
Abstract
Here we provide empirical evidence of the presence of an energetic pathway between jellyfish and a commercially important invertebrate species. Evidence of scavenging on jellyfish carcasses by the Norway lobster (Nephrops norvegicus) was captured during two deployments of an underwater camera system to 250–287 m depth in Sognefjorden, western Norway. The camera system was baited with two Periphylla periphylla (Scyphozoa) carcasses to simulate the transport of jellyfish detritus to the seafloor, hereby known as jelly-falls. N. norveigus rapidly located and consumed a large proportion (>50%) of the bait. We estimate that the energy input from jelly-falls may represent a significant contribution to N. norvegicus energy demand (0.21 to 10.7 times the energy required for the population of N. norvegicus in Sognefjorden). This potentially high energetic contribution from jelly-falls highlights a possible role of gelatinous material in the support of commercial fisheries. Such an energetic pathway between jelly-falls and N. norvegicus could become more important with increases in jellyfish blooms in some regions.
Similar content being viewed by others
Introduction
Carrion that sinks to the seafloor represents a major energy transfer in the marine environment1. Carrion is detected, located and consumed by marine scavengers2, enabling the transfer of nutrients and energy back into pelagic and benthic marine food webs3. The aggregation of jellyfish carcasses on the seafloor (jelly-falls) are also a source of organic matter input to the benthos4,5,6. Recently, increased frequencies of jellyfish blooms have been observed in a number of regions around the world, including in Norwegian fjords, where Periphylla periphylla is now highly abundant7,8. A growing body of evidence suggests that the role of gelatinous zooplankton in the biological carbon pump in jellyfish-dominated fjords may be significant, and jelly-falls are actively scavenged here. For example, rapid scavenging on jelly-falls was first shown in Sognefjorden in 20129, with scavenging on jelly-falls being dominated by Atlantic hagfish (Myxine glutinous), galatheid crabs (Munida sp.) and decapod shrimp (Pandalus borealis).
The Norway lobster (Nephrops norvegicus) is an economically important commercial species in the Atlantic10, and is common in Norwegian coastal regions11. In Norway, the revenue from the N. norvegicus fishery exceeded 3 million US dollars in 201512. Although the species has been shown to predate and scavenge on a wide range of marine species, primarily decapod crustaceans and fish13,14, it has never been observed scavenging on jelly-falls. Here, we describe the first photo-documentation of N. norvegicus rapidly scavenging on jellyfish carcasses, and provide empirical evidence of the presence of an energetic pathway between jellyfish populations and this commercially-important species. The role of jelly-falls in the energy budget of N. norvegicus is also estimated using values for energy input from jelly-falls and N. norvegicus energy demand.
Material and Methods
Two baited underwater camera (BUC) deployments were made at 250 and 287 m across the outer fjord sill (061° 04.476′ N, 004° 59. 236′ E and 061° 04.087′ N, 005° 00. 378′ E respectively) of Sognefjorden, western Norway in June 2016. Both deployments lasted approximately 10 hours and were conducted during the day (09:10 to 19:07 UTC) and the night (22:25 to 08:02 UTC) respectively. Photographic still images of the jellyfish carcass and attending fauna were taken every 2 minutes by a deep-sea digital single lens reflex camera (Ocean Imaging Systems DSC 24000) system positioned 1.5 meters directly above a square bait plate (0.5 m2). For each BUC deployment, the bait plate was baited with two defrosted P. periphylla carcasses (~266 g ± 26, mean ± range). The number of scavengers from each species at the bait, the maximum number of scavengers observed at the bait at a single time (MaxN, a proxy for scavenger abundance) and the time to first scavenger arrival (tarrival) were recorded from photographic images from each deployment. The flux of jellyfish material from the water column as jelly-falls (kJ m−2 d−1) to the seafloor was estimated using mass input data and bomb-calorimetry analysis described in previous studies5,9. Jellyfish carrion flux rates were compared to N. norvegicus daily energy intake rates (kJ d−1) using daily food intake data from a previous study that was adjusted for temperature using Q10 14. Also, estimates of the energy content of P. periphylla tissue that were attached to each BUC (kJ g dry weight d−1) were calculated based on the mass of jellyfish and bomb calorimetry analysis from9.
Data on the density of N. norvegicus on the seafloor is required to determine the contribution of jelly-falls to their energy demand. Previous BUC studies have calculated the density of scavengers using the tarrival method15. This model works well with abyssal tarrival data sets, where tarrival is generally longer (e.g. >100 minutes) than for datasets collected from shallower depth zones, as highlighted in a previous study16. Therefore, N. norvegicus seafloor densities in Sognefjorden were based on minimum and maximum N. norvegicus densities from other boreal coastal/fjord environments. The data used came from earlier studies in the Firth of Clyde (0.10 to 0.55 individuals m−2, 17), Scottish sea lochs (Loch Torridon, (0.13 individuals m−2, 18) and Loch Aline (0.18 individuals m−2, 19)), the Irish Sea (0.13 to 0.31 individuals m−2)20), the Kattegat and Skagerrak (0.2 to 0.4 individuals m−2, 11) and the North Sea (0.09 to 0.73 individuals m−2)21.
Data Availability Statement
No restrictions exist on the availability of material and data.
Results and Discussion
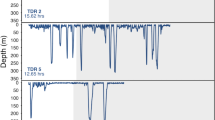
A number of different scavenger species consumed the bait in both BUC deployments. Hagfish (Myxine glutinosa) always arrived at the bait first (tarrival = 2 minutes and 14 minutes). Other scavengers, such as M. glutinosa, P. borealis and Munida sp., also consumed the bait, but often declined in abundance when N. norvegicus was present (Fig. 1a and b). A maximum of 1 N. norvegicus arrived and fed at the bait in deployment 1, while a maximum of 2 N. norvegicus were observed in deployment 2 (Fig. 2), with the first N. norvegicus arriving 24 (first deployment) and 18 minutes (second deployment) after the lander reached the seafloor. In both deployments N. norvegicus consumed a large proportion (>50%) of the bait. N. norvegicus first removed what remained of the nutritional gonad tissue, and then continued to feed on the remaining mesoglea tissue. N. norvegicus feed during day-light and night hours. It was not possible to detect any influence of time of day on the abundance of N. norvegicus (Fig. 1) owing to the low abundance of animals observed, yet this species has been observed to display diurnal patterns of emergence at the depths where we photographed it22,23.

Maximum number of Nephrops norvegicus and other scavengers (inc. Myxine glutinosa, Munida sp. and Pandalus borealis) observed in the BUC deployment at (a) 250 m (arrival time of BUC on seafloor 07:10 UTC; 19 June 2016) and (b) 287 m (arrival time of BUC on seafloor 20:25 UTC; 21 June 2016). Time represents the minutes that elapsed after arrival of the BUC on the seafloor.

(A) Two Nephrops norvegicus feeding and approaching the Periphylla periphylla bait; and (B) an individual N. norvegicus removing and feeding on a large portion of gelatinous mesoglea at the bait plate. White lines are spaced 10 cm apart.
The energy intake rate of a 26 g N. norvegicus in the Firth of Clyde, Scotland, was found to be approximately 1.97 kJ d−1 at a mean temperature of 11 °C14. At an in-situ temperature of 7.7 °C in Sognefjorden, Q10-adjusted energy intake rates would be 1.6 (Q10 = 2) to 1.4 (Q10 = 3) kJ d−1. Therefore, assuming that these energy intake rates are similar to that of the N. norvegicus individuals photographed in our study (N. norvegicus mass of ~29.8 g estimated from length-mass relationships), 50% of the jellyfish bait consumed in our experiments by N. norvegicus (mean energy content of 16.7 kJ g dry weight−1) would provide enough energy for a single N. norvegicus to survive for 90 (Q10 = 2) to 103 days (Q10 = 3).
Despite the relatively low energy content of jellyfish material24, it is known to be an important food source to a variety of marine predators. For example, the leatherback turtle (Dermochelys coriacea) relies upon a diet of low energy-content gelatinous zooplankton25, and salps are an important contribution to the diets of bentho-pelagic fish26. Commercially exploited invertebrates have been recorded in traps baited with the giant jellyfish Nemopilema nomurai 27. However, to the best of our knowledge, this is the first study that has directly photographed N. norvegicus feeding on a gelatinous organism, and attempted to quantify the importance of jelly-falls as an energy resource to this particular commercially-exploited invertebrate species.
Jellyfish are known to dominate several fjords along the Norwegian coast, including in Lurefjorden and Sognefjorden7,8. In Sognefjorden, the abundance of P. periphylla is high (100–300 individuals m−2), and biomasses here are several orders of magnitude higher than those in the open ocean7. This is also true for Lurefjorden, where large pelagic populations of P. periphylla contribute (as jelly-falls) to an efficient jelly-pump that can be as important in transporting C and N as the classic phytodetritus pump5. Therefore, jellyfish carcass flux data from Lurefjorden was used to estimate how important jellyfish carcasses could be to N. norvegicus communities in Sognefjorden. The flux of jelly-fall material transported to the seafloor in Lurefjorden between November 2010 and November 2011 ranged from 12.5 mg to 72.8 mg C m−2 d−1 or 0.5 to 3.0 kJ m−2 d−1. The density of N. norvegicus in similar environments in other regions of Northern Europe ranges from a minimum of 0.10 to 0.73 individuals m−2 17. Therefore, assuming similar seafloor densities for N. norvegicus in Sognefjorden, and similar gelatinous carrion flux rates in both Lurefjorden and Sognefjorden, and a conservative consumption of half of the jellyfish, daily jelly-fall fluxes could provide 0.21 to 10.7 times the daily energy requirement for N. norvegicus in Sognefjorden. This high energetic contribution from jelly-falls to N. norvegicus clearly highlights a potentially important role of gelatinous material in the support of a commercially important species along the Norwegian margin. Even at high N. norvegicus densities (0.73 m−2) and low gelatinous flux rates (0.5 kJ m−2 d−1), jelly-falls potentially still provide almost a quarter of the daily energetic demands of N. norvegicus populations. Although information on N. norvegicus stock sizes have not been collected within fjords, population-size information combined with the type of data presented here may enable the total number of N. norvegicus that can be supported by jellyfish carrion to be calculated. This represents valuable information for fisheries management. Such estimates could be further improved with data on N. norvegicus foraging patterns, scavenging rates, and the contribution of other food sources to their diets.
This work demonstrates that jelly-falls can provide an important source of nutrition to a commercially-important species in Norway, and suggests that energy transfer pathways from jellyfish to benthic species may become more important in regions where jellyfish blooms presently occur, or are becoming more common (e.g. in numerous Norwegian fjords). There is evidence that the role of fish in some pelagic ecosystems may decline, and by inference, the transport of fish carrion to the seafloor, with increasing jellyfish biomass8,28,29,30. Carrion supply influences deep-sea scavenger community dynamics and changes in the amount of fish carrion reaching the seafloor have been linked to the abundance of deep-sea grenadiers31. The findings presented here provide empirical evidence that a loss of energetic resources from fish (and other pelagic animal) carrion to deep-water scavengers could potentially be partially offset by sinking gelatinous material.
References
Payne, L. & Moore, J. Mobile scavengers create hotspots of freshwater productivity. Oikos 115, 69–8 (2006).
Britton, J. C. & Morton, B. Marine carrion and scavengers. Oceanogr. Mar. Biol. Ann. Rev. 32, 369–434 (1994).
King., N. J., Bailey, D. M. & Priede, I. G. Role of scavengers in marine ecosystems: introduction. Mar. Ecol. Prog. Ser. 350, 175–178 (2007).
Sweetman, A. K. & Chapman, A. First observations of jelly-falls at the seafloor in a deep-sea fjord. Deep-Sea Res. 58, 1206–1211 (2011).
Sweetman, A. K. & Chapman, A. First assessment of flux rates of jellyfish carcasses (jelly-falls) to the benthos reveals the importance of gelatinous material for biological C-cycling in jellyfish-dominated ecosystems. Front. Mar. Sci. 47 (2015).
Lebrato, M. et al. Depth attenuation of organic matter export associated with jelly falls. Limnol. Oceanogr. 56, 1917–1928 (2011).
Sørnes, T. A., Aksnes, D. L., Båmstedt, U. & Youngbluth, M. J. Causes for mass occurrences of the jellyfish Periphylla periphylla: a hypothesis that involves optically-conditioned retention. J. Plank. Res. 29, 157–167 (2007).
Aksnes, D. L. et al. Coastal water darkening and implications for mesopelagic regime shifts in Norwegian fjords. Mar. Ecol. Prog. Ser. 387, 39–49 (2009).
Sweetman, A. K., Smith, C. R., Dale, T. & Jones, D. O. B. Rapid scavenging of jellyfish carcasses reveals the importance of gelatinous material to deep-sea food webs. Proc. R. Soc. B 281, 20142210 (2014).
Bell, M. C., Redant, F. & Tuck I. Nephrops Species. In Lobsters: Biology, Management, Aquaculture and Fisheries (Ed. Phillips, B.F) 412–461 (Blackwell, 2006).
Frandsen, R. P. et al. Sustainable development of the Nephrops fishery in the Kattegat-Skagerrak region. DTU Aqua report no. 298-2015. National Institute of Aquatic Resources, Technical University of Denmark. 23 pp (2015).
Statistics Norway. http://www.ssb.no. Accessed 06/01/2017.
Cristo, M. Feeding ecology of Nephrops norvegicus (Decapoda: Nephropidae). J. Nat. Hist. 32, 1493–1498 (1998).
Parslow-Williams, P., Goodheir, C., Atkinson, R. J. A. & Taylor, A. C. Feeding energetics of the Norway lobster, Nephrops norvegicus in the Firth of Clyde, Scotland. Ophelia 56, 101–120 (2002).
Priede, I. G., Smith, K. L. & Armstrong, J. D. Foraging behavior of abyssal grenadier fish: inferences from acoustic tagging and tracking in the North Pacific Ocean. Deep Sea Res. 37, 81–101 (1990).
Heagney, E. C., Lynch, T. P., Babcock, R. C. & Suthers, I. M. Pelagic fish assemblages assessed using mid-water baited video: standardising fish counts using bait plume size. Mar. Ecol. Prog. Ser. 350, 255–266 (2007).
Tuck, I. D., Chapman, C. J., Atkinson, R. J. A., Bailey, N. & Smith, R. S. M. A comparison of methods for stock assessment of the Norway lobster, Nephrops norvegicus, in the Firth of Clyde. Fish. Res. 32, 89–100 (1997).
Chapman, C. J. & Rice, A. L. Some direct observations on the ecology and behaviour of the Norway lobster Nephrops norvegicus. Mar. Biol. 10, 321–329 (1971).
Atkinson, R. J. A. Spatial distribution of Nephrops burrows. Estuar. Coast. Mar. Sci. 2, 171IN3175–174176 (1974).
ICES. Report of the Working Group on the Assessment of Demersal Stocks in the North Sea and Skagerrak (WGNSSK), Hamburg. ICES, Copenhagen. ICES CM 2016/ACOM (2017, April).
ICES Interim Report of the Working Group on Nephrops Surveys WGNEPS 2016 Report. Reykjavik. ICES, Copenhagen. ICES CM 2016/SSGIEOM (2016, November).
Sardà, F. & Aguzzi, J. A review of burrow counting as an alternative to other typical methods of assessment of Norway lobster populations. Rev. Fish Biol. Fisher. 22, 409–422 (2012).
Aguzzi, J. & Sardà, F. A history of recent advancements on Nephrops norvegicus behavioral and physiological rhythms. Rev. Fish Biol. Fisher. 18, 235–248 (2008).
Doyle, T. K. et al. The energy density of jellyfish: estimates from bomb-calorimetry and proximate-composition. J. Exp. Mar. Biol. Ecol. 343, 239–252 (2007).
Heaslip, S. G., Iverson, S. J., Don Bowen, W. & James, M. C. Jellyfish support high energy intake of leatherback sea turtles (Dermochelys coriacea): video evidence from animal-borne cameras. PLOS ONE 7, e33259 (2012).
Clark, M. R., King, K. J. & McMillan, P. J. The food and feeding relationships of black oreo, Allocyttus niger, smooth oreo, Pseudocyttus maculatus, and eight other fish species from the continental slope of the south‐west Chatham Rise, New Zealand. J. Fish Biol. 35, 465–484 (1989).
Yamamoto, J. et al. Transportation of organic matter to the sea floor by carrion falls of the giant jellyfish Nemopilema nomurai in the Sea of Japan. Mar. Biol. 153, 311–317 (2008).
Haraldsson, M., Tönnesson, K., Tiselius, P., Thingstad, T. F. & Aksnes, D. L. Relationship between fish and jellyfish as a function of eutrophication and water clarity. Mar. Ecol. Prog. Ser. 471, 73–85 (2012).
Pauly, D., Graham, W., Libralato, S., Morissette, L. & Palomares, M. D. Jellyfish in ecosystems, online databases, and ecosystem models. Hydrobiologia 616, 67–85 (2009).
Richardson, A. J., Bakun, A., Hays, G. C. & Gibbons, M. J. The jellyfish joyride: causes, consequences and management responses to a more gelatinous future. Trends Ecol. Evol. 24, 312–322 (2009).
Drazen, J. C., Bailey, D. M., Ruhl, H. A. & Smith, K. L. Jr. The role of carrion supply in the abundance of deep-water fish off California. PLOS ONE 7, e49332 (2012).
Acknowledgements
We would like to thank Captain Leon Pedersen of the RV “Solvik” for skillful assistance at sea, and Nick Higgs and an anonymous reviewer for suggesting improvements to the manuscript. This research was funded by the Norwegian Research Council grant: Combined effects of multiple organic stressors from jellyfish blooms and aquaculture operations on seafloor ecosystems (244572).
Author information
Authors and Affiliations
Contributions
Drs. K. Dunlop, D.O.B. Jones and Dr. A.K. Sweetman were all involved in the field data collection. Dr. K. Dunlop analysed the images and data, and all authors contributed to the writing of the manuscript. All authors reviewed the manuscript prior to submission.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dunlop, K.M., Jones, D.O.B. & Sweetman, A.K. Direct evidence of an efficient energy transfer pathway from jellyfish carcasses to a commercially important deep-water species. Sci Rep 7, 17455 (2017). https://doi.org/10.1038/s41598-017-17557-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-17557-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
