed emotions correlated significantly with neuroticism scores suggesting this area as key structure underlying increased perception of affect in neurotic individuals.
Here, we decided to define brain areas processing facial emotions by directly contrasting activations to dynamic facial emotional and neutral expressions. This approach yields better emotion specific results than application of an fMRI localizer which compares activation to different classes of stimuli (e.g. faces versus objects) as such comparisons are burdened with differences in a variety of low-level features (e.g. size, shape, contrast) and associated higher-level processes (e.g. faces, but not object reflect individuals, objects, but not faces are associated with typical movements to use them).
Contrasting emotional with neutral dynamic facial expressions resulted in significantly stronger responses in bilateral MTG adjacent to the pSTS. This area is thought to represent the key structure for processing dynamic facial information20,24,25,34 as well as their integration with vocal information35,36. Furthermore, this region has been previously described to react preferentially to emotionally salient stimuli15,37. In our study, this activation difference was more pronounced and yielded a larger cluster in the right than in the left hemisphere which corroborates previous neuroimaging results38. Concerning gender differences, however, the left MTG showed a significantly stronger enhancement of activity in the contrast emotional versus neutral faces in female than in male participants. This probably reflects the fact that the female participants scored higher in neuroticism which is in accordance with the literature39,40,41,42.
Our results showed that activation in bilateral MTG during the processing of emotional faces correlated positively with neuroticism scores, i.e. the higher the participant’s neuroticism score the stronger the activation in bilateral MTG for both male and female participants. This suggests that the personality factor neuroticism and the processing of salient facial stimuli within the MTG are strongly related and that this mechanism occurs irrespective of gender. For negative stimuli, a positive correlation was found for bilateral MTG. It is well known that neuroticism is particularly associated with an enhanced experience of negative affect4,5,6. Therefore, it is not surprising that the relation between processing of salient facial information and neuroticism is more consistently found for negative information. Hyperreactivity to negative stimuli in affective brain regions, including the MTG, in high neurotic individuals has been described previously and has been attributed to increased arousal by emotional conflict and negative affect4,6,43,44,45 or more intensive visual processing of threat-relevant (face) stimuli14. In right MTG, activation to positive stimuli was also correlated with neuroticism indicating that enhanced responsiveness of neurotic individuals to emotional stimuli is not restricted to negative valence. Future studies should investigate whether the effects observed for the emotions happiness and anger occur similarly for other emotions (e.g. sadness, fear, disgust). Furthermore, it should be clarified whether the correlation between neuroticism and responses to emotional stimuli depend on the intensity of the portrayed emotions and whether the effects which we found here for the MTG and faces can be generalized across sensory modalities (i.e., voices or bodies). Corresponding structures for a correlation between neuroticism and responses to emotional voices and bodies would be the temporal voice area46 and extrastriate body area47 which also show a particular sensitivity for affective information48,49. No correlation between amygdala activation and neuroticism scores was found (even at an uncorrected threshold of p < 0.05). The implicit nature of the paradigm employed here might offer an explanation for this finding as a recent resting-state fMRI study50 showed a correlation between neuroticism and connectivity of the amygdala with frontal structures involved in explicit evaluation of emotional faces51, but not with face processing areas in sensory cortices.
In theory, the “Big Five” factors are conceptually independent dimensions. However, we observed a significant negative correlation between neuroticism and extraversion scores in our cohort. Such intercorrelations make it difficult to fully disambiguate the contribution of single personality factors to brain responses and thus represent a limitation of the current study. However, it should be noted that this negative correlation between neuroticism and extraversion does not reflect a phenomenon which is due to the specific group of participants recruited for this study as this intercorrelation has been reported repeatedly15,29,42,52,53. To address this issue, we removed the variance correlating with the other four personality factors in separate simple regression analyses as well as a multiple regression analysis which allows for their combined contribution. In all cases, the correlation between responses in bilateral MTG remained significant indicating that neural activity is modulated primarily by neuroticism irrespective of the other four personality factors.
Our results underline the importance of the bilateral MTG for processing emotional facial stimuli. Critically, correlations of MTG activity with neuroticism scores indicate that neuroticism and neural activity in this region are associated providing a potential neural correlate for increased sensitivity of neurotic individuals to emotional information which occurs similarly in women and men. Thus, we conclude that neuroticism modulates neural activity in MTG during processing of facial emotions. It should be noted that correlations never indicate causality and that the reverse explanation is also possible (i.e. that increased activity within emotion processing areas induces neuroticism). However, given the fact that personality traits are conceptually not restricted to judgment of emotional facial expressions, but cover a much wider range of interpersonal relationships, we deem the first explanation more likely than the second.
Methods
Participants
68 volunteers (46 females, 22 males, mean age: 26.0 ± 4.9 years, age range: 18–36 years completed the German version of the NEO Five Factor Inventory (NEO FFI)29 before functional magnetic resonance imaging (fMRI). The NEO FFI contains 60 items with 12 statements for each of the five personality factors (Neuroticism, Extraversion, Openness, Agreeableness, and Conscientiousness). None of the participants had a history of neurological or psychiatric disorders or was on any psychotropic medication. All participants were right-handed as determined by the Edinburgh Handedness Inventory54 and had normal or corrected to normal vision. The study conformed to the code of Ethics of the World Medical Association (Declaration of Helsinki). The study protocol was approved by the ethics committee of the University of Tübingen and the methods were carried out in accordance with the relevant guidelines and regulations. All participants gave written informed consent prior to participating.
Stimulus material and experimental design
Short video clips (mean duration: 1384 ms ± 264 ms) depicting the faces of professional actors (5 male, 5 female) speaking German sentences consisting of four words (e.g.”Ich fühle mich ruhig” translation: “I feel calm”) in happy, neutral, or angry tone of voice with a corresponding facial expression were used as stimulus material in an event-related design (for a detailed description of the stimuli, see55. These stimuli (20 per emotional category) were presented without auditory stimulation (mute video clips) and the participants were engaged in a gender decision task to keep the participants attentive of the stimuli during scanning. We employed an implicit emotion perception paradigm as this more closely resembles natural situations than paradigms that include explicit instructions to judge emotions. Responses were conveyed via button press via a fiber optic system and recorded by the stimulation software (Software Presentation, Neurobehavioral Systems). Responses had no influence on stimulus presentation and no feedback regarding whether the responses were correct or not was given during the experiment. A black fixation cross was presented during an inter trial interval of varying duration ranging from 8.5 to 15 s (jittered in steps of TR/4).
Image acquisition
All participants were examined in a 3 Tesla PRISMA scanner (Siemens, Erlangen, Germany) using a 20 channel head coil. Functional imaging data comprised 360 volumes acquired with an echo-planar-imaging sequence covering the whole cerebral cortex (30 axial slices acquired in sequential descending order, slice thickness 4 mm + 1 mm gap, TR = 2 s, TE = 30 ms, voxel size = 3 × 3 × 4 mm, flip angle = 90°). For distortion correction, a fieldmap (TR = 400 ms, TEs = 4.92 and 7.38 ms, slice thickness = 3 mm, flip angle = 60°) was acquired prior to each functional run. A high-resolution T1-weighted anatomical 3D magnetization prepared rapid acquisition gradient echo (MPRAGE, FOV = 256 × 256 mm, 1 mm isotropic voxel size, TR = 2.3 s, TE = 4.18 ms, TI = 900 ms, flip angle = 9°) data set was obtained to enable precise normalization of the functional scans.
Analysis of questionnaires and behavioral data
Behavioral data are reported as mean ± standard error. Questionnaire data obtained from the NEO FFI were examined for intercorrelations (Pearson’s r) between the five personality factors. To allow for multiple comparisons a Bonferroni correction for ten intercorrelations between the personality factors was employed. Behavioral data (hit rates and reaction times) of the fMRI experiment are reported as mean ± standard error. Responses were classified as hits if the gender was correctly classified within 5 seconds after stimulus onset. Correlations between fMRI activations and personality scores are reported including 95% confidence intervals.
Analysis of fMRI data
The images were analyzed using statistical parametric mapping software (SPM12, Wellcome Department of Imaging Neuroscience, London, UK). The first five functional volumes were discarded to allow for T1 equilibrium. Preprocessing included correction for differences in slice acquisition time with the middle slice as reference, realignment and unwarping to correct for movement as well as static and movement dependent field distortions56, coregistration to the high-resolution anatomical image, normalization57 to Montreal Neurological Institute (MNI) space (resampled voxel size: 3 × 3 × 3 mm), and spatial smoothing with an isotropic Gaussian filter of 8 mm full width at half maximum. Event-related responses were analyzed using a general linear model with three regressors for videos displaying happy, angry, and neutral emotional expressions using a stick function convolved with the hemodynamic response function. The six movement parameters were included as covariates of no interest. Low frequency components were filtered out by applying a high-pass-filter with cut-off-frequency of 1/128 Hz. The error term was modeled as a first-order autoregressive process (AR coefficient = 0.2) plus white noise to account for serial autocorrelations within the data. The contrasts of emotional (happy and angry) versus neutral facial expressions obtained at first-level for each participant were used in a second-level random-effects analysis (one-sample t-test). Statistical significance was defined using a voxel-wise family-wise error (FWE) corrected height threshold of p < 0.05 and an extent threshold of k > 10 voxels. Assignment of brain structures to activated areas was based on the automated anatomical labelling (AAL) atlas58. To examine the impact of personality factors, correlation analyses between responses to emotional versus neutral stimuli in significantly activated brain regions (i.e. beta estimates across all voxels of activated clusters) and the five personality factors as obtained by the NEO-FFI were carried out (two-sided tests). Analogous analyses were carried out for the contrasts happy versus neutral and angry versus neutral to examine whether correlations found for the contrast emotional versus neutral are driven by one of the two conditions or occur similarly for both conditions. To allow for multiple comparisons a Bonferroni correction for the number of examined regions x five personality factors was employed.
Significant Pearson’s r correlation coefficients were further examined for effects of gender of the study participants and differential contributions of positive and negative emotions. To this end, we performed separate correlation analyses for female and male participants as well as emotional category (i.e. correlation between neuroticism and activation to positive versus neutral and negative versus neutral facial expressions). Resulting correlation coefficients were tested for significant differences between groups and conditions using t-tests. Finally, we tested whether correlation coefficients between brain activity and one of the five personality factor remain significant if the effects of the other four personality factors are regressed out. This was done using both simple regression analysis for each of the other four personality factors in isolation as well as multiple regression analysis to allow for potential conjoint effects of the four other personality factors.
Finally, we conducted exploratory whole-brain correlation analyses between brain activity to emotional versus neutral facial expressions and the five personality factors assessed by the NEO-FFI to enable inclusion of these data in future meta-analyses. The results of these correlation analyses are reported at a height threshold of p < 0.001 (uncorrected). Correction for multiple comparisons was carried out at cluster level (k > 90 voxels) corresponding to p < 0.05, corrected.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
McCrae, R. R. & John, O. P. An introduction to the five-factor model and its applications. Journal of personality 60, 175–215 (1992).
Kotov, R., Gamez, W., Schmidt, F. & Watson, D. Linking “big” personality traits to anxiety, depressive, and substance use disorders: a meta-analysis. Psychol Bull 136, 768–821, https://doi.org/10.1037/a0020327 (2010).
Lahey, B. B. Public health significance of neuroticism. The American psychologist 64, 241–256, https://doi.org/10.1037/a0015309 (2009).
Ormel, J. et al. The biological and psychological basis of neuroticism: current status and future directions. Neuroscience and biobehavioral reviews 37, 59–72, https://doi.org/10.1016/j.neubiorev.2012.09.004 (2013).
Costa, P. T. Jr & McCrae, R. R. Influence of extraversion and neuroticism on subjective well-being: happy and unhappy people. Journal of personality and social psychology 38, 668–678 (1980).
Canli, T. et al. An fMRI study of personality influences on brain reactivity to emotional stimuli. Behavioral neuroscience 115, 33–42 (2001).
Britton, J. C., Ho, S. H., Taylor, S. F. & Liberzon, I. Neuroticism associated with neural activation patterns to positive stimuli. Psychiatry research 156, 263–267, https://doi.org/10.1016/j.pscychresns.2007.06.003 (2007).
Cunningham, W. A., Arbuckle, N. L., Jahn, A., Mowrer, S. M. & Abduljalil, A. M. Reprint of: Aspects of neuroticism and the amygdala: chronic tuning from motivational styles. Neuropsychologia 49, 657–662, https://doi.org/10.1016/j.neuropsychologia.2011.02.027 (2011).
Harenski, C. L., Kim, S. H. & Hamann, S. Neuroticism and psychopathy predict brain activation during moral and nonmoral emotion regulation. Cognitive, affective & behavioral neuroscience 9, 1–15, https://doi.org/10.3758/CABN.9.1.1 (2009).
Kehoe, E. G., Toomey, J. M., Balsters, J. H. & Bokde, A. L. Personality modulates the effects of emotional arousal and valence on brain activation. Social cognitive and affective neuroscience 7, 858–870, https://doi.org/10.1093/scan/nsr059 (2012).
Madsen, M. K. et al. Threat-related amygdala functional connectivity is associated with 5-HTTLPR genotype and neuroticism. Social cognitive and affective neuroscience 11, 140–149, https://doi.org/10.1093/scan/nsv098 (2016).
Hyde, L. W., Gorka, A., Manuck, S. B. & Hariri, A. R. Perceived social support moderates the link between threat-related amygdala reactivity and trait anxiety. Neuropsychologia 49, 651–656, https://doi.org/10.1016/j.neuropsychologia.2010.08.025 (2011).
Haas, B. W., Constable, R. T. & Canli, T. Stop the sadness: Neuroticism is associated with sustained medial prefrontal cortex response to emotional facial expressions. NeuroImage 42, 385–392, https://doi.org/10.1016/j.neuroimage.2008.04.027 (2008).
Chan, S. W., Norbury, R., Goodwin, G. M. & Harmer, C. J. Risk for depression and neural responses to fearful facial expressions of emotion. The British journal of psychiatry: the journal of mental science 194, 139–145, https://doi.org/10.1192/bjp.bp.107.047993 (2009).
Cremers, H. R. et al. Neuroticism modulates amygdala-prefrontal connectivity in response to negative emotional facial expressions. NeuroImage 49, 963–970, https://doi.org/10.1016/j.neuroimage.2009.08.023 (2010).
Everaerd, D., Klumpers, F., van Wingen, G., Tendolkar, I. & Fernandez, G. Association between neuroticism and amygdala responsivity emerges under stressful conditions. NeuroImage 112, 218–224, https://doi.org/10.1016/j.neuroimage.2015.03.014 (2015).
Jimura, K., Konishi, S. & Miyashita, Y. Temporal pole activity during perception of sad faces, but not happy faces, correlates with neuroticism trait. Neuroscience letters 453, 45–48, https://doi.org/10.1016/j.neulet.2009.02.012 (2009).
Feng, C., DeMarco, A. C., Haroon, E. & Rilling, J. K. Neuroticism modulates the effects of intranasal vasopressin treatment on the neural response to positive and negative social interactions. Neuropsychologia 73, 108–115, https://doi.org/10.1016/j.neuropsychologia.2015.05.004 (2015).
Gobbini, M. I. & Haxby, J. V. Neural systems for recognition of familiar faces. Neuropsychologia 45, 32–41, https://doi.org/10.1016/j.neuropsychologia.2006.04.015 (2007).
Haxby, J. V., Hoffman, E. A. & Gobbini, M. I. The distributed human neural system for face perception. Trends in cognitive sciences 4, 223–233 (2000).
Ishai, A. Let’s face it: it’s a cortical network. NeuroImage 40, 415–419, https://doi.org/10.1016/j.neuroimage.2007.10.040 (2008).
Pitcher, D., Walsh, V. & Duchaine, B. The role of the occipital face area in the cortical face perception network. Experimental brain research 209, 481–493, https://doi.org/10.1007/s00221-011-2579-1 (2011).
Kanwisher, N., McDermott, J. & Chun, M. M. The fusiform face area: a module in human extrastriate cortex specialized for face perception. The Journal of neuroscience: the official journal of the Society for Neuroscience 17, 4302–4311 (1997).
Allison, T., Puce, A. & McCarthy, G. Social perception from visual cues: role of the STS region. Trends in cognitive sciences 4, 267–278 (2000).
Engell, A. D. & Haxby, J. V. Facial expression and gaze-direction in human superior temporal sulcus. Neuropsychologia 45, 3234–3241, https://doi.org/10.1016/j.neuropsychologia.2007.06.022 (2007).
Pitcher, D., Dilks, D. D., Saxe, R. R., Triantafyllou, C. & Kanwisher, N. Differential selectivity for dynamic versus static information in face-selective cortical regions. Neuroimage 56, 2356–2363, https://doi.org/10.1016/j.neuroimage.2011.03.067 (2011).
Hasson, U., Malach, R. & Heeger, D. J. Reliability of cortical activity during natural stimulation. Trends in cognitive sciences 14, 40–48, https://doi.org/10.1016/j.tics.2009.10.011 (2010).
Scherf, K. S., Luna, B., Minshew, N. & Behrmann, M. Location, Location, Location: Alterations in the Functional Topography of Face- but not Object- or Place-Related Cortex in Adolescents withAutism. Frontiers in human neuroscience 4, 26, https://doi.org/10.3389/fnhum.2010.00026 (2010).
McCrae, R. R. & Costa, P. T. A contemplated revision of the NEO Five-Factor Inventory. Pers Individ Dif 36, 587–596 (2004).
Choi, D. et al. Gender difference in N170 elicited under oddball task. J Physiol Anthropol 34, 7 (2015).
Tiedt, H. O., Weber, J. E., Pauls, A., Beier, K. M. & Lueschow, A. Sex-differences of face coding: evidence from larger right hemispheric M170 in men and dipole source modelling. PLoS One 8, e69107, https://doi.org/10.1371/journal.pone.0069107 (2013).
Wager, T. D., Phan, K. L., Liberzon, I. & Taylor, S. F. Valence, gender, and lateralization of functional brain anatomy in emotion: a meta-analysis of findings from neuroimaging. Neuroimage 19, 513–531 (2003).
Weisenbach, S. L. et al. Reduced emotion processing efficiency in healthy males relative to females. Soc Cogn Affect Neurosci 9, 316–325, https://doi.org/10.1093/scan/nss137 (2014).
Ethofer, T., Gschwind, M. & Vuilleumier, P. Processing social aspects of human gaze: a combined fMRI-DTI study. Neuroimage 55, 411–419, https://doi.org/10.1016/j.neuroimage.2010.11.033 (2011).
Kreifelts, B., Ethofer, T., Shiozawa, T., Grodd, W. & Wildgruber, D. Cerebral representation of non-verbal emotional perception: fMRI reveals audiovisual integration area between voice- and face-sensitive regions in the superior temporal sulcus. Neuropsychologia 47, 3059–3066, https://doi.org/10.1016/j.neuropsychologia.2009.07.001 (2009).
Ethofer, T. et al. Functional responses and structural connections of cortical areas for processing faces and voices in the superior temporal sulcus. Neuroimage 76, 45–56, https://doi.org/10.1016/j.neuroimage.2013.02.064 (2013).
Critchley, H. et al. Explicit and implicit neural mechanisms for processing of social information from facial expressions: a functional magnetic resonance imaging study. Human brain mapping 9, 93–105 (2000).
De Winter, F. L. et al. Lateralization for dynamic facial expressions in human superior temporal sulcus. NeuroImage 106, 340–352, https://doi.org/10.1016/j.neuroimage.2014.11.020 (2015).
Costa, P. T. Jr, Terracciano, A. & McCrae, R. R. Gender differences in personality traits across cultures: robust and surprising findings. Journal of personality and social psychology 81, 322–331 (2001).
Chapman, B. P., Duberstein, P. R., Sorensen, S. & Lyness, J. M. Gender Differences in Five Factor Model Personality Traits in an Elderly Cohort: Extension of Robust and Surprising Findings to an Older Generation. Pers Individ Dif 43, 1594–1603, https://doi.org/10.1016/j.paid.2007.04.028 (2007).
Weisberg, Y. J., Deyoung, C. G. & Hirsh, J. B. Gender Differences in Personality across the Ten Aspects of the Big Five. Frontiers in psychology 2, 178, https://doi.org/10.3389/fpsyg.2011.00178 (2011).
Nostro, A. D., Muller, V. I., Reid, A. T. & Eickhoff, S. B. Correlations Between Personality and Brain Structure: A Crucial Role of Gender. Cerebral cortex, https://doi.org/10.1093/cercor/bhw191 (2016).
Chan, S. W., Harmer, C. J., Goodwin, G. M. & Norbury, R. Risk for depression is associated with neural biases in emotional categorisation. Neuropsychologia 46, 2896–2903, https://doi.org/10.1016/j.neuropsychologia.2008.05.030 (2008).
Haas, B. W., Omura, K., Constable, R. T. & Canli, T. Emotional conflict and neuroticism: personality-dependent activation in the amygdala and subgenual anterior cingulate. Behavioral neuroscience 121, 249–256, https://doi.org/10.1037/0735-7044.121.2.249 (2007).
Hooker, C. I., Verosky, S. C., Miyakawa, A., Knight, R. T. & D’Esposito, M. The influence of personality on neural mechanisms of observational fear and reward learning. Neuropsychologia 46, 2709–2724, https://doi.org/10.1016/j.neuropsychologia.2008.05.005 (2008).
Belin, P., Zatorre, R. J., Lafaille, P., Ahad, P. & Pike, B. Voice-selective areas in human auditory cortex. Nature 403, 309–312, https://doi.org/10.1038/35002078 (2000).
Downing, P. E., Jiang, Y., Shuman, M. & Kanwisher, N. A cortical area selective for visual processing of the human body. Science 293, 2470–2473, https://doi.org/10.1126/science.1063414 (2001).
Ethofer, T. et al. Emotional voice areas: anatomic location, functional properties, and structural connections revealed by combined fMRI/DTI. Cereb Cortex 22, 191–200, https://doi.org/10.1093/cercor/bhr113 (2012).
Peelen, M. V., Atkinson, A. P., Andersson, F. & Vuilleumier, P. Emotional modulation of body-selective visual areas. Soc Cogn Affect Neurosci 2, 274–283, https://doi.org/10.1093/scan/nsm023 (2007).
Pang, Y. et al. Extraversion and neuroticism related to the resting-state effective connectivity of amygdala. Sci Rep 6, 35484, https://doi.org/10.1038/srep35484 (2016).
Kitada, R., Johnsrude, I. S., Kochiyama, T. & Lederman, S. J. Brain networks involved in haptic and visual identification of facial expressions of emotion: an fMRI study. Neuroimage 49, 1677–1689, https://doi.org/10.1016/j.neuroimage.2009.09.014 (2010).
Egan, V., Deary, I. & Austin, E. The NEO-FFI: emerging British norms and an item-level analysis suggest N, A and C are norms reliable than O and E. Pers Indiv Differ 29, 907–920, https://doi.org/10.1016/S0191-8869(99)00242-1 (2000).
van der Linden, D., Nijenhuis, J. T. & Bakker, A. B. The General Factor of Personality: A meta-analysis of Big Five intercorrelations and a criterion-related validity study. J Res Pers 44, 315–327, https://doi.org/10.1016/j.jrp.2010.03.003 (2010).
Oldfield, R. C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113 (1971).
Jacob, H. et al. Cerebral integration of verbal and nonverbal emotional cues: impact of individual nonverbal dominance. NeuroImage 61, 738–747, https://doi.org/10.1016/j.neuroimage.2012.03.085 (2012).
Andersson, J., Hutton, C., Ashburner, J., Turner, R. & Friston, K. Modeling geometric deformations in EPI time series. NeuroImage 13, 903–912 (2001).
Ashburner, J. & Friston, K. J. Unified segmentation. NeuroImage 26, 839–851, https://doi.org/10.1016/j.neuroimage.2005.02.018 (2005).
Tzourio-Mazoyer, N. et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 15, 273–289, https://doi.org/10.1006/nimg.2001.0978 S1053811901909784 (2002).
Acknowledgements
This work was supported by the Deutsche Forschungsgemeinschaft (DFG grants ET112/5-1 and ET 112/6-1). We acknowledge funding of the Open Access funds of the Deutsche Forschungsgemeinschaft. The funding source had no role in study design, the collection, analysis and interpretation of data, the writing of the manuscript and the decision to submit the article for publication.
Author information
Authors and Affiliations
Contributions
T.E. and L.S. designed the study. L.S. and O.K. acquired the data. S.K. and T.E. analyzed the data. S.K. wrote the first draft of the manuscript. All authors participated in the preparation, modification and revision of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Klamer, S., Schwarz, L., Krüger, O. et al. Association between Neuroticism and Emotional Face Processing. Sci Rep 7, 17669 (2017). https://doi.org/10.1038/s41598-017-17706-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-17706-2
This article is cited by
-
Atypical neural activation associated with implicit negative emotional facial processing in fMRI tasks in individuals with neuroticism personality traits
Current Psychology (2023)
-
Lower cognitive reappraisal capacity is related to impairments in attachment and personality structure in poly-drug use: an fMRI study
Brain Imaging and Behavior (2021)
-
Neuroticism predicts the impact of serotonin challenges on fear processing in subgenual anterior cingulate cortex
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
