
Abstract
The pervasive and unabated nature of global amphibian declines suggests common demographic responses to a given driver, and quantification of major drivers and responses could inform broad-scale conservation actions. We explored the influence of climate on demographic parameters (i.e., changes in the probabilities of survival and recruitment) using 31 datasets from temperate zone amphibian populations (North America and Europe) with more than a decade of observations each. There was evidence for an influence of climate on population demographic rates, but the direction and magnitude of responses to climate drivers was highly variable among taxa and among populations within taxa. These results reveal that climate drivers interact with variation in life-history traits and population-specific attributes resulting in a diversity of responses. This heterogeneity complicates the identification of conservation ‘rules of thumb’ for these taxa, and supports the notion of local focus as the most effective approach to overcome global-scale conservation challenges.
Similar content being viewed by others

Introduction
Amphibian decline | demography | populations | climate
The response of biodiversity to climate change is often examined at macroecological and temporal scales, treating demographic mechanisms implicitly (e.g.,1, but see2). Demographic rates describe changes in abundance and distribution for populations, but reflect abilities of individual animals to respond to changing conditions and local environments. This implies that environmental variation is a powerful determinant of natural selection and ultimately determines individual fitness, and the dynamics and viability of populations3,4. Thus, a better understanding of the extent to which climate forces changes to population demographic rates, and how this affects the synchronicity of population responses to large-scale environmental fluctuations is critical to understanding the scope of potential losses, predicting future population viability, and developing strategies to preserve biodiversity5,6,7. For example, if a common, and predictable, demographic response (e.g., increasing or decreasing synchrony) to climate change could be identified, global conservation strategies, or “rules of thumb” might also be identified to combat declines.
While large-scale climate conditions can synchronize population dynamics, local environmental conditions often mediate the effects of regional drivers, such that local populations vary asynchronously and are thus more resilient to large-scale climate variation (e.g.,8). Conversely, if demographic rates of populations are synchronized by large-scale environmental forcing, then adverse change in regional conditions may lead to large-scale population declines. Synchronous responses to climate drivers have been identified for populations of different species over large regions1,2,9, suggesting that the potential exists for a single dominant driver to force synchrony in population dynamics by affecting one or more demographic rates. Furthermore, there is evidence that climatic fluctuations contribute to shifts in demographic processes of populations10,11,12,13, and that climate-mediated declines in animal populations5,14 occur. The geographic extent of amphibian decline contributes to the appeal of this phenomenon as a model system to understand the effects of climate on populations and persistence, and widespread declines in amphibian populations15 suggest that a single dominant driver may be forcing regional synchrony in amphibian population dynamics by affecting one or more demographic rates.
Demographic rates that characterize the response of animals to change may respond to variation occurring at two scales16: landscape, where over-arching processes produce similarity in demographic rates (e.g., alignment across populations17,18, and local, where over-arching processes manifest differently and produce differences in demographic rates (e.g., increased local variation). Directional changes in climate, such as long-term shifts in mean precipitation and timing, can act at a landscape scale and influence populations across a large area19, whereas outcomes of such shifts (e.g., amount of snowpack or hydroperiod length) can act at local scales and affect only individual populations because the shifts are mediated by other factors that vary locally (e.g., soil type). For example, a broad scale effect is illustrated by the link between climate change and the collapse of population cycles20, and a local effect is illustrated by the influence of snow depth as a consequence of the weather on the synchrony in population growth rates21.
While climate can influence vertebrate populations at large scales22, most taxa evaluated have either large ranges or are migratory. Amphibians have relatively small ranges and may not track climate changes as well as more mobile taxa23, and thus be more sensitive. Climate change is expected to affect both mean and variance of environmental temperature and water availability24,25, and this shift is accelerating26. As ectotherms with permeable skin, amphibians are sensitive to environmental temperature and moisture27, and we expect that demographic rates in amphibians are especially sensitive to a changing climate, especially to increases in the frequency or intensity of extreme conditions28,29,30. Typically, local conditions, unrelated to weather, buffer against climatic fluctuations. However, the current magnitude and rate of climate change suggest that forcing of regional water availability (i.e., drought) and temperature has the potential to over-ride local variation in these characteristics, leading to an alignment in among-population demographic rates. For example, many temperate amphibian species exist as metapopulations31, but demography in populations fluctuates in response to local conditions11,32, suggesting that large metapopulations are maintained when regional synchrony (due to broad-scale environmental variation) is low.
Current knowledge about the response of amphibian populations to climate is based largely on analyses of single-population datasets. Furthermore, when large-scale drivers are filtered through local site and population characteristics8, it is difficult to identify common factors driving demographic trends. The identification of common population responses to changes in climate can be improved by articulating and testing hypotheses about responses of demographic rates to environmental covariates using long-term data from multiple species within single taxa. This information will improve understanding of the relative contribution of regional and local drivers which are critical for estimating adaptive capacity and choosing conservation actions.
Based on evidence of the substantial magnitude and increasing rate of climate change24,25,26, and the knowledge that weather affects demographic rates28,33,34,35, we quantified survival and recruitment in relation to locally derived climate variables to gain a mechanistic understanding of amphibian demographic responses across a range of species and locations. We used long-term (≥10 or more consecutive years), individual-based, information from 31 populations of amphibians from four countries in Europe and North America representing three climatic “zones”: montane, Mediterranean, and maritime (Canada [n = 1], France [n = 5], Germany [n = 2], and USA [n = 23]) (Fig. 1 and S1) with which to estimate the effect of climate forcing on demography. Capture-mark-recapture data are uniquely rich and provide the most detailed information on free-ranging populations. Our depth of data allows consideration of hypotheses that relate demographic parameters to broad-scale climate, as represented by specific local climate variables. We selected covariates representing local environmental conditions that are driven by broad-scale climatic changes in temperature and water availability. We predicted demographic responses in a suite of a priori hypotheses (Table 1) iteratively developed by expert amphibian researchers with specific knowledge of local populations and habitats – collectively representing >300 years of field expertise. We expected that the demography of species with similar life histories (i.e., terrestrial vs aquatic hibernation), and those inhabiting similar environments (i.e., maritime, Mediterranean, and montane zones) would have similar associations with climate drivers. We also expected that survival, recruitment (i.e., the per capita number of individuals recruited into the breeding populations per year) and population growth rate (i.e., survival + recruitment) might each be influenced in a common manner across populations of amphibian species when they experience similar changes in abiotic conditions. While the magnitude of effect is not expected to be identical among all species and populations, we expected that the direction of the effect would be consistent. Although commonalities among demographic rates or characteristics can exist without synchrony in environmental conditions, synchronous relationships may be an early indicator of population collapse as the buffering that diversity provides is weakened.

Location of study sites. (A) North America, (B) Europe. Numbers correspond to data sets (S1). Black = bufonids; light grey = treefrogs; dark grey = ranids; white = salamanders and newts. Size of dot represents length of data set: large >20 yrs (n = 6); medium = 15–20 yrs (n = 5); small = 10–14 yrs (n = 20). Map produced using ArcMap Version 10.3.
Results
We analyzed data from 11 species and 31 populations where individuals were captured annually from 10 to 28 years (average = 15.3 yrs). We found evidence for an influence of climate on demography (Akaike weight >0.40, 18/31 datasets), although the magnitude and direction of responses to climate drivers was highly variable. Capture probabilities ranged from <0.01–0.74, mean 0.16 (S2). Survival and recruitment estimates were within published norms for all species, taxa were not evenly distributed among climate zones (5 species in maritime, 2 in Mediterranean, and 5 in montane) and population sizes varied. For example, in the Mediterranean zone, 8 of 9 datasets were from small populations of ranid frogs in California.
Adult survival
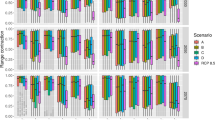
No single hypothesis was strongly supported across all datasets; models including each of our hypothesized drivers were the best supported in at least one case (Fig. 2, S3 and S4). While most of the sites in the Mediterranean zone (8/9) followed predictions for the hypothesized direction of the effect, only about half of the maritime (4/7) and montane (7/15) sites followed our predictions. For most sites (23/31), top models for survival included winter-related covariates (H3, H4, H5). QAICc weights and r2 values (S3, S5) indicated that models that included the length of winter (H3) and winter severity (H4) were often best supported. Covariates in top models more often followed the predicted direction for frogs (13/17 populations) and salamanders and newts (3/5 populations) than for toads (3/9 populations). The direction of the effect of the covariate in the top model was more often as predicted for aquatic hibernators (11/15) than for terrestrial hibernators (8/16).

Hypotheses supported by models (based on the covariate [log-odds] coefficient parameter) are indicated by an arrow. The predicted effect for hypotheses H1 – H8 was negative, but the predicted effect for H9 was positive. Direction of arrow indicates direction of support (H1-H8: down = as predicted, up = contrary to prediction; H9: up = as predicted, and down = contrary to prediction). Data sets are ordered from low to high elevation. See S1 for details on datasets and species names and Table 1 for hypotheses. Zone: MAR – maritime, MED – Mediterranean, MON – montane.
Recruitment
Similarly, no single recruitment hypothesis received overwhelming support across datasets (see Methods for definition). Each of the hypothesized drivers were in the top model for at least one amphibian population (Figs 2, S4 and S5). Top models for recruitment also included winter-related covariates (H7, H8, H9) for 24 of 31 sites. Model results were consistent with predictions 39% of the time (13/31 datasets). The direction of even well-supported effects sometimes differed among sites. For example, cold temperature (H8) was in the top model for 11/31 data sets but the direction of the effect at 7/11 sites was contrary to our prediction of decreased recruitment with increased freezing events. Other covariates appeared in top models but results were split evenly between the direction of the effect supporting or countering our predictions. Parameter estimates (model-averaged across all sites) indicated the most support for models that included drought (H6, β = 0.001, SE = 3.60E-05) and early winter (H9, β = 0.001, SE = 4.60E-05) (S3 and S4). The covariates in the top models for recruitment were poor predictors of the hypothesized effect (i.e., they had weak estimated effect and high uncertainty) in all zones, although half of the top models in the montane zone followed our predictions. A posteriori examination of the results indicated that the correspondence of estimated and predicted direction of effect was not related to taxon.
Variation
The influence of climate on demography was best illustrated in the variance analyses. The most supported models for survival explained over 25% of the temporal deviance (aka variation) in 19/31 datasets (12/19 where model results were as predicted). The best supported models for recruitment explained over 25% of the variation in 16/31 datasets (6/16 where model results were as predicted) (S5).
By zone
In top models for survival, where the effect was in the predicted direction, hypotheses explained 11–44% of the variation in survival for amphibians in the maritime zone; 10–69% of the variation in survival in the Mediterranean zone; and 10–55% of the variation in survival in the Montane zone (S5). In top recruitment models where the effect was in the predicted direction, the amount of variation explained was 19–41% maritime; 50–53% Mediterranean; and 6–46% montane (S5).
By taxon
In top models for survival, where the effect was in the predicted direction, hypotheses explained 10–69% of the variation for survival in frogs (ranids and treefrogs), 11–46% of the variation for survival in toads, and 10–44% of the variation in newts (S6B). In top recruitment models where the effect was in the predicted direction, the amount of variation explained was 6–53% for frogs (ranids and treefrogs), 8–46% for toads, and 19–41% for newts (S5).
By hibernation mode
In top models for survival, where the effect was in the predicted direction, hypotheses explained 10–48% of the variation for survival in amphibians that hibernate terrestrially and 19–69% of the variation for survival in amphibians that hibernate aquatically (S5). In top recruitment models where the effect was in the predicted direction, the amount of variation explained was 8–46% for terrestrial hibernators and 6–53% for aquatic hibernators (S5).
Discussion
We provide empirical evidence for the idea that climate is one of the overarching drivers for change in amphibian demographics and that demographic parameters are influenced by climate-related covariates for amphibian populations in multiple temperate climates. Although there are numerous single species (or single locale) studies that examine how amphibians respond to their environment36,37, and link climate to changes in demography14,29,34,35,38, we capitalized on the greater taxonomic, spatial and temporal scales of our data, and hypotheses formulated on theory, empirical evidence, and expert elicitation (i.e., our collective field experience), to assess the potential for common climate-driven responses of multiple species across large geographic regions.
Our analyses of 31 long-term demographic datasets revealed variable demographic responses to climate variables both within and across species, and within and across climatic zones. In 18 of 31 datasets, the top candidate models were strongly supported with AICc wts of >0.40; in nine of those 18, AICc wts were >0.90. We considered this to be strong support because 0.4 is 10 times greater than the AICc wt one would expect if all models were equally supported (i.e., for each dataset we developed 30 models for survival and 25 for recruitment. If all models had equal support, we would expect an AICc wt of 0.04 (1/25) or 0.03 (1/30) respectively. Thus, our analysis provides empirical support for the effect of climate variables on demographic processes across multiple amphibian taxa and across multiple localities. The influence of the covariates was also supported by the amount of variation explained by the hypotheses. In a majority of datasets, ≥25% of the variation was explained for both survival and recruitment, further highlighting the importance of climate.
While no single covariate emerged as a main driver of demographic variation, in the majority of datasets, winter-related covariates were in the top models for both survival and recruitment, and 65% and 46% of these top models indicated that the effect was negative (decreased survival and decreased recruitment respectively). Although we might expect winter covariates to occur in top models 60% of the time by chance alone, a range of previous work highlights the importance of winter covariates. For example a synthesis of organismal responses to winter climate change reports that community composition, ecological interactions, and individual performance are affected by winter39. Other species-specific studies, including those focused on amphibians, indicate sensitivity to winter covariates29,40,41, and support our finding that populations are more sensitive to variation in winter conditions than conditions during the breeding and active seasons.
We hypothesized that we would see similar demographic responses to climate-related covariates across climatic zones and species, and that such uniformity might suggest that local responses to specific local conditions were being over-ridden. Contrary to our hypothesis, we identified a variety of responses both within and among amphibian species, suggesting that response to climate is highly context-dependent and that conservation planning will require a local approach despite the expected magnitude of climate change and potential for climate forcing. For example, there is a need to better determine how context-dependency can mediate the effect of climate on local populations and promote their resilience. Local contexts may include geographic, biotic and abiotic factors that differ in importance (e.g., climate forcing42; range position43; phenotypes44; geography45; habitat characteristics35). We expand on three examples from our analysis, but note that there are a number of interesting patterns that could be assessed further.
Three boreal toad populations in the Rocky Mountains occur in similar habitats, yet there was little uniformity in the survival model best supported by the data, and demographic response to covariates was often contrary to predictions. Models including the recurrence of warm events during winter (H5) had the most support, but affected survival as predicted (negatively) in only one population (at the highest elevation). High snowpack provides insulation during winter and lack of insulation can precipitate inappropriate rousing and waste of energy40. The criticality of this insulation, and thus the response in survival rate, likely depends on the elevation and physiogeography of local sites.
Results from great crested newts in France supported the importance of winter-related covariates (H5 and H4), but might also represent differences in demography related to the position of populations within the species’ range. Our newt populations are at the very southern edge of their range in contrast to newt studies in Britain34. The effect on survival of the number of warm days during hibernation (H5) had the most support in one population, but the direction of the effect, increased survival, was contrary to our prediction. Interestingly, results reported in Griffiths et al.34 – decreased survival with mild winters and heavy rain – support our H5. In our second newt dataset, survival decreased as winter severity increased (H4), as we predicted. Griffiths et al.34 provide evidence for the effect of climate factors at a regional level, but also asynchronies in subpopulation dynamics. Disparate results among our newt datasets echo these results.
There were also discrepancies compared to results published previously for Columbia spotted frogs. The best-supported model for one of two Columbia spotted frog populations in Montana suggested that survival increased as winter length (severity) increased, contrary to what we predicted, and contrary to an earlier analysis46. We attribute this to differences in characterization of winter covariates and possibly the inclusion of 5 additional years of data. This also highlights the challenge of using real field data to search for “cause” using correlational analyses.
Our analysis convincingly illustrates a pattern of local responses to similar climate-related stressors and fails to support an overarching effect of climate change on amphibian demographics. The variability we observed in demographic response, within and among species and populations, emphasizes the potential for species to adapt to local conditions and persist in changing environments, and is concomitant to the variability found among regions (i.e., different stressors in different regions, sensu47). These data provide population-level evidence to support a local, rather than a centralized, approach to conservation and illustrate the critical conservation need to maintain variability at every taxonomic level.
Methods
Species and sites
We used capture-mark-recapture data from pond-breeding amphibians (5 frogs, 3 toads and 2 newts), and one stream-breeding salamander (Salamandra salamandra) collected between 1965 and 2014. Eleven sites had 15–28 yr of data and 20 sites had 10–14 yr of data. Study sites were located in temperate regions; Mediterranean, montane or maritime climatic zones in Europe and North America (S1), and all animals were native to the locations where they were captured.
Field methods
Animals were captured by hand or net; individual marks were applied to animals captured for the first time and existing marks (signifying recaptured animals) were recorded. Marking techniques varied by study and included passive integrated transponder tags, toe-clipping, visual implant elastomer, and identification based on photographs48. Data collection followed standard capture-mark-recapture study designs49 and all methods were carried out in accordance with relevant guidelines and regulations. In most studies, sites were visited at least three times during the breeding season each year and data conformed to Pollock’s robust design49. For datasets that included information for both sexes, we controlled for sex differences in survival and recruitment when modelling covariate effects.
Analyses
We used the temporal-symmetry approach50 to estimate annual apparent survival, recruitment, and capture probabilities, and tested hypotheses about the influence of covariates on these parameters using a model selection approach. We assessed goodness-of-fit and accounted for potential over-dispersion using c-hat49. Apparent survival is defined as the probability of an adult individual surviving and not permanently emigrating from the study site between sampling occasions t and t + 1 and recruitment is defined as the ratio of new adults entering the focal population between year t and year t + 1 to the number of adults that were already present in the population (and exposed to capture) in sampling season t50. Recruitment encompasses two sources of new adults entering the focal population: 1) juveniles from the local population becoming adults and 2) immigration of adults from nearby populations. We assume that the majority of recruitment originated from local juvenile-to-adult transitions such that our measures of recruitment are representative of the local breeding process, although we acknowledge that this assumption may not hold for species with high dispersal rates51. Capture probability is the probability that a marked individual is available for detection (i.e., present at the site) and is detected during a sampling period. We used the robust design version of the Pradel model for all datasets except 7 and 8, where there was only a single visit per season, for those datasets we used the non-robust design version of the Pradel model50.
We developed a set of 9 a priori hypotheses (Table 1) to investigate the potential influence of environmental covariates with input from all the data providers. Hypotheses related environmental factors (water availability and temperature) to probabilities of survival (ϕ) and recruitment (f). Hypotheses were based on the reliance of our study species on standing water bodies and on our interpretation of the likely response (physiological or physical) of amphibians to extremes in temperatures. We developed five hypotheses for survival and four for recruitment.
In some cases, hypotheses were irrelevant for particular species and not assessed and in some of the recruitment hypotheses covariates were necessarily lagged (S1). For example, Hypothesis 4 (Table 1) is irrelevant for species inhabiting localities where snowpack does not occur, or for those species that hibernate aquatically. The covariates can reflect local variation as well as broad-scale change but hypotheses were developed considering local effects.
For each hypothesis a focal covariate was developed using daily precipitation, temperature, and snowpack data derived from local climate data (S6) specific for each study site. This strategy ensured that while we addressed consistent ecological hypotheses that might influence population dynamics, we allowed for the specific form of the influence to reflect local conditions. Because our interest was the mechanisms relating population fluctuations to relevant climate variables, this distinction, and focus on local conditions, is more appropriate than selecting a single consistent (large-scale) variable, and ensures that any deviation from expectation does not result from an inappropriate large-scale variable.
Models were implemented in Program MARK52 accessed through R (v. 3.1.0; R Development Core Team 2011), using the RMark interface53. Relative support for competing hypotheses was assessed via model selection54 performed in several steps, 1) select the best covariate structure for capture probability, keeping a full-time varying structure for survival and recruitment, 2) select the best covariate structure, regarding sex effects for survival and recruitment probabilities, and 3) assess the relative support for each hypothesis for each dataset. Models were selected using AIC corrected for small sample size (AICc) and overdispersion (QAICc) by a variance inflation factor (ĉ). For each dataset, ĉ was calculated (program RELEASE global test: TEST 2 + TEST 3; 51). A value of ĉ = 1 was assumed when the calculated ĉ was<1. QAICc weights were also used to assess the degree of support for each model. First, we assessed three hypotheses for capture probability: 1) p constant [p(.)]; 2) p as a linear function of capture occasion within each year; and 3) p as a quadratic function of capture occasion within each year. We tested for differences in detection probability between sexes when data were available. We selected the model with the most support for p and used that parameterization in all subsequent models. Next, we assessed the effect of sex on survival and recruitment for datasets with that information. If an effect of sex was supported, it was included in subsequent models. We then assessed the support for each hypotheses to quantifying the influence of each covariate on demographic parameters. Survival and recruitment are estimated simultaneously in Pradel models, thus, for each dataset, we assessed all possible combinations of 6 hypotheses for survival (H0-H5) and 5 hypotheses for recruitment (H0, H6-H9): 30 competing models (6 × 5 = 30 combinations). Hypothesis 4 was not relevant for 15 datasets (25 competing models (5 × 5 combinations). Parameter estimates from top models, and cumulative QAICc weights were used to quantify the degree of support for each hypothesis. We then compared the magnitude and the direction of influence of each covariate relative to a priori predictions.
We performed an analysis of deviance to quantify the amount of variation in survival and recruitment explained by the covariates16 (S6). This test relies on an F-statistic based on the deviances, and the corresponding degrees of freedom, of models assuming full-time variation (Ft), constancy (F0) and covariate-driven variation (FH, our nine covariates) for the parameter of interest (e.g., survival), and assuming the best supported parameterization for the other model parameters (i.e., capture probability and recruitment). Using ratios of deviances from these models, we calculated the proportion of variation (r²) explained by each covariate (r² = Dev(F0) − Dev(FH)/Dev(F0) − Dev(Ft)). We assessed the amount of variation explained by grouping data sets by taxonomy; life history (i.e., hibernation site – aquatic vs terrestrial); and by environment (maritime, Mediterranean, or montane) (S6A, B).
We ran full-time varying models for survival (Φt) and recruitment (ft) to provide annual estimates of these parameters, for each site. We plotted annual estimates and predicted annual values from the best supported covariate model (S3). We subsequently derived inter-annual values of population growth λt, as λt = Φt + ft for each and plotted annual estimates to illustrate the trajectory for each population (S7).
Data Availability
Derived data are available in the online appendices and at the John Wesley Powell Center for Synthesis and Analysis (https://powellcenter.usgs.gov/data-resources). Raw data are available from the authors of each respective dataset.
References
Koenig, W. D. & Liebhold, A. M. Temporally increasing spatial synchrony of North American temperature and bird populations. Nat. Clim. Change 6, 614–617 (2016).
Pomara, L. Y. & Zuckerberg, B. Climate variability drives population cycling and synchrony. Divers. Distrib. 23, 421–434 (2017).
Darwin, C. On the origins of species by means of natural selection 247 (Murray, London, 1859).
Den Boer, P. J. Spreading of risk and stabilization of animal numbers. ACTA Biotheor. 18, 165–194 (1968).
Selwood, K. E., McGeoch, M. A. & MacNally, R. The effects of climate change and land‐use change on demographic rates and population viability. Biol. Rev. 90, 837–853 (2014).
Schurr, F. M. et al. How to understand species’ niches and range dynamics: a demographic research agenda for biogeography. J. Biogeogr. 39, 2146–2162 (2012).
Amburgey, S. M. et al. Range position and climate sensitivity: The structure of among‐population demographic responses to climatic variation. Glob. Change Biol. 00, 1–16, https://doi.org/10.1111/gcb.13817 (2017).
Schindler, D. E., Armstrong, J. B. & Reed, T. E. The portfolio concept in ecology and evolution. Front. Ecol. Environ. 13, 257–263 (2015).
Moran, P. A. P. The statistical analysis of the Canadian lynx cycle. II. Synchronization and meteorology. Aust. J. Zool. 1, 291–298 (1953).
Gaillard, J. M. et al. How does climate change influence demographic processes of widespread species? Lessons from the comparative analysis of contrasted populations of roe deer. Ecol. Lett. 16, 48–57 (2013).
Cayuela, H. et al. The impact of severe drought on survival, fecundity, and population persistence in an endangered amphibian. Ecosphere 7, e01246 (2016).
Green, T., Das, E. & Green, D. M. Springtime emergence of overwintering toads, Anaxyrus fowleri, in relation to environmental factors. Copeia 104, 393–401 (2016).
Pacifici, M. Species’traits influenced their response to recent climate change. Nat. Clim. Change On-line 13 February 2017, 101038/NCLIMATE3223 (2017).
Lowe, W. H. Climate change is linked to long-term decline in a stream salamander. Biol. Conserv. 145, 48–53 (2012).
Houlahan, J. E., Findlay, C. S., Schmidt, B. R., Meyer, A. H. & Kuzmin, S. L. Quantitative evidence for global amphibian population declines. Nature 404, 752–755 (2000).
Grosbois, V. et al. Assessing the impact of climate variation on survival in vertebrate populations. Biol. Rev. Camb. Philos. 83, 357–99 (2008).
Ims, R. A. & Andreassen, H. P. Spatial demographic synchrony in fragmented populations. Landscape ecology of small mammals. (Springer New York, 1999).
Schaub, M., Hirschheydt, J. & Grüebler., M. U. Differential contribution of demographic rate synchrony to population synchrony in barn swallows. J. Anim. Ecol. 84.6, 1530–1541 (2015).
Post, E. & Forchhammer, M. C. Synchronization of animal population dynamics by large-scale climate. Nature 420(6912), 168–171 (2002).
Ims, R. A., Henden, J. A. & Killengreen, S. T. Collapsing population cycles. Trends Ecol. Evol. 23, 79–86 (2008).
Grøtan, V. et al. Climate causes large-scale spatial synchrony in population fluctuations of a temperate herbivore. Ecology 86, 1472–1482 (2005).
Liebhold, A., Koenig, W. D. & Bjørnstad, O. N. Spatial synchrony in population dynamics. Annu. Rev. Ecol. Evol. Syst. 35, 467–490 (2004).
Lawler, J. J. et al. Projected climate-induced faunal change in the Western Hemisphere. Ecology 90, 588–97 (2009).
Barnett, T. P., Adam, J. C. & Lettenmaier, D. P. Potential impacts of a warming climate on water availability in snow-dominated regions. Nature 438, 303–309 (2005).
Taylor, R. G. et al. Ground water andclimate change. Nat. Clim. Change 3, 322–329 (2013).
Smith, S. J., Edmonds, J., Hartin, C. A., Mundra, A. & Calvin, K. Near-term acceleration in the rate of temperature change. Nat. Clim. Change 5, 333–336 (2015).
Carey, C. & Alexander, M. A. Climate change and amphibian declines: is there a link? Divers. Distrib. 9, 111–121 (2003).
Reading, C. J. Linking global warming to amphibian declines through its effects on female body condition and survivorship. Oecologia 151, 125–131 (2007).
McCaffery, R. M., Solonen, A. & Crone, E. Frog population viability under present and future climate conditions: a Bayesian state-space approach. J. Anim. Ecol. 81, 978–985 (2012).
Ficetola, G. F. Habitat conservation research for amphibians: methodological improvements and thematic shifts. Biodivers. Conserv. 24, 1293–1310 (2015).
Smith, A. M. & Green, D. M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: are all amphibian populations metapopulations? Ecography 28(1), 110–128 (2005).
Green, D. M. The ecology of extinction: population fluctuation and decline in amphibians. Biol. Conserv. 111, 331–343 (2003).
Scherer, R. D., Muths, E., Noon, B. R. & Corn, P. S. An evaluation of weather and disease as causes of decline in two populations of boreal toads. Ecol. Appl. 15, 2150–2160 (2005).
Griffiths, R. A., Sewell, D. & McCrea, R. S. Dynamics of a declining amphibian metapopulation: survival, dispersal and the impact of climate. Biol. Conserv. 143, 485–491 (2010).
Cayuela, H. et al. Demographic responses to weather fluctuations are context dependent in a long‐lived amphibian. Global Change Biol 22, 2676–2687 (2016).
Orizaola, G., Quintela, M. & Laurila, A. Climatic adaptation in an isolated and genetically impoverished amphibian population. Ecography 33, 730–737 (2010).
Hossack, B. R. et al. Roles of patch characteristics, drought frequency, and restoration in long-term trends of a widespread amphibian. Conserv. Biol. 27, 1410–1420 (2013).
McCaffery, R. M., Eby, L. A., Maxell, B. A. & Corn, P. S. Breeding site heterogeneity reduces variability in frog recruitment and population dynamics. Biol. Conserv. 170, 169–176 (2014).
Williams, C. M., Henry, H. A. & Sinclair, B. J. Cold truths: how winter drives responses of terrestrial organisms to climate change. Biol. Rev. 90, 214–235 (2014).
Sinclair, B. J. et al. Real-time measurement of metabolic rate during freezing and thawing of the wood frog, Rana sylvatica: implications for overwinter energy use. J. Exp. Biol. 216, 292–302 (2013).
Roland, J. & Matter, S. F. Pivotal effect of early‐winter temperatures and snowfall on population growth of alpine Parnassius smintheus butterflies. Ecol. Monogr. 86, 412–428 (2016).
Stenseth, N. C. et al. Ecological effects of climate fluctuations. Science 297, 1292–1296 (2002).
Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669 (2006).
Ozgul, A. et al. Coupled dynamics of body mass and population growth in response to environmental change. Nature 466, 482–485 (2010).
Dobrowski, S. Z. A climatic basis for microrefugia: the influence of terrain on climate. Global change biol. 17, 1022–1035 (2011).
McCaffery, R. M. & Maxell, B. A. Decreased winter severity increases viability of a montane frog population. P. Natl. Acad. Sci. USA 107, 8644–8649 (2010).
Grant, E. H. C. et al. Quantitative evidence for the effects of multiple drivers on continental-scale amphibian declines. Scientific reports 6, 25625 (2016).
Dodd, C. K. (ed). Amphibian Ecology and Conservation 566 (Oxford University Press, 2010).
Williams, B. K., Nichols, J. D. & Conroy, M. J. Analysis and management of animal populations (Academic Press, 2002).
Pradel, R. Utilization of capture-mark-recapture for the study of recruitment and population growth rate. Biometrics 52, 703–709 (1996).
Perret, N., Pradel, R., Miaud, C., Grolet, O. & Joly, P. Transience, dispersal and survival rates in newt patchy populations. J. Anim. Ecol. 72, 567–575 (2003).
White, G. C. & Burnham, K. P. Program MARK: survival estimation from populations of marked animals. Bird study 46(sup1), S120–S139 (1999).
RMark: an R interface for analysis of capture-recapture data with MARK U. S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Alaska Fisheries Science Center (2013).
Burnham, K. P. & Anderson, D. R. Model Selection and Multimodel Inference, a Practical Information–theoretic Approach 2nd edn 98–143 (Springer, 2002).
Acknowledgements
We thank the legions of field technicians that participated in the collection of these data, as well as the many government and non-government organizations that made data collection possible. This analysis was initiated at the Amphibian Decline Working Group supported by the John Wesley Powell Center for Analysis and Synthesis, funded by the US Geological Survey. This manuscript is contribution #605 of the USGS Amphibian Research and Monitoring Initiative. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Contributions
E.M., T.C., D.A.W.M., E.H.C.G. conceived the study with contributions from the Amphibian Decline Working Group, T.C. analyzed the data, E.M., T.C., E.H.C.G., B.R.S., D.A.W.M., and B.R.H. wrote the manuscript with contributions from P.J., O.G., D.M.G., D.S.P., M.C., R.N.F., R.M.M., M.J.A., A.B., W.P., J.W.A., J.G., G.F., and J.M.T. E.M., B.R.S., B.R.H., P.J., O.G., D.M.G., D.S.P., M.C., R.N.F., R.M.M., M.J.A., W.P., J.W.A., J.G., G.F., and J.M.T. provided capture-recapture data. After the first 5 authors, authors are listed based on the length of the contributed data set, if data sets were the same length, authors are listed alphabetically (except for the final author).
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Muths, E., Chambert, T., Schmidt, B.R. et al. Heterogeneous responses of temperate-zone amphibian populations to climate change complicates conservation planning. Sci Rep 7, 17102 (2017). https://doi.org/10.1038/s41598-017-17105-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-17105-7
This article is cited by
-
Modelling Climate-Change Impact on the Spatial Distribution of Garra Rufa (Heckel, 1843) (Teleostei: Cyprinidae)
Iranian Journal of Science and Technology, Transactions A: Science (2021)
-
Amphibian and reptile phenology: the end of the warming hiatus and the influence of the NAO in the North Mediterranean
International Journal of Biometeorology (2020)
-
Mapping the current and future distributions of Onosma species endemic to Iran
Journal of Arid Land (2020)
-
Modelling and mapping the current and future potential habitats of the Algero-Tunisian endemic newt Pleurodeles nebulosus under climate change
European Journal of Wildlife Research (2020)
-
A demographic approach to understanding the effects of climate on population growth
Oecologia (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
