
Abstract
Multiple infections are common in honey bees, Apis mellifera, but the possible role of nutrition in this regard is poorly understood. Microsporidian infections, which are promoted by protein-fed, can negatively correlate with virus infections, but the role of protein nutrition for the microsporidian-virus interface is unknown. Here, we challenged naturally deformed wing virus - B (DWV-B) infected adult honey bee workers fed with or without pollen ( = protein) in hoarding cages, with the microsporidian Nosema ceranae. Bee mortality was recorded for 14 days and N. ceranae spore loads and DWV-B titers were quantified. Amongst the groups inoculated with N. ceranae, more spores were counted in protein-fed bees. However, N. ceranae infected bees without protein-diet had reduced longevity compared to all other groups. N. ceranae infection had no effect on protein-fed bee’s longevity, whereas bees supplied only with sugar-water showed reduced survival. Our data also support that protein-feeding can have a significant negative impact on virus infections in insects. The negative correlation between N. ceranae spore loads and DWV-B titers was stronger expressed in protein-fed hosts. Proteins not only enhance survival of infected hosts, but also significantly shape the microsporidian-virus interface, probably due to increased spore production and enhanced host immunity.
Similar content being viewed by others


Introduction
Host nutrition can play a key role for the outcome of pathogen infections in humans and animals1, since it is critical for immune-defense and resistance to pathogens2. Poor nutrition, in particular protein depletion, is a major factor in high incidence and mortality due to infectious diseases2,3.
In insects, the role of nutrition for the outcome of infections is less well understood4. The importance of proteins for pathogen resistance has been suggested in caterpillars, Spodoptera littoralis, where their resistance to viral infection increased as the protein to carbohydrate ratio in their diet increased5. Moreover, infected larvae of the African moth, Spodoptera exempta, select a higher protein diet, suggesting that nutrition has a self-medication value6.
Feeding protein to honey bee, Apis mellifera, workers infected with microsporidian endoparasites, Nosema apis, resulted in increased spore development, but also improved the longevity of infected hosts7. Similar findings were reported for Nosema ceranae 8. In bumblebees, Bombus terrestris, protein deprivation nutrition can functionally alter not only general resistance, but also alter the pattern of specific host–parasite interactions, probably due to reduced immune responses9.
Since hosts infected by more than one pathogen are common, pathogen-pathogen interactions require more attention10,11,12,13. This is especially true for managed honey bees, A. mellifera, which are exposed to a long list of pathogens, which can act as drivers for colony losses especially in areas with established ectoparasitic mite, Varroa destructor, populations14,15. Since many honey bee pathogens are ubiquitous16 multiple viral, fungal and bacterial infections of colonies and even individual bees are most likely and can result in lethal effects to the host14.
However, the actual outcome of such multiple infections depends on the nature of interactions between the pathogens in one host. These parasite-parasite interactions in infected individual honey bee hosts can potentially range from competition to cooperation17,18,19,20,21,22.
For example, V. destructor is intimately associated with viruses, e.g. deformed wing virus (DWV)23,24,25, especially because it is a very efficient virus vector, generating a disease epidemic within the colony, which dwindles until it dies26. V. destructor can also activate latent virus infections27. On the other hand, there is evidence for antagonistic interactions between honey bee parasites, e.g. between the microsporidians Nosema ceranae and Nosema apis 28, N. ceranae and DWV29,30. Synergistic effects have been reported between N. apis and several viruses, e.g. filamentous virus, bee virus Y and black queen cell virus (BQCV)31,32. In contrast, no association was found between Israel acute paralysis virus (IAPV) and Nosema ceranae 33. This large range of possible interactions between pathogens in one host calls for investigation of possible mechanisms driving this interface.
Since it is known that proteins can impact both virus and microsporidian infections in insects, we regard it as likely that this will affect the outcome of virus-microsporidian interactions in multiple infected hosts. Pollen is the main natural source of protein for honey bees, especially for young bees. It is an essential protein source and may interfere with pathogen-pathogen interactions, infection intensity and longevity of the honey bee host34,35.
Here, we investigated the possible role of protein feed via pollen on the interface between DWV-B (formerly Varroa destructor virus-1) and Nosema ceranae in individual honey bee workers. We hypothesize that protein fed in the form of pollen will not only have significant beneficial effects for the hosts, but will also amplify the virus-microsporidian interface.
Results
Since the experimental pollen was not irradiated, some bees (N = 19) of the Pollen-only treatment were naturally infected with N. ceranae. These contaminated bees were excluded from further data analyses.
The virus strain-specific PCR36 showed that only DWV-B was infesting the experimental bees. Neither BQCV, DWV, nor acute bee paralysis virus (ABPV) were found in any of the analyzed individual honey bee workers (N = 120).
All data (N. ceranae spore loads, DWV-B infection levels, sugar and pollen consumption) were not normally distributed (Shapiro-Wilk’s test for normality, P < 0.05 in all cases). Therefore, the non-parametric Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Tests were performed.
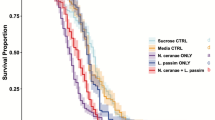
The survival in the different groups is shown in (Fig. 1). While a significantly reduced longevity was observed for Nosema-only infected bees compared to all other groups (Kaplan-Meier, Log-Rank test, P < 0.05), N. ceranae infection had no significant effect on the longevity of pollen-fed bees (Kaplan-Meier, Log-Rank test, P > 0.05). Bees that were supplied only with sugar water showed a reduced survival compared to the bees which received both sugar and pollen (Kaplan-Meier, Log-Rank test, P < 0.05, Fig. 1). Hence, workers exposed to both N. ceranae and pollen showed a non-additive effect when compared to both treatments individually (χ2 = 22.73 (equation-3), theoretical χ2 = 7.879, df = 1, P = 0.005). Due to the calculated negative value −33.87 (equation-4), the observed effect on mortality can be considered antagonistic.

Cumulative survival of honey bee workers exposed to the different treatments over time. Workers contaminated with N. ceranae from the pollen-only treatment are not considered. Significant differences (Log-Rank test P < 0.05) among treatments are indicated by different letters (a, b, c).
Among the four treatments (Control; Pollen-only; Nosema-only; Nosema-pollen) significant differences in N. ceranae spore loads were only found between Nosema-only and Nosema-pollen groups (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 2.64, P = 0.025, Fig. 2). Non-inoculated bees from the control group showed no N. ceranae infections.

N. ceranae spore loads of individual honey bee workers in the four treatment groups (Controls, Pollen-only [orange], Nosema-only [green], Nosema-pollen [blue]). Medians, ranges, confidence intervals and outliers ( = dots) are shown at a log scale. Significant differences were found between the groups Nosema-pollen and Nosema-only, as well as between the two Nosema groups and the Controls. Please note that N = 19 bees in the Pollen-only treatment were naturally contaminated with N. ceranae. When excluding these contaminated bees from the Pollen-only group, significant differences were still found between the groups Nosema-pollen and Nosema-only (*P = 0.025, **P < 0.0001).
The naturally occurring DWV-B infections were significantly different between the four treatment groups (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 2.64, P = 0.006, Fig. 3). A significant higher virus load was observed in the Control group compared to the Pollen-only treatment (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test * = z > 1.96, P < 0.0001). There were no significant differences between the other groups: (i) for Control and Nosema-only (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.14), (ii) Controls and Nosema-pollen (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.1), (iii) Pollen-only and Nosema-only (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.22), (iv) Pollen-only and Nosema-pollen (Kruskal- Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.06), (v) Nosema-only and Nosema-pollen (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.75).

DWV-B infection loads of individual honey bee workers in the four treatment groups (Controls [white], Pollen-only [orange] Nosema-only [green], Nosema-pollen [blue]). Medians, ranges, confidence intervals and outliers ( = dots) are shown at a log scale. Workers contaminated with N. ceranae from the pollen-only treatment are not considered. Significant differences were found between the controls and the Pollen-only group (*P < 0.0001).
The correlation between N. ceranae spore loads and DWV-B infection levels was not significant in the Nosema-only treatment (Pearson Correlation: Pearson |r| = −0.22, P = 0.12; Fig. 4). However, a significant negative correlation was found between N. ceranae spore loads and DWV-B infection levels in the Nosema-pollen treatment (Pearson Correlation: Pearson |r| = −0.34, P = 0.0035). The expected interaction of virus for combined agents was calculated between N. ceranae and DWV-B in the combined treatments (Nosema-only and Nosema-pollen) and can be considered close to antagonistic due to the calculated negative value −10.58 (equation-5) between the two pathogens (χ2 = 4.905 (equation-3), theoretical χ2 = 3.841, df = 1, P = 0.05). However, the interactive effects of N. ceranae in the combined treatments (Pollen-only, Nosema-only and Nosema-pollen) is close to additive due to the calculated smaller χ2-value (χ2 = 0.025 (equation-3), theoretical χ2 = 3.841, df = 1, P = 0.05).

Correlations between N ceranae spore loads per bee and DWV-B copy numbers per bee in the treatment groups Nosema-only (A) and Nosema-pollen (B) at a log scale. While there was no significant correlation in the Nosema-only treatment (Pearson Correlation, Scatter Plot |r| = −0.22, P = 0.12), a highly significant negative correlation was found in the Nosema-pollen treatment (Pearson Correlation, Scatter Plot |r| = −0.34, P = 0.0035).
The daily sugar water consumption was significantly different between the four treatments (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 2.64, P = 0.017, Fig. 5). A significantly higher sugar water consumption was observed for the Nosema-only group compared to the Pollen-only group (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test * = z > 1.96, P = 0.0089). No significant differences in sugar water consumption were observed between the other groups: (i) Controls and Pollen-only (Kruskal- Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.09), (ii) Controls and Nosema-only (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.79), (iii) Controls and Nosema-pollen (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.1), (iv) Pollen-only and Nosema-pollen (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.3) and (v) Nosema-only and Nosema-pollen (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P = 0.082).

Sugar water consumption in [mg/bee] of individual honey bee workers in the four treatment groups (Controls [white], Pollen-only [orange] Nosema-only [green], Nosema-pollen [blue]). Workers contaminated with N. ceranae from the pollen-only treatment are not considered. Medians, ranges, confidence intervals and outliers ( = dots). Significant differences were found between the Pollen-only and Nosema-only group (*P = 0.0089).
The two pollen-fed treatments (Pollen-only, Nosema-pollen) showed no significant difference in pollen consumption over the 14 days (Kruskal-Wallis multiple comparisons One Way ANOVA, Dunn’s Test z > 1.96, P > 0.05, Fig. 6).

Pollen consumption [mg/bee] of individual honey bee workers in the two pollen treated groups (Pollen-only [orange], Nosema-pollen [blue]). Medians, ranges, confidence intervals and outliers ( = dots) are shown. Workers contaminated with N. ceranae from the pollen-only treatment are not considered. No significant differences were found between the Pollen-only and the Nosema-pollen group (P > 0.05).
Discussion
Our data show for the first time that protein-feeding can have a significant impact on the microsporidian-virus interface in double-infected insect hosts. Taken together with the here confirmed impact on microsporidian8 and virus5,37 infections and on host survival, our results also provide strong support that protein nutrition can functionally alter not only general resistance in insects, but also alter the pattern of host–parasite interactions9.
The experimental pollen was not sterilized similar to other studies8. Therefore, some of the Pollen-only treated workers in our experiment became naturally infected with N. ceranae. These bees were excluded from any further data analyses. Due to possibly associated probiotic microorganisms38, such non-irradiated pollen is probably more beneficial than irradiated one and thus more likely to reveal the full potential of adequate pollen-fed for honey bee resilience. It also appears likely that bees from the Nosema-pollen treatment have obtained additional spores. However, this seems not relevant due to the very high number of N. ceranae spores used for the treatments.
Our data confirm that Nosema spp. infected honey bee workers display higher spore loads when they are pollen-fed (N. apis:6; N. ceranae:7,39,40). This is most likely because Nosema spp. are highly dependent on host nutritional status for their own development, e.g. host amino acids41,42, Adenosine triphosphate (ATP)43,44 or other core nutrients45. Therefore, it appears obvious that any individual host with a pollen-rich diet becomes ideal for Nosema spp. reproduction simply by providing a supreme nutritional environment. However, since protein fed is likely to enhance the immune system5,9, the question emerges, why the bees were not able to reduce the N. ceranae spore load. This might be explained by the lack of host-parasite co-evolution between A. mellifera and the fairly recent invasive species N. ceranae. This scenario seems likely because beekeepers limit natural selection, thereby preventing adaptation of honey bees to this and other novel parasites (i.e. Varroa destructor)46. Alternatively, but not mutually exclusive, N. ceranae might have interfered with the host immune response47. Nevertheless, despite higher spore loads these N. ceranae infected individuals showed an improved survival confirming earlier findings8. Adequate pollen availability might compensate for the energy and nutrients lost in honey bees with high N. ceranae infection intensity, thereby enabling improved survival8. These results are also in line with another study48 that nutrition influences survival in colony level N. ceranae infections. In a simultaneous choice test between sunflower honeys and honeydew N. ceranae infected bees significantly preferred sunflower honey over honey dew49 Such bees consuming sunflower honey showed significantly lower N. ceranae spore loads compared to the honeydew group, probably because of the higher antibiotic activity of sun flower honey49. These findings are well in line with therapeutic self-medication reported from primates50 and butterflies51.
Our data also provide support to the key role of protein nutrition for the outcome of virus infections in insect hosts5,37. Indeed, bees only receiving sugar water showed a significantly higher mortality and higher DWV-B infection levels compared to pollen-fed ones. This confirms that DWV infected bees fed with pollen show lower viral loads than bees only fed with sugar water52. Since just a subsample of 30 bees per treatment was tested for other viruses (BQCV, DWV, ABPV), it can obviously not be excluded that those viruses might also have contributed to a higher bee mortality. However, none of the other viruses were found in any of the analyzed honey bees suggesting a high probability for their absence.
Most interestingly, the results show for the first time that protein-feeding can significantly impact the microsporidian-virus interface in double-infected insect hosts. What are possible reasons for this shifted pathogen interface? In line with other studies30, pre-infection of DWV did not interfere with N. ceranae replication, but N. ceranae did interfere with DWV replication. Since N. ceranae replication in midgut cells disrupts protein metabolism and causes energetic stress8,53,54, this microsporidian is likely to compete with orally acquired viruses for cell resources or for DWV-B’s accessibility to midgut cells30. This could explain why an increase in cell resources due to better protein fed led to higher N. ceranae spore counts, but also resulted in lower virus loads. Moreover, N. ceranae was shown to induce a significant increase in phenol oxidase in bees fed on sugar and workers’ longevity in cages was positively linked to phenol oxidase activity55. This suggests a second possible explanation for the stronger negative correlation between N. ceranae and DWV-B in protein-fed hosts because both phenol oxidase and upregulation of antimicrobial peptides are linked with effective antiviral responses in A. mellifera 56. Since pollen promotes development of the main honey bee immune organ, the fat body57, its maturation may enable a better performance of the immune system including activation of phenol oxidase. Indeed, pollen diet promotes fat body development and enhances survival of N. ceranae parasitized workers that have expressed higher levels of vitellogenin and immunoprotein55,58,59,60,61. Moreover, it is known that pollen supplies are in general immensely important for overwintering honey bee colonies to effectively oppose pathogen stress40. Interestingly, Spaetzle, an activator of the Toll pathway was upregulated by pollen feeding in healthy bees as well as a gene coding for the antimicrobial peptide Defensin and the Peptidoglycan recognition protein PGRP-LC62. In conclusion, the conjunction of increased N. ceranae spore production and enhanced host immunity acquired by proficient protein nutrition most likely explains the observed significant increase in the negative correlation between N. ceranae and DWV-B loads. Alternatively, but not mutually exclusive, the negative correlations between N. ceranae and DWV infections as found in this study and in others29,30,63 can also be explained by individual workers being more susceptible to microsporidians, but less susceptible to virus infections. For example, some bees may be more susceptible to Nosema spp. infections, but could be better able to resist viruses resulting in an overall significant correlation. Indeed, a genetic basis for disease susceptibility is long known in honey bees64, incl. Nosema spp. infections65, and colonies consist of a large number of subfamilies, so-called patrilines, due to the high degree of polyandry by queens66. This provides an alternative explanation for the observed negative correlation between the two pathogens in individual hosts without assuming any antagonistic interactions between them.
No differences in the amount of consumed sugar water were found between control and the Pollen-only group compared to the two N. ceranae -treated groups similar to an earlier study8. The higher amounts of sugar-water consumption by N. ceranae-infected bees observed in other studies67,68,69 may be linked to the lack of protein fed in the respective experimental designs. Our study also revealed a significantly higher sugar water consumption in the Nosema-only group compared to Pollen-only workers, suggesting that lack of protein combined with microsporidian infections may result in higher hunger levels67. In line with an earlier study8, we found no significant differences in the amount of pollen consumed between the Pollen-only and the Nosema-pollen groups. Even though N. ceranae infections certainly induces costs for the hosts, those do apparently not result in significant differences in protein consumption between infected and non-infected bees at least over our experimental period. In conclusion, irrespective of the actual mechanisms underlying the observed stronger negative correlation between microsporidian and virus infection levels in protein-fed hosts, our data strongly suggest that proteins can govern the pathogen-pathogen interface in double-infected insect hosts. Our results further provide support that protein nutrition is an overall key factor for the outcome of infections in insects5,6,7,8,9.
Material and Methods
Study design
The experiment was conducted in September 2014 at the Institute of Zoology, Karl-Franzens University, Graz, Austria using honey bee workers from four randomly chosen queenright local colonies (predominantly A. m. carnica). All colonies were routinely treated against V. destructor in late summer using formic acid and oxalic acid in the previous winter70,71,72.
To test whether pollen nutrition has an effect on N. ceranae interactions with naturally occurring virus infections, a fully-crossed hoarding cage experiment was performed with four replicates each: 1. Workers fed with sugar, but not with pollen (=Controls); 2. Workers fed with both sugar and pollen (=Pollen-only); 3. Workers fed with N. ceranae spores and sugar, but without pollen (=Nosema-only); 4. Workers fed with N. ceranae spores, sugar and pollen (=Nosema-pollen).
Spore solutions
The N. ceranae spore solutions were prepared following routine protocols73. In brief, 12 foragers were collected from the hive entrances of four local infested colonies and dissected. Then, three midguts of N. ceranae infested workers were pooled together in a vial with 0.5 ml water. After homogenization, each vial solution was checked under the light microscope (x400 objective) for the presence of N. ceranae spores74. After all four vials were checked for positive spore loads; the spore solutions were mixed and centrifuged at 5 000 rpm for 5 min. The supernatant containing tissue debris was discarded and the spore pellet was re-suspended in 0.5 ml water by vortexing for 5 sec until spores were uniformly distributed in the solution. This washing step was repeated twice until the N. ceranae spore solution had a concentration of at least 85% purity73. The re-suspended solution was 500 µl water in a 1.5 ml Eppendorf® tube prior to spore load quantification using a haemocytometer and light microscopy (Thermo Fisher Scientific, Waltham, Massachusetts, USA)75,76 focusing on five large squares (each containing 16 small squares) in which the N. ceranae spores were counted. The final concentrations of the spore solutions were quantified using the following calculation77:
where SN is the number of spores per honey bee in 500 µl and SH is the number of spores in 5 large haemocytometer squares (80 small squares). The taxonomic status of the spores was confirmed using N. ceranae species-specific PCR73 for 30 individual honey bees of each treatment.
Experimental set up
Four frames with sealed worker brood were taken from each of the four experimental colonies and placed in an incubator at (34.5 °C) until adult emergence. To ensure that the bees were not older than 24 h, all bees on the brood frames were removed the evening before the experiment started. Each treatment group consisted of six standard hoarding cages with 50 workers randomly assigned to each cage78. Bees were fed with 50% sugar water (w/v) ad libitum until the 3rd day28, at which pollen feeding and N. ceranae infection started. Prior to the treatment, all bees were starved for 2 h29,53,75 before N. ceranae infection was done by bulk feeding73 over 24 h. Workers of the two N. ceranae treatments were challenged with ∼100’000 spores per bee75,78,79,80. All workers were fed until the end of the experiment (14 days) ad libitum with 50% sugar water (w/v). In addition, pollen-treated bees were provided with pollen dough containing corbicula pollen and sucrose candy78. Prior to the experiments, this pollen was not gamma ray irradiated. The experiment lasted 14 d, in which the honey bees were kept in an incubator at brood nest temperature (34.5 °C)81 with 75% RH82, for the first 6 days, before the temperature was decreased to 30 °C78 for the remaining experimental period of 8 days.
To test for potential differences in nutritional demand, pollen dough and sugar water consumption was measured in all cages on a daily basis67. The syringes prepared as feeders were refilled every other day in order to avoid the formation of mold78. Dead bees were removed daily76,83. Possible cage effects were expected as random effects, whereas all replicates had the same conditions in the incubator, temperature and random mixture of bees.
N. ceranae infection levels
Fourteen days post treatment (N. ceranae life cycle = 14 days73 the experiment was terminated and the surviving individuals (N = 401) were separately freeze-killed and stored at −80 °C until further analyses. To test for N. ceranae infection levels, the workers were crushed in 2 ml microcentrifuge tubes containing a 5 mm metal beads and 200 µl TN-buffer (1 M Tris; 1 M NaCl) for 30 sec at 25 shakes per sec. Spore counts and calculations were performed as described above.
Virus infection levels
All 401 bees were individually analyzed for DWV-B. Prior to RNA extraction, individual bees were crushed in 2 ml microcentrifuge tubes containing 5 mm metal beads and 200 µl TN-buffer (10 mM Tris, 10 mM NaCl; pH 7.6). The samples were homogenized with a tissue-lyser for 30 sec at 25 1/s frequency using a Qiagen Retsch® MM 300 mixer mill84. Then, the homogenates were centrifuged at 2500 rpm and 50 µl of the supernatant was destined for total RNA extraction using NucleoSpin RNA extraction kit (Macherey-Nagel) following the manufactures guidelines. Reverse transcription was performed using 2 µg of extracted RNA incubated with random hexamer primers for 5 min at 70 °C. Then mixed with 5 µl of 5x buffer, 1.25 µl dNTP (10 mM) and 1 µl M-MLV before incubating the 25 µl volume reaction for 60 min at 37 °C. For virus quantification, 10-fold diluted cDNA was mixed with Kapa SYBR® FAST qPCR Master Mix kit. Briefly, 6 µl 2x reaction buffer, 0.24 µl forward and reverse primers for DWV-B and β-actin (Table 1)85 merged with 2.52 µl water and 3 µl template in a total of 12 µl final reaction volume86. The real-time qPCR cycling profile consisted of 3 min incubation at 95 °C and 40 cycles of 3 sec at 95 °C for denaturation, 30 sec at 57 °C for annealing and data collection. The melting-curve analysis was performed with the following conditions: 15 s at 95 °C, 55 °C and 95 °C, respectively. Purified DWV-B PCR products of know concentration (10−3–10−6 ng) were used as standard curves on each individual plate, along with non-template controls (R2: 0.992; Slope: −3.198; Intercept: 30.240; PCR efficiency: 2.054). Quantification of the β-actin gene was performed in parallel for each sample as reference gene for DWV-B normalization85. A Cq cut-off value (according to the value of the negative control) was used to define the disease status (positive or negative). The ECO Software real-time PCR system (Illumina, San Diego, CA, USA) was used to evaluate the performance of the qPCR reactions and to analyze the qPCR quantification. These Cq-values were used to calculate the virus infection levels in Log [copies/bees] which were then used for the statistical analyses.
A further subsample of 30 bees per treatment (N = 5 each cage) was screened to determine if other honey bee viruses were also present in the colonies during the experiment: BQCV, which is known to be associated with N apis infection31,32 and occurs in 30% of Austrian honey bees87. Three more viruses associated with honey bees in Austria were screened: DWV which is present in 91% of Austrian honey bees including its variant DWV-B and ABPV reported to be present in 68% of Austrian bees87.
Statistical analyses
Data were tested for normality using Shapiro-Wilk’s test (P > 0.05). If, however, normality was rejected (Shapiro-Wilk’s test, P < 0.05), groups were compared by performing non-parametric Kruskal-Wallis multiple comparison One Way ANOVAs (Dunn’s test) and Pearson correlation.
Longevity analyses for the four individual treatment groups were conducted by using Kaplan-Meier Survival Curves and a Log-Rank assessment.
Interactions between treatments on worker mortality were determined by using χ2 tests88,89. The expected interaction mortality value, ME for combined treatment was calculated using the following formula:
where MPT and MN are the observed percent mortalities caused by pollen treatment and N. ceranae infection.
The resulting values from each equation were then compared to the χ2 table value with 1 df, using the formula:
where M O is the observed mortality for the combined N. ceranae with pollen treatment.
A non-additive effect between the two agents was expected when the χ2 value exceeded the given table value. If, however the difference between
or
had a positive or a negative value, an interaction was then regarded as being synergistic or antagonistic, respectively88.
Synergistic, additive, or antagonistic interactions between agents in the combination treatments for DWV-B loads and N. ceranae spores were determined using a χ2 test90,91,92,93.
Comparisons of N. ceranae spore loads, DWV-B infection levels and sugar/pollen consumption rates in the different treatment groups were performed using Kruskal-Wallis One Way ANOVAs (and multiple comparisons, Dunn’s Test).
Pearson correlations between N. ceranae spore loads and DWV-B infections levels (Log [copies/bee]) were performed for both Nosema-only and Nosema-pollen groups.
All statistical analyses were performed using the program NCSS (NCSS 9 Statistical Analysis and Graphics).
References
Ponton, F. et al. Integrating nutrition and immunology: A new frontier. J. Insect Physiol. 59, 130–137 (2012).
Cunningham-Rundles, S., McNeeley, D. F. & Moon, A. Mechanisms of nutrient modulation of the immune response. J. Allergy Clin. Immunol. 115, 1119–1129 (2005).
Ponton, F., Wilson, K., Cotter, S. C., Raubenheimer, D. & Simpson, S. J. Nutritional immunology: A multi-dimensional approach. PLoS Pathog. 7, 1–4 (2011).
De Grandi-Hoffman, G. & Chen, Y. Nutrition, immunity and viral infections in honey bees. Curr. Opin. Insect Sci. 10, 170–176 (2015).
Lee, K. P., Cory, J. S., Wilson, K., Raubenheimer, D. & Simpson, S. J. Flexible diet choice offsets protein costs of pathogen resistance in a caterpillar. Proc. R. Soc. B 273, 823–829 (2006).
Povey, S., Cotter, S. C., Simpson, S. J., Lee, K. P. & Wilson, K. Can the protein costs of bacterial resistance be offset by altered feeding behaviour? J. Anim. Ecol. 78, 437–446 (2009).
Rinderer, T. E. & Elliott, K. D. Worker honey bee response to infection with Nosema apis: Influence of diet. J. Econ. Entomol. 70, 431–433 (1977).
Jack, C. J., Uppala, S. S., Lucas, H. M. & Sagili, R. R. Effects of pollen dilution on infection of Nosema ceranae in honey bees. 87, 12–19 (2016).
Brunner, F. S., Schmid-Hempel, P. & Barribeau, S. M. Protein-poor diet reduces host-specific immune gene expression in Bombus terrestris. Proc. R. Soc. B 281, 20140128 (2014).
Woolhouse, M. E. J., Webster, J. P., Domingo, E., Charlesworth, B. & Levin, B. R. Biological and biomedical implications of the co-evolution of pathogens and their hosts. Nat. Genet. 32, 569–577 (2002).
Bordes, F. & Morand, S. The impact of multiple infections on wild animal hosts: a review. Infect. Ecol. Epidemiol. 1, 1–10 (2011).
Rau, D. et al. Co-evolution in a landrace meta-population: two closely related pathogens interacting with the same host can lead to different adaptive outcomes. Sci. Rep. 5, 12834 (2015).
Olive, A. J. & Sassetti, C. M. Metabolic crosstalk between host and pathogen: sensing, adapting and competing. Nat. Rev. Microbiol. 14, 221–234 (2016).
Potts, S. G. et al. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353 (2010).
Dainat, B., Evans, J. D., Chen, Y. P., Gauthier, L. & Neumann, P. Predictive markers of honey bee colony collapse. PLoS One 7, e32151 (2012).
Ellis, J. D. & Munn, P. A. The worldwide health status of honey bees. Bee World 86, 88–101 (2005).
Dobson, A. The population dynamics of competition between parasites. Parasitology 91, 317–347 (1985).
Poulin, R. Interactions between species and the structure of helminth communities. Parasitology 122, 3–11 (2001).
Read, A. F. & Taylor, L. H. The ecology of genetically diverse infections. Science 292, 1099–1102 (2001).
West, S. A. & Buckling, A. Cooperation, virulence and siderophore production in bacterial parasites. Proc. Biol. Sci. 270, 37–44 (2003).
Griffin, A. S., West, S. A. & Buckling, A. Cooperation and competition in pathogenic bacteria. Earth 430, 1024–1027 (2004).
Alizon, S. & Lion, S. Within-host parasite cooperation and the evolution of virulence. Proc. R. Soc. B 278, 3738–3747 (2011).
Ongus, J. R. et al. Complete sequence of a picorna-like virus of the genus Iflavirus replicating in the mite Varroa destructor. J. Gen. Virol. 85, 3747–3755 (2004).
Shen, M., Cui, L., Ostiguy, N. & Cox-Foster, D. Intricate transmission routes and interactions between picorna-like viruses (Kashmir bee virus and sacbrood virus) with the honeybee host and the parasitic varroa mite. J. Gen. Virol. 86, 2281–2289 (2005).
Yue, C. & Genersch, E. RT-PCR analysis of Deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J. Gen. Virol. 86, 3419–3424 (2005).
Neumann, P., Yañez, O., Fries, I. & De Miranda, J. R. Varroa invasion and virus adaptation. Trends Parasitol. 28, 353–354 (2012).
Ball, B. V. & Allen, M. F. The prevalence of pathogens in honey bee (Apis mellifera) colonies infested with the parasitic mite Varroa jacobsoni. Ann. Appl. Biol. 113, 237–244 (1988).
Natsopoulou, M. E., Mc Mahon, D. P., Doublet, V., Bryden, J. & Paxton, R. J. Interspecific competition in honeybee intracellular gut parasites is asymmetric and favours the spread of an emerging infectious disease. Proc. R. Soc. B 282, 20141896 (2014).
Costa, C., Tanner, G., Lodesani, M., Maistrello, L. & Neumann, P. Negative correlation between Nosema ceranae spore loads and deformed wing virus infection levels in adult honey bee workers. J. Invertebr. Pathol. 108, 224–225 (2011).
Doublet, V., Natsopoulou, M. E., Zschiesche, L. & Paxton, R. J. Within-host competition among the honey bees pathogens Nosema ceranae and Deformed wing virus is asymmetric and to the disadvantage of the virus. J. Invertebr. Pathol. 124, 31–34 (2015).
Bailey, B. Y. L., Ball, B. V. & Perry, J. N. Association of viruses with two protozoal pathogens of the honey bee. Ann. appl. Biol. 103, 13–20 (1983).
Allen, M. & Ball, B. The incidence and world distribution of honey bee viruses. Bee World 77, 141–162 (1996).
Cox-Foster, D. L. et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science. 318, 283–287 (2007).
Haydak, M. H. Honey bee nutrition. Annu. Rev. Entomol. 15, 143–156 (1970).
Brodschneider, R. & Crailsheim, K. Nutrition and health in honey bees. Apidologie 41, 278–294 (2010).
Mc Mahon, D. P. et al. A sting in the spit: widespread cross-infection of multiple RNA viruses across wild and managed bees. J. Anim. Ecol. 84, 615–624 (2015).
Porrini, M. P. et al. Nosema ceranae development in Apis mellifera: influence of diet and infective inoculum. J. Apic. Res. 50, 35–41 (2011).
Engel, P. et al. The Bee Microbiome: Impact on Bee Health and Model for Evolution and Ecology of Host-Microbe Interactions. mBio 7, 1–9 (2016).
Rinderer, T. E. & Sylvester, A.H. Variation in response to Nosema apis longevity and hoarding behaviour in a free mating population of honey bee. Entomol. Soc. Am. 372–374 (1977).
Naug, D. Nutritional stress due to habitat loss may explain recent honeybee colony collapses. Biol. Conserv. 142, 2369–2372 (2009).
Wang, D. & Moeller, F. Comparison of the free amino acid composition in the hemolymph of healthy and -infected female honey bees. J. Invertebr. Pathol. 15, 202–206 (1970).
Panek, J. et al. Hijacking of host cellular functions by an intracellular parasite, the microsporidian Anncaliia algerae. PLoS One 9, (2014).
Weidner, E., Findley, A. M., Dolgikh, V. & Sokolova, J. Microsporidian biochemistry and physiology. Am. Soc. Microbiol. 89, 209–214 (1999).
Williams, B. A. P. Unique physiology of host-parasite interactions in microsporidia infections. Cell. Microbiol. 11, 1551–1560 (2009).
Weiss, L. M. & Becnel, J. J. Microsporidia: Pathogens of Opportunity. Wiley Blackwell, Hoboken, NJ. (2014).
Neumann, P. & Blacquiére, T. The Darwin cure for apiculture? Natural selection and managed honeybee health. Evol. Appl. 10, 226–230 (2017).
Antúnez, K. et al. Immune suppression in the honey bee (Apis mellifera) following infection by Nosema ceranae (Microsporidia). 11, 2284–2290 (2009).
Eischen, F. A. & Graham, R. H. Feeding overwintering honey bee colonies infected with Nosema ceranae. Am. Bee J. 148, 555 (2008).
Gherman, B. I. et al. Pathogen-associated self-medication behavior in the honeybee Apis mellifera. Behav. Ecol. Sociobiol. 68, 1777–1784 (2014).
Lozano, G. A. Parasitic Stress and Self-Medication in Wild Animals. Adv. Study Behav. 27, 291–317 (1998).
Sternberg, E. D. et al. Food Plant-Derived Disease Tolerance and Resistance in a Natural Butterfly - Plant-Parasite Interactions. Evolution. 66, 3367–3377 (2012).
Degrandi-Hoffman, G., Chen, Y., Huang, E. & Hua, M. The effect of diet on protein concentration, hypopharyngeal gland development and virus load in worker honey bees (Apis mellifera L.). J. Insect Physiol. 56, 1184–1191 (2010).
Higes, M., García-Palencia, P., Martín-Hernández, R. & Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 94, 211–217 (2007).
Holt, H. L., Aronstein, K. A. & Grozinger, C. M. Chronic parasitization by Nosema microsporidia causes global expression changes in core nutritional, metabolic and behavioral pathways in honey bee workers (Apis mellifera). BMC Genomics 14, 1–16 (2013).
Di Pasquale, G. et al. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS One 8, e72016 (2013).
Brutscher, L. M., Daughenbaugh, K. F. & Flenniken, M. L. Antiviral defense mechanisms in honey bees. 21, 193–201 (2016).
Alaux, C. et al. Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environ. Microbiol. 12, 774–782 (2010).
Alaux, C., Ducloz, F., Crauser, D. & Le Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 6, 562–565 (2010).
Amdam, G. V. et al. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 39, 767–773 (2004).
Zheng, H. et al. Spore Loads May Not be Used Alone as a Direct Indicator of the Severity of Nosema ceranae Infection in Honey Bees Apis mellifera (Hymenoptera: Apidae). J. Econ. Entomol. 107, 2037–2044 (2014).
Lee, K. P., Simpson, S. J. & Wilson, K. Dietary protein-quality influences melanization and immune function in an insect. Funct. Ecol. 22, 1052–1061 (2008).
Alaux, C., Dantec, C., Parrinello, H. & Conte, Y. L. Nutrigenomics in honey bees: digital gene expression analysis of pollen’s nutritive effects on healthy and varroa-parasitized bees. BMC Genomics 496, 1–13 (2011).
Martin, S. J. et al. Do the honeybee pathogens Nosema ceranae and deformed wing virus act synergistically? Environ. Microbiol. Rep. 5, 506–510 (2013).
Rothenbuhler, W. C. Behavior Genetics of Nest Cleaning in Honey Bees Responses of F1 and Backcross Generations to Disease-Killed Brood. Am. Zool 4, 111–123 (1964).
Huang, Q., Kryger, P., Le Conte, Y. & Moritz, R. F. A. Survival and immune response of drones of a Nosemosis tolerant honey bee strain towards N. ceranae infections. J. Invertebr. Pathol. 109, 297–302 (2012).
Neumann, P., Moritz, R. F. A. & Praagh, J. P. V. Queen mating frequency in different types of honey bee mating apiaries. J. Apic. Res. 38, 11–18 (1999).
Mayack, C. & Naug, D. Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J. Invertebr. Pathol. 100, 185–188 (2009).
Naug, D. & Gibbs, A. Behavioral changes mediated by hunger in honeybees infected with Nosema ceranae. Apidologie 40, 595–599 (2009).
Martín-Hernández, R. et al. Comparison of the energetic stress associated with experimental Nosema ceranae and Nosema apis infection of honeybees (Apis mellifera). Parasitol. Res. 109, 605–612 (2011).
Charrière, J. & Imdorf, A. Oxalic acid treatment by trickling against Varroa destructor: recommendations for use in central Europe and under temperate climate. Bee World 82, 51–60 (2002).
Godfray, H. C. J. et al. A restatement of the natural science evidence base concerning neonicotinoid insecticides and insect pollinators. Proc. R. Soc. B 281, 20140558 (2014).
Dietemann, V. et al. Standard methods for varroa research. J. Apic. Res. 52, 1–54 (2013).
Fries, I. et al. Standard methods for nosema research. J. Apic. Res. 52, 1–28 (2013).
Cantwell, G. E. Standard methods for counting Nosema spores. 110, 222–223 (1970).
Retschnig, G., Neumann, P. & Williams, G. R. Thiacloprid – Nosema ceranae interactions in honey bees: Host survivorship but not parasite reproduction is dependent on pesticide dose. J. Invertebr. Pathol. 118, 18–19 (2014).
Williams, G. R., Shutler, D., Burgher-Mac Lellan, K. L. & Rogers, R. E. L. Infra-Population and - Community Dynamics of the Parasites Nosema apis and Nosema ceranae, and Consequences for Honey Bee (Apis mellifera) Hosts. 9, 5–10 (2014).
Hornitzky, M. Nosema diagnosis. Dep. Prim. Ind. 1, 1–2 (2009).
Williams, G. R. et al. Standard methods for maintaining adult Apis mellifera in cages under in vitro laboratory conditions. J. Apic. Res. 52, 1–36 (2013).
Groh, C., Tautz, J. & Rössler, W. Synaptic organization in the adult honey bee brain is influenced by brood-temperature control during pupal development. Proc. Natl. Acad. Sci. USA 101, 4268–4273 (2004).
Wolf, S. et al. So Near and Yet So Far: Harmonic Radar Reveals Reduced Homing Ability of Nosema Infected Honeybees. PLoS One 1–15 (2014).
Crailsheim, K. et al. Standard methods for artificial rearing of Apis mellifera larvae. J. Apic. Res. 52, 1–16 (2013).
Ellis, M. B., Nicolson, S. W., Crewe, R. M. & Dietemann, V. Hygropreference and brood care in the honeybee (Apis mellifera). J. Insect Physiol. 54, 1516–1521 (2008).
Paxton, R. J., Klee, J., Korpela, S. & Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 38, 558–565 (2007).
Evans, J. D. et al. Standard methods for molecular research in Apis mellifera. J. Apic. Res. 52, 1–54 (2013).
Gauthier, L. et al. Viruses Associated with Ovarian Degeneration in Apis mellifera L. Queens. PLoS One 6, 1–13 (2011).
de Miranda, J. R. et al. Standard methods for virus research in Apis mellifera. J. Apic. Res. 52, 1–56 (2013).
Berenyi, O., Bakonyi, T., Derakhshifar, I., Köglberger, H. & Nowotny, N. Occurrence of Six Honeybee Viruses in Diseased Austrian Apiaries. Appl. Environ. Microbiol. 72, 2414–2420 (2006).
Morales-Rodriguez, A. & Peck, D. C. Synergies between biological and neonicotinoid insecticides for the curative control of the white grubs Amphimallon majale and Popillia japonica. Biol. Control 51, 169–180 (2009).
Aufauvre, J. et al. Parasite-insecticide interactions: a case study of Nosema ceranae and fipronil synergy on honeybee. Sci. Rep. 2, 1–7 (2012).
Mc Vay, J. R., Gudauskas, R. T. & Harper, J. D. Effects of Bacillus thuringiensis Nuclear-Polyhedrosis Virus Mixtures on Trichoplusia ni Larvae. J. Invertebr. Pathol. 29, 367–372 (1977).
Koppenhöfer, A. M. & Kaya, H. K. Synergism of imidacloprid and an entomopathogenic nematode: a novel approach to white grub (Coleoptera: Scarabaeidae) control in turfgrass. J. Econ. Entomol. 91, 618–623 (1998).
Koppenhöfer, A. M. & Fuzy, E. M. Steinernema scarabaei for the control of white grubs. Biol. Control 28, 47–59 (2003).
Koella, J. C. & Sørensen, F. L. Effect of adult nutrition on the melanization immune response of the malaria vector Anopheles stephensi. Med. Vet. Entomol. 16, 316–320 (2002).
Locke, B., Forsgren, E., Fries, I. & de Miranda, J. Acaricide Treatment Affects Viral Dynamics in Varroa destructor - Infested Honey Bee Colonies via both Host Physiology and Mite Control. Appl. Environ. Microbiol. 78, 227–235 (2012).
Acknowledgements
The authors are grateful to Elodie Cristina, Kaspar Roth and Lars Straub for technical support and to Lars Straub for linguistic assistance. Financial support was granted to PN by the Vinetum Foundation and the Ricola Foundation Nature and Culture.
Author information
Authors and Affiliations
Contributions
P.N. and K.C. conceived the study and designed the experiment; M.T., J.V. and O.Y. carried out the experiment; M.T. and O.Y. analysed the data and prepared figures; P.N., M.T., K.C. and N.C. wrote the manuscript; all authors edited the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tritschler, M., Vollmann, J.J., Yañez, O. et al. Protein nutrition governs within-host race of honey bee pathogens. Sci Rep 7, 14988 (2017). https://doi.org/10.1038/s41598-017-15358-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-15358-w
This article is cited by
-
Geographical and Seasonal Analysis of the Honeybee Microbiome
Microbial Ecology (2023)
-
Effects of different artificial diets on commercial honey bee colony performance, health biomarkers, and gut microbiota
BMC Veterinary Research (2022)
-
You are what you eat: relative importance of diet, gut microbiota and nestmates for honey bee, Apis mellifera, worker health
Apidologie (2021)
-
From antagonism to synergism: Extreme differences in stressor interactions in one species
Scientific Reports (2020)
-
Can supplementary pollen feeding reduce varroa mite and virus levels and improve honey bee colony survival?
Experimental and Applied Acarology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
