
Absract
The most anthropized regions of the world are characterized by an impressive abundance of invasive plants, which alter local biodiversity and ecosystem services. An alternative strategy to manage these species could be based on the exploitation of their fruits in a framework of bioprospecting to obtain high-added value compounds or phytocomplexes that are useful for humans. Here we tested this hypothesis on three invasive plants (Lonicera japonica Thunb., Phytolacca americana L., and Prunus serotina Ehrh.) in the Po plain (northern Italy) which bear fruits that are highly consumed by frugivorous birds and therefore dispersed over large distances. Our biochemical analyses revealed that unripe fruit shows high antioxidant properties due to the presence of several classes of polyphenols, which have a high benchmark value on the market. Fruit collection for phytochemical extraction could really prevent seed dispersal mediated by frugivorous animals and produce economic gains to support local management actions.
Similar content being viewed by others

Introduction
Biological invasions constitute a global environmental, economic, and social change driver as they can irremediably alter ecosystem compositions and services, thus determining a loss in terms of biodiversity, agriculture productivity, and human health1,2. In Europe, the number of invasive species has been rapidly increasing and reached an estimated value of over 12000 species including plants, animals, fungi, and microorganisms in 20163. Concerning plants, their intentional introduction for ornamental purposes is a major pathway for invasion4. The effect of invasive plants on natural ecosystems can be disastrous, firstly for the competition with indigenous species5, and secondly, because they limit the stability and availability of natural resources to the whole native species community6,7. Detrimental effects on human health mediated by invasive plants have also been documented2. These include the occurrence of immunostimulant substances or allergenic factors, such as those on the pollen of some species or toxic metabolites in fruits, leaves, or other plant portions8. A typical example is Ambrosia artemisiifolia L. and A. trifida L. which pollens induce respiratory diseases, and management costs to avoid allergy outbreaks are very high9.
In the attempt to contrast the risks posed by alien and invasive plant species, management actions have been implemented to prevent introductions4, eradicate infestations before they become established10,11, and contain the spread of abundant or resistant taxa4,12,13,14. In many cases, management strategies (e.g., cutting, mowing and extirpation) are not completely effective, either for eradication or containment purposes, because invasive species rely on efficient dispersal systems ensuring their long-term persistence. Moreover, these strategies demand huge economic resources, not only to assist direct containment, but also for the management of the biomass resulting from extirpation and cutting activities15.
Considering that climate change is likely to exacerbate the negative effects of invasive alien species, as it will foster their further spread especially in the most fertile areas9, the economic resources and human energy demanded for managing invasive plants are expected to increase. The engagement of the citizen and other stakeholders to improve actions aimed at controlling the invaders creates societal challenges that policy makers are asked to prioritize16. At the same time, identifying innovative ways to transform invasive plants from ‘problems to resources’ also represent an important challenge for modern societies. In this context, we propose a bioprospecting strategy to find new compounds and antioxidant phytocomplexes to be used in different sectors of human interest (e.g. pharmaceutical, cosmetic, and nutrition). Usually, bioprospecting activities involve wild species from unexplored forests17, however, invasive species could also hide interesting chemical profiles or biological activities.
The technical goal of this study was to explore the chemical and biological properties of fruit from three invasive plants; Lonicera japonica Thunb., Phytolacca americana L., and Prunus serotina Ehrh. These species produce a large amount of fleshy fruits that are rich in secondary metabolites. Moreover, seeds of these plants represent the main tool for plant invasion and therefore, their collection and disposal for bioprospecting aims, could represent a suitable and innovative strategy for containment purposes.
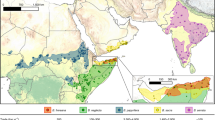
All the three selected invasive species are very abundant in the Po plain region of Northern Italy. Like other European floodplains, this geographical area is increasingly colonized by invasive species due to the huge amount of water availability, fertile soils, and suitable climatic conditions18. The Po plain is characterized by high human disturbance resulting in a consistent alteration of local floral communities and an increasing rate of habitat fragmentation; these conditions support the process of invasion19. In addition, the Po plain is crossed by important migration routes (Fig. 1) and contains many refueling stopover sites for migratory birds species crossing the Alpine barrier to reach breeding sites farther north in spring and wintering grounds farther south in autumn20. Considering that the fruit from L. japonica, P. americana, and P. serotina are highly appreciated by frugivorous bird species, these last represent the primary vector for the diffusion and settlement of fleshy-fruiting plants20,21,22. For these reasons, the selection of efficient management activities to contrast the diffusion of invasive plant species represents a priority issue, not only to preserve local biodiversity but also to prevent the spread of these species in bird stopover areas along transcontinental migratory routes.

Study area. Location of the Italian Po Plain in the Western Palearctic region (upper right box) and sampling sites (1: Cassinazza di Baselica; 2: Valle Lambro Park; 3: Ticino Park). Mountain chains are indicated in light gray, whereas the parks investigated in this study are indicated in dark gray. Arrows represent some of the main bird migration routes crossing the Po plain region. The map has been generated using QGis desktop 2.18.3 and modified with Adobe Photoshop CC. The source map was downloaded from the public dataset (http://thematicmapping.org/downloads/world_borders.php) licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License (https://creativecommons.org/licenses/by-sa/3.0/).
The rationale of the present study is to attract the general public and industry (i.e. pharmaceutical, cosmetic, and food) attention on these plants and their fruits in order to induce a massive exploitation for the extraction and valorization of their phytochemicals, which have beneficial effects on human health. This strategy could contribute to preventing the zoochorous dispersal of invasive seeds, while also producing economic benefits to support biodiversity management.
Methods
Study area and investigated plant species
The study area selected for sampling invasive plants was in the Lombardy Region and specifically, the natural areas of the Po plain (Fig. 1).
Parks and reserves of this floodplain are full of invasive plants (i.e., about 619 aliens out of 3820 plant recorded species)19 and include a lot of suitable stopover sites exploited by millions of frugivorous birds crossing the Alps during fall migration. These areas are also subjected to a progressive erosion of local plant diversity due to the consistent spread of invasive species20. Among alien taxa inhabiting the Po plain, L. japonica, P. americana, and P. serotina represent the most invasive species.
L. japonica is a Japanese plant introduced in Europe for ornamental purposes during the 19th century. In the Po plain, this species colonized different habitats, particularly mesophyll degraded woods and floodplains. This species is very attractive to birds, which feed on its fruit and contribute to its dispersal23. It produces about 90 seeds per plant and it is able to propagate by exploiting an efficient vegetative reproductive strategy and reaching sexual maturity within one year after settlement24. For these reasons, this species is included in the Black List of exotic invasive species in Lombardy19.
P. americana is another North American species introduced in Europe since the 16th century and is considered by some authors as naturalized in many European countries19. Nevertheless, it outcompetes local flora, since its numerous seeds (i.e. about 1500 seeds per plant) are scattered by a wide panel of frugivorous mammals and birds25.
P. serotina is a North American species that was introduced in Italy in 1922. This infesting plant is especially spreading in floodplain areas and poses a great risk to local flora due to its allelopathic strategies that change the chemical and microbial composition of the soil while also limiting the germination of seeds from local species19. Sexually mature plants produce an average of 6000 seeds each26, which are diffused mainly by frugivorous bird species27.
Collection of plant material
For each species, 100 unripe and ripe fruits respectively, were sampled in natural areas of the Lombardy Po plain (Fig. 1). Specifically, L. japonica was collected in the Southern part of the Lombardy Po plain at the naturalized wet area of Cassinazza di Baselica (45°12′16.1″N, 8°36′00.7″E). P. americana was sampled in the Park of Valle Lambro (45°40′11.8″N, 9°15′23.2″E), and P. serotina was collected in the Ticino Park in the North-West Lombardy (45°40′37.9″N, 8°41′02.6″E).
Fruit from Vaccinium myrtillus L. were used as a standard reference for chemical analyses due to its high concentration of antioxidant compounds28. V. myrtillus was collected in Northern Lombardy. (45°59′00.0″N, 9°24′00.0″E).
Fruits were immediately stored at −20 °C after collection until laboratory analyses. Chemical composition and antioxidant activity analyses were performed on both unripe and ripe fruits of the three invasive plant species and of the control species as well.
Fruit extracts
For each species and for both categories (i.e. ripe and unripe), fruits were further processed using liquid nitrogen and were powdered. A total of 3 g of material was subjected to two different extraction procedures: hydro-alcoholic and aqueous. The hydro-alcoholic procedure was performed by following the method proposed by Amigoni and co-workers29 without filtering with 0.45 μm PTFE filters and with a modified drug/solvent ratio: 3 g of powder were extracted using 30 mL of 70% (v/v) ethanol. The aqueous extraction was performed following the protocol described by Zhu and co-workers30. Both extractions for each species/fruit category were performed three times. After extraction, solvents were evaporated by a rotary evaporator and then lyophilized. Dry extracts were stored at −20 °C until analyses.
Assessment of antioxidant activity
To evaluate the antioxidant properties of plant extracts, the DPPH assay31 was performed. Due to the sensitivity of this test to acid and basic pH values32, it was verified that the pH of extracts ranged from 4 to 8. Values obtained from unripe and ripe fruit extracts were compared with the extracts of the control species V. myrtillus. Moreover, the Folin-Ciocalteu assay33 was also performed to verify whether the antioxidant activity of the extracts could be related to the presence of polyphenols. Results of the DPPH and Folin-Ciocalteau assays were expressed as Trolox and gallic acid (GA) equivalents respectively, by means of calibration curves.
Characterization of phytocomplexes
The obtained fruit extracts were characterized spectrophotometrically for the specific assessment of total flavonoids33,34, catechins35, and hydroxycinnamic acids36. These are colorimetric assays based on the reaction between a specific class of phytochemical and the specific reagents. A calibration curve is prepared for each assay by using increasing concentrations of a standard compound representative of the class: catechin (CAT) for flavonoids and catechins and ferulic acid (FA) for hydroxycinnamic acids. The color intensity of reacted samples was measured by a spectrophotometer and the concentration of the searched phytochemical family was calculated by means of the calibration curve. The amount of tannins was analyzed according to Porter and co-workers protocol37 with minor modifications; briefly, two aliquots of each sample were mixed with the reagent (50% n-butanol, 50% 12 N HCl, 0.015% FeCl3) and incubated for 30 min, one at room temperature and the second at 100 °C. Absorbance (Abs) was measured at 550 nm and tannin content (g/L) was calculated as (Abs100 °C−Absroom temperature) × 0.1736.
Quantification of specific polyphenols by HPLC-DAD
Plant fruit extracts were analyzed for phenolic compounds by HPLC-DAD38. Samples were loaded onto a Strata-X column (33 mm polymeric sorbent 60 mg in 3 mL, Phenomenex, Torrence, CA, USA), and polyphenols were eluted by 100% v/v methanol. The eluates were completely dried in a speed vacuum at 45 °C and resuspended in 200 mL of 1:9 acetonitrile/0.2% v/v acetic acid before being directly injected into the HPLC. Polyphenols were analyzed by RP HPLC separation (column Gemini C18, 5 µm particles 250 × 4.60 mm, precolumn SecurityGuard Ea, Phenomenex, Torrence, CA, USA) equipped with an on-line diode array detector (MD-2010, Plus, Jasco Instruments, Großumstad, Germany) analyzing from 220 to 550 nm.
The adopted HPLC-DAD separation procedure allowed for the simultaneous analysis of 28 compounds among which stilbenes, phenolic acids, and flavonoids: trans- and cis-resveratrol (tRES, cRES), trans- and cis-piceid (tPIC, cPIC), trans- and cis-resveratroloside (tRDE, cRDE), piceatannol (PICEAT); gallic, protocatechuic, syringic, and vanillic acids (GA, PROTA, SIRA, VANA); caffeic, chlorogenic, p-coumaric, ferulic, sinapic, and trans-cinnamic acids (CAFA, CLORA, CUMA, FA, SINA, tCINA); catechin (CAT), epicatechin (EC), epigallocatechin 3-O-gallate (EGCG), epicatechin gallate (ECG), epigallocatechin (EGC); vanillin (VAN), naringenin (NAR), quercetin (QUERC), rutin (RUT), myricetin (MYR), and kaempferol (KAE). Based on SciFinder (https://scifinder.cas.org/), the average market price of these metabolites was assessed and used to evaluate the potential chemical value of the analyzed fruits.
Seed viability
To estimate the invasive potential of each species through seed dispersal, the presence of mature seeds and their viability starting from unripe and ripe fruits was investigated. A total of 50 seeds per species belonging to 10 different plants were analyzed using the tetrazolium chloride test to verify the respiration of the embryos through a chemical reaction that produces a reddish stain21. Negative controls were performed on dead seeds obtained by heating fresh seeds to 100 °C for 1 h.
Statistical analyses
Extraction yields data were log transformed and then analyzed by one way ANOVA with the Welch correction due to their non-homogenous variances. The Duncan test was performed to identify the samples significantly different from the others. Antioxidant activity and total phenol content of the samples were compared to those obtained from V. myrtillus extracts using the Neyman method of confidence interval at 95% level. Then to verify whether the antioxidant activity was due to the presence of polyphenols, to the degree of ripening, to the solvent of extraction, and to understand whether it could be of interest to better characterize polyphenols, we analyzed our data using linear models (LM) or, if necessary, using the generalized least squares method (GLS). Data concerning the chemical characterization of flavonoids, catechins, hydroxicinnamic acids, and tannins were analyzed as follows. Normality was verified using the Shapiro-Wilk test and homogeneity of variances was verified by Levene test. Data lacking normality were transformed by i) log transformation (flavonols), ii) square root transformation (hydroxicinnamic acids, tannins), and iii) rank transformation (catechins). One-way analysis of variance (ANOVA) was performed on transformed data (in the case of heteroscedasticity, the Welch’s correction was applied), and then data were analyzed by the Tukey-Kramer post hoc test (if the assumption of homogeneity of variances was satisfied) or by the Duncan’s multiple range test (if the assumption of homogeneity of variances was not satisfied) at a significance level of 0.05. All the analyses and graphics were produced using the Software R 1.0.44.
Data availability
All the data supporting the results presented here have been provided within the manuscript’s tables or as Supplementary material.
Results
Yields of extraction and antioxidant activity
Figure 2 provides a brief comparison of extraction yields for all the investigated species and tested protocols. P. americana showed the highest yield of extraction in all cases with the only exception of unripe fruits extracted with water. L. japonica showed a statistically lower yield of extraction than the control V. myrtillus in the hydro-alcoholic condition, and the opposite situation occurred with the aqueous extraction protocol. Finally, P. serotina extraction yields starting from unripe fruits were lower than in the ripe ones, even though yields of aqueous extraction were comparable to that of V. myrtillus.

Yields of the hydro-alcoholic (A) and aqueous (B) extractions conducted on fruits from the three invasive plant species and the control species V. myrtillus. Data are reported as mg of dry powder obtained from 1 g of fresh fruit and expressed as the mean ± standard deviation (SD). The letters above each bar indicate the results of Duncan’s multiple range test (p < 0.05). Average values with standard deviation followed by the same letters are not significantly different.
Antioxidant activity values estimated by the DPPH assay are shown in Fig. 3(A,B). The highest value was observed for P. serotina unripe fruit, where both hydro-alcoholic and aqueous extracts showed to be 1.6 times more active than V. myrtillus unripe extracts. When fruits ripen, the statistical difference between the antioxidant activity of P. serotina extracts and that of the control species disappears, and ripe P. serotina fruits are generally two times less active that unripe ones. Phytolacca americana and L. japonica showed values of antioxidant activity quite lower than V. myrtillus, and these species also show a higher antioxidant activity in unripe conditions.

Results of DPPH (A,B) and Folin-Ciocalteu assays (C,D). Results are showed for the hydro-alcoholic extraction (A,C) and the aqueous one (B,D) conducted on fruits of the three investigated invasive plants and the control species V. myrtillus. Green bars refer to the unripe fruits, whereas red bars indicate ripe ones. White dots express the mean values and the bars represent the 95% confidence interval. TE: Trolox Equivalents; GAE: Gallic Acid Equivalents.
The total phenol content assessed through Folin-Ciocalteu assay is reported in Fig. 3(C,D). Generally, unripe fruits are up to 2.3 times richer in polyphenols than the ripe ones (i.e. P. serotina aqueous extracts). Although L. japonica and P. americana extracts were not as rich in polyphenols as V. myrtillus, the hydro-alcoholic extraction enhanced their quantity, especially in the case of the latter species.
To evaluate the possible elements influencing antioxidant activity, we adopted a regression analysis to assess the effect of biological (ripening) and chemical (polyphenols and solvent of extraction) variables (Table 1). The occurrence of polyphenols is one of the most significant factors inducing antioxidant activity (p < 0.001) in all the investigated plants. In P. serotina and P. americana, unripeness plays a relevant role in the antioxidant activity (P. serotina p < 0.001, and P. americana p = 0.052). In the case of P. serotina, the antioxidant activity also depends on the interaction between the amount of polyphenols and the method of extraction (p < 0.001). This condition can be explained by taking into account the different extraction capacity and specificity of hydro-alcoholic solvent and water. For example, it was previously reported that 95% (v/v) ethanol is able to extract a larger amount and spectrum of phenolic compounds from red and white grape pomace with respect to water, and alcoholic extract possesses higher antioxidant activity39,40.
For the antioxidant properties of L. japonica, the interaction between the amount of polyphenols and the degree of ripening also appears to be significant, meaning that unripe fruit contain a higher amount of polyphenols than the ripe ones (p = 0.006).
Chemical characterization and identification of high-added values compounds
Given the well-recognized antioxidant activity of polyphenols, the occurrence of the most important classes was spectrophotometrically evaluated (Table 2). Results showed that aqueous extracts of unripe fruit from P. serotina had a higher amount of flavonoids compared to the ripe ones (up to 7.41 times) including catechins (not detected in ripe fruits), tannins (up to 5.5 times), and hydroxycinnamic acids (up to 4.1 times). A similar pattern was observed in hydro-alcoholic conditions. Noteworthy, the amount of hydroxycinnamic acids in P. serotina reached an average value of 650 mg of ferulic acid equivalents (FA eq.) per g of dry extracts in the aqueous extract from unripe fruits (more than two times the amount of FA eq. in V. myrtillus). Also, hydro-alcoholic extracts of unripe L. japonica fruits exhibited high amounts of hydroxycinnamic acids (386.68 mg of FA eq. per gram of dry extract). The highest amount of catechins was found in the aqueous extracts of P. serotina (average value of 73.69 mg of catechin equivalents per gram of dry extract), whereas no catechins were found in extracts from L. japonica and P. americana fruits.
All the unripe fruits processed with a hydro-alcoholic extraction showed similar or higher amounts of flavonoids than V. myrtillus. However, aqueous extracts of unripe P. serotina fruit showed flavonoid and tannin contents 2.8 and 2.4 times higher than V. myrtillus in the same conditions.
To better understand the biochemical profile of extracts obtained from the studied species, a total of 28 standard high-added value compounds were investigated by HPLC-DAD (Supplementary Table 1). Noteworthy, the most interesting compounds found in unripe fruit from P. serotina were chlorogenic acid (CLORA, avg. 3.137 mg*g−1) and rutin (RUT, avg. 1.856 mg*g−1 in hydro-alcoholic extract and 1.938 mg*g−1 in the aqueous one). In the same samples, catechins (e.g., epigallocatechin 3-O-gallate EGCG) were also detected (avg. 0.180 mg*g−1 in aqueous extract). A large amount of the unidentified chromatographic peaks showed an absorbance spectrum characteristic of catechins, thus confirming that these compounds are present in significant amounts. Finally, P. serotina unripe fruit showed some glycosylated stilbenes, particularly trans-piceid (tPIC avg. 0.596 mg * g−1).
RUT and some catechins were also detected in unripe fruits of P. americana, especially in the hydro-alcoholic extracts (RUT avg. 1.885 mg*g−1).
The most abundant metabolite found in L. japonica unripe fruits was caffeic acid (CAFA avg. 7.052 mg*g−1 in hydro-alcoholic extracts). In addition, some flavonoids such as RUT (avg. 3.350 mg*g−1) and quercetin (QUERC avg. 0.672 mg*g−1) were also detected in hydro-alcoholic extracts from unripe fruits.
Invasion risk and values of alien plant fruits
To better evaluate the risk of invasion in the three investigated plants, the viability of 100 seeds of each species was assessed (Supplementary Figure 1). Only seeds belonging to ripe fruits were analyzed, because unripe ones had no or contained only undeveloped seeds. The obtained results suggested that 54% of L. japonica, 56% of P. americana, and 72% of P. serotina were viable and therefore potentially active in enhancing the invasiveness of the sampled populations (Table 3). To estimate the chemical and biological value of these fruits/seeds for human aims, the yield of extraction for each species was also reported. Starting from 100 fruit we obtained a minimum of 0.749 g of dry extract (ripe fruits, hydro-alcoholic extraction) from L. japonica to a maximum of 7.069 g of dry extract (unripe fruits, aqueous extraction) from P. americana. Considering the best yield of extraction, the aqueous procedure obtained 7.043 mg of CLORA, 4.277 mg of RUT, and almost 0.400 mg of EGCG in P. serotina. Based on a rough estimate conducted by exploring the SciFinder database, these extracted chemicals could have a commercial value ranging from € 7.77 with the hydro-alcoholic procedure to € 58.97 with the aqueous one. In the case of P. americana, 100 unripe fruits extracted with the hydro-alcoholic solution produced about 8.010 mg of RUT and 2.418 mg of CAT with a total commercial value of € 10.89. Finally, L. japonica yielded 6.227 mg of CAFA from the hydro-alcoholic extraction of unripe fruits and almost 3 mg of RUT as well as 0.595 of QUERC in the same conditions with a total commercial value of € 13.45. In some cases, ripe fruit also showed a conspicuous economic value (e.g., aqueous extracts of P. americana and hydro-alcoholic extracts of P. serotina reached an estimated value of € 56.13 and of € 77.12 respectively). However, antioxidant activity detected by the DPPH assay was higher in unripe conditions than ripe ones.
Discussion
In this work we propose the exploitation of invasive plant species in a framework of bioprospecting41 to obtain high-added value compounds or phytocomplexes. Fruit collection for phytochemical extraction could also be seen as a way to prevent seed dispersal and therefore limit plant invasiveness.
Our results suggest that fruits from the investigated plants contain compounds with beneficial effects on human health that also have a high benchmark value on the market. This is the case of rutin, which is commonly known as a natural anti-inflammatory and antidiabetic compound42, stilbenes (trans-piceid and picetannol) known for their antimicrobial, anti-inflammatory, and antitumoral interest43, and hydroxycinnamic acids, such as chlorogenic and caffeic acids for which beneficial properties have been recognized as well44. Moreover, some of these metabolites show a high stability during gastric digestion, as is the case of piceid and rutin, which are glycolsylated and therefore protected during gastric digestion45. These two molecules are converted in the gut into the corresponding aglycones resveratrol and quercetin, which are polyphenols displaying a variety of biological activities in humans46,47 and are used as active components of several food supplements commercially available. This condition enhances the potential nutraceutical value of these extracts and supports their exploitation through bioprospecting.
We are aware that our proposal, if misunderstood, could open the possibility for the spread of invasive plants or even stimulate their deliberate cultivation. At the same time, we underline that the evolutionary history of plants shows that anthropogenic stress and overexploitation (including biopiracy) could result in a reliable containment or extinction of plant populations48. A typical example is Taxus contorta L., a Pleistocene tree on the brink of extinction in several Middle East regions, because bark and needles are harvested to produce anti-cancer medicines49. A more dramatic case is that of Coco de Mer (Lodoicea maldivica (J.F.Gmel.) Pers.) from the Seychelles, whose fruits are overexploited due to their supposed, but not scientifically confirmed, aphrodisiac properties50. Also plants having a wider geographical distribution, such as the European mountain species Arnica montana L.51 and Gentiana lutea L.52 have being interested by overexploitation and consequent extinction risk due to a growing market demand.
Given these considerations, we hypothesize that the harvest of invasive plants for bioprospecting purposes could help limit their uncontrolled spread in European and other countries opening new scenarios in the field of biodiversity conservation.
In Europe, the cost to manage invasive plants is estimated at 12 billion Euros per year53, in the USA, it is about 35 billion of dollars54, and in Australia, it is in about 4 billion dollars15. Despite these economic investments, the eradication effectiveness is limited to spot areas and the dissemination of invasive species mediated by natural ecosystem services is still advancing20,55. In the Lombardy Po plain, containment activities that have been conducted on the species investigated in this study are not effective. For example, in the case of P. serotina, an intensive management action was started in 1997 in the Ticino valley Park to try to limit the species. After 11 years, and despite costing 830000 Euros to contain it, this species enhanced its diffusion, overcoming the boundaries of the park56. Moreover, management activities produced indirect costs, such as those related to the waste disposal of the biomasses derived by pruning and extirpation actions. In this context, bioprospecting on plant biomass could help solve not only the problem of spreading invasive species, but also the management of waste. This agrees with current guidelines of agro-bioeconomy (see UE n.1141/2014), that stimulates the development of circular solutions that are able to transform waste into resources57.
An important element is the selection of the most suitable biomass. This study was focused on fruits, and our analysis suggested that most chemical compounds (e.g. polyphenols) and biological activities occur in unripe fruits rather than in ripe ones.
This is really relevant, because collecting fruit before they reach the ‘invasive developmental stage’ prevents seed dispersal. Moreover, we would like to stress that the Po plain is an important bird stopover area, and in a previous study, it was demonstrated that local and migratory birds are particularly attracted to dark fruits, such as L. japonica and P. americana20. Many migratory species prefer these fruits over those of local species, such as Viburnum spp. and Sambucus spp. Only after eradication actions of L. japonica and the massive restoration of local species did the diet of birds changed towards a greater exploitation of indigenous plant species20. Thus, at the local scale, bioprospecting activities on invasive species could enhance the success of ecological restoration plans.
From a technical point of view, the harvesting of unripe fruits could sound expensive and not sustainable. However, this activity could fit with a circular economy strategy for biodiversity management where ‘plant pickers’ are directly involved by forest and territory authorities to collect high-added value plant biomasses destined to processing by phytochemical companies. We underline that many bioactive metabolites and food ingredients are still today obtained from spontaneous or proto-domestic species and harvested by pickers58. Mediterranean diet is recently facing a comeback to the collection and use of spontaneous herbs such as the wild asparagus (Asparagus acutifolius L.) that is rich in antioxidants, wild cabbages (Brassica spp.) that contains high levels of folates and the maidenstears (Silene vulgaris (Moench) Garcke) that shows important amount of Omega-3 fatty acid59,60. Many companies involved in drugs and cosmetics production, regularly purchase great batches of wild plants to exploit their well-known bioactive components, including poisonous species as well. This is the case of Atropa belladonna L. that is already used to extract analgesic and antispasmodic phytocomplexes. Also fruit belonging to V. myrtillus, our experimental control species, are harvested at high altitudes on the Alps and Appennins61 (where they grow spontaneously), and other Vaccinium sp. are still harvested in nature in Canada and in the USA. The cultivated species V. corymbosum L. is polyploid, and although it has a higher production yield, is not as rich in antioxidants and anthocyanins as V. myrtillus62. The occurrence of secondary metabolites is also related to the response of the plant to different environmental conditions. Some studies highlighted how invasive plants show an altered metabolic profile in terms of secondary compounds (e.g. polyphenols) when they colonize new non-native territories63,64. An allelopathic response could explain this phenomenon: for example, caffeic acid extracted from L. japonica fruits is a typical allelopathic compound65. Moreover, it is known that also flowers and buds of this species are particularly rich in valuable compounds, such as chlorogenic acid and luteoloside that contribute to increase the economic value of L. japonica in terms of secondary metabolites66,67. Modifications in plant phytocomplexes during invasions also occur in P. serotina drupes. These fruits are appreciated by humans in native territories (USA and Mexico) where they are eaten fresh or utilized to prepare jams68, while in European countries they are poorly consumed due to bitter taste.
In this study, the chemical composition of the three alien species in their native areas was not assessed; however, a comparative analysis of secondary compounds from the original area vs. those in invaded areas could permit the identification of interesting metabolites for human aims and the evaluation of factors enhancing their presence. This could represent a core aspect of a bioprospecting approach along with the study of metabolic pathways and genes involved in the biosynthesis of high-added value compounds and the role played by the environment69.
In a conservation context, the characterization of allelopathic metabolites and of plant invasiveness must be performed before the import of an alien species in order to prevent its uncontrolled diffusion as has happened in the cases of L. japonica and P. serotina. The latter was introduced in Europe due to the high wood quality in its native territories; however, once established in Po plain, its xilema developed enormous tracheas making it rotten and of low interest for wood industries19. Therefore, this plague was introduced without any economic advantage.
In conclusion, a bioprospecting approach focused on alien species represents a promising strategy for discovering new metabolites, as well as to contain, or at least plan prevention strategies against plant invasiveness. This kind of approach could also support a sort of circular economy in the framework of invasive species management procedures, meaning that the metabolites found in the extracted phytocomplexes could produce economic gains that can be used to support, at least partially, the costs required for eradication actions.
Nowadays, there is an increasing trend to integrate ecosystem service topics within the management plans and strategies of protected areas. In this context, we encourage the adoption of bioprospecting focused especially on invasive species as an integrative element of these services to economically support (e.g. involving phytochemical industry) local biodiversity.
Change history
19 July 2018
A correction to this article has been published and is linked from the HTML and PDF versions of this paper. The error has been fixed in the paper.
References
Li, X., Liu, X., Kraus, F., Tingley, R. & Li, Y. Risk of biological invasions is concentrated in biodiversity hotspots. Front. Ecol. Environ. 14, 411–417 (2016).
Vilà, M. & Hulme, P. E. In Impact of Biological Invasions on Ecosystem Services (eds Vilà, M. & Hulme, P. E) Ch. 1, 1–14 (Springer International (2017).
Commission Implementing Regulation (EU) 2016/1141 of 13 July 2016 adopting a list of invasive alien species of Union concern pursuant to Regulation (EU) No 1143/2014 of the European Parliament and of the Council. Official Journal of the European Union L. 189, 4–8 (2016).
Hulme, P. E. et al. Grasping at the routes of biological invasions: a framework for integrating pathways into policy. J. Appl. Ecol. 45, 403–414 (2008).
Gioria, M. & Osborne, B. A. Resource competition in plant invasions: emerging patterns and research needs. Front. Plant Sci. 5, 501 (2014).
Stinson, K. et al. Invasive plants suppresses the growth of native tree seedlings by disrupting belowground mutualism. PLOS Biol. 4, e140 (2006).
Funk, J. L., Standish, R. J., Stock, W. D. & Valladares, F. Plant functional traits of dominant native and invasive species in mediterranean-climate ecosystems. Ecology 97, 75–83 (2016).
D’amato, G. et al. Allergenic pollen and pollen allergy in Europe. Allergy 62, 976–990 (2007).
Plank, L. et al. Benefits and costs of controlling three allergenic alien species under climate change and dispersal scenarios in Central Europe. Environ. Sci. Policy 56, 9–21 (2016).
Simberloff, D. How much information on population biology is needed to manage introduced species? Conserv. Biol. 17, 83–92 (2003).
Panetta, F. D. Evaluation of weed eradication programs: containment and extirpation. Divers. Distrib. 13, 33–41 (2007).
Panetta, F. D. Weed eradication - an economic perspective. Invasive Plant Sci. Manag. 2, 360–368 (2009).
Radosevich, S. R., Prather, T., Ghersa, C. M. & Lass, L. In Management of Invasive Weeds (ed Inderjit) Ch. 17, 345–359 (Springer, Dordrecht, the Netherlands, 2009).
Panetta, F. D. & Cacho, O. J. Beyond fecundity control: which weeds are most containable? J. Appl. Ecol. 49, 311–321 (2012).
Hoffmann, B. & Broadhurst, L. The economic cost of managing invasive species in Australia. NeoBiota 31, 1–18 (2016).
Carey, M. P., Sanderson, B. L., Barnas, K. A. & Olden, J. D. Native invaders–challenges for science, management, policy, and society. Front. Ecol. Environ. 10, 373–381 (2012).
Skirycz, A., Kierszniowska, S., Méret, M., Willmitzer, L. & Tzotzos, G. Medicinal bioprospecting of the Amazon rainforest: a modern Eldorado? Trends Biotechnol. 34, 781–790 (2016).
Bolpagni, R. & Piotti, A. Hydro-hygrophilous vegetation diversity distribution patterns in riverine wetlands agricultural landscapes: a case of study from the Oglio river (Po Plain, Northern Italy). Phytocoenologia 45, 69–94 (2014).
Banfi, E. & Galasso, G. La flora esotica lombarda (ed. Museo Di Storia Naturale Di Milano, 2010).
Galimberti, A. et al. Evaluating the efficacy of restoration plantings through DNA barcoding of frugivorous bird diets. Conserv. Biol. 30, 763–773 (2016).
Viana, D. S., Santamaría, L. & Figuerola, J. Migratory birds as global dispersal vectors. Trends Ecol. Evol. 31, 763–775 (2016).
Gray, E. R. & van Heezik, Y. Exotic trees can sustain native birds in urban woodlands. Urban Ecosyst. 19, 315–329 (2016).
McCusker, C. E., Ward, M. P. & Brawn, J. D. Seasonal responses of avian communities to invasive bush honeysuckles (Lonicera spp.). Biol. Invasions 12, 2459–2470 (2010).
Schierenbeck, K. A. Japanese Honeysuckle (Lonicera japonica) as an invasive species; history, ecology and context. CRC Crit. Rev. Plant. Sci. 23, 391–400 (2004).
McDonnell, M. J., Stiles, E. W., Cheplick, G. P. & Armesto, J. J. Bird-dispersal of Phytolacca americana L. and the influence of fruit removal on subsequent fruit development. Am. J. Bot. 71, 895–901 (1984).
Sebert-Cuvillier, E. et al. Local population dynamics of an invasive tree species with a complex life-history cycle: A stochastic matrix model. Ecol. Model. 201, 127–143 (2007).
Deckers, B. et al. Impact of avian frugivores on dispersal and recruitment of the invasive Prunus serotina in an agricultural land. Biol. Invasions 10, 717–727 (2008).
Fidaleo, M., Lavecchia, R. & Zuorro, A. Extraction of bioactive polyphenols with high antioxidant activity from bilberry (Vaccinium myrtillus L.) processing waste. Orient. J. Chem. 32, 759–767 (2016).
Amigoni, L. et al. Green coffee extract enhances oxidative stress resistance and delays aging in. Caenorhbditis elegans, J. Funct. Foods 33, 297–306 (2017).
Zhu, L., Liu, X., Tan, J. & Wang, B. Influence of harvest season on antioxidant activity and costituents of rabbiteye blueberry (Vaccinium ashei) leaves. J. Agric. Food Chem. 61, 11477–11483 (2013).
Blois, M. S. Antioxidant determinations by the use of a stable free radical. Nature 181, 1199–1200 (1958).
Ferri, M., Gianotti, A. & Tassoni, A. Optimisation of assay conditions for the determination of antioxidant capacity and polyphenols in cereal food components. J. Food Comp. Anal. 30, 94–101 (2013).
Singleton, V. L., Orthofer, R. & Lamuela-Raventos, R. M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 299, 152–178 (1999).
Zhishen, J., Mengcheng, T. & Janming, W. The determination of flavonoid in mulberry and their scavenging effects on superoxide radicals. Food Chem. 64, 555–559 (1999).
McMurrough, I. & McDowell, J. Chromatographic separation and automated analysis of flavanols. Anal. Biochem. 91, 92–100 (1978).
Stefan, M. B., Rodriguez, J. V., Blažeković, B., Kindl, M. & Vladimir-Knežević, S. Total Hydroxycinnamic Acids Assay: Prevalidation and Application on Lamiaceae Species. Food Anal. Methods 7, 326–336 (2014).
Porter, L. J., Hrstich, L. N. & Chan, B. G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 25, 223–230 (1986).
Ferri, M. et al. Chitosan treatment induces changes of protein expression profile and stilbene distribution in Vitis vinifera cell suspensions. Proteomics 9, 610–624 (2009).
Ferri, M. et al. Recovery of polyphenols from red grape pomace and assessment of their antioxidant and anti-cholesterol activities. N. Biotechnol. 33, 338–344 (2016).
Ferri, M. et al. White grape pomace extracts, obtained by a sequential enzymatic plus ethanol-based extraction, exert antioxidant, anti-tyrosinase and anti-inflammatory activities. N. Biotechnol. https://doi.org/10.1016/j.nbt.2017.07.002 (2017).
Reid, W. et al. Biodiversity Prospecting (World Resources Institute: Washington, 1993).
Gullón, B., Lù-Chau, T. A., Moreira, M. T., Lema, J. M. & Eibes, G. Rutin: a review on extraction, identification and purification methods, biological activities and approaches to enhance its bioavailability. Trends Food Sci. Technol. 67, 220–235 (2017).
Singh, C. K., Liu, X. & Ahmad, N. Resveratrol, in its natural combination in whole grape, for health promotion and disease management. Ann. N. Y. Acad. Sci. 1348, 150–160 (2015).
Taofiq, O., Gonzàlez-Paramàs, A., Berreiro, M. F. & Ferreira, I. Hydroxycinnamic Acids and Their Derivatives: Cosmeceutical Significance, Challenges and Future Perspectives, a Review. Molecules 22, e281 (2017).
Kim, H. et al. Metabolic and pharmacological properties of rutin, a dietary quercetin glycoside, for treatment of inflammatory bowel disease. Pharm. Res. 22, 1499–1508 (2005).
Gambini, J. et al. Properties of resveratrol: in vitro and in vivo studies about metabolism, bioavailability and biological effects in animal models and humans. Oxid. Med. Cell. Longev. 2015, Article ID837042 (2015).
David, A. V. A., Arulmoli, R. & Parasuraman, S. Overviews of biological importance of quercetin: a bioactive flavonoid. Pharmacogn. Rev. 10, 84–89 (2016).
Efferth, T. et al. Biopiracy of natural products and good bioprospecting practice. Phytomedicine 23, 166–173 (2016).
Poudel, R. C., Möller, M., Li, D. Z., Shah, A. & Gao, L. M. Genetic diversity, demographical history and conservation aspects of the endangered yew tree Taxus contorta (syn. Taxus fuana) in Pakistan. Tree Genet. Genomes 10, 653–665 (2014).
Castro, P., Azul, A. M. & Paiva, J. In Biodiversity and Education for Sustainable Development (eds Castro, P., Azeiteiro, U. M., Bacelar-Nicolau, P., Leal Filho, W. & Azul, A. M.) Ch. 2, 13–28 (Springer International, 2016).
Sugier D., Sugier, P. & Gawlik-Dziki, U. Propagation and introduction of Arnica montana L. into cultivation: a step to reduce the pressure on endangered and high-valued medicinal plant species. Sci. World J. 2013, Article ID 414363.
Rodanovic, D., Marković, T., Aiello, N. & Fusani, P. Cultivation trials on Gentiana lutea L. in Southern and South-easternEurope. J. Appl. Res. Med. Aromat. Plants 1, 113–122 (2014).
Kettunen, M. et al. Technical support to EU strategy on invasive alien species (IAS). London: Institut for European Environmental Policy (IEEP) (2009).
Pimentel, D., Ziniga, R. & Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 52, 273–288 (2005).
Schmiedel, D. et al. Evaluation system for management measures of invasive alien species. Biodivers. Conserv. 25, 357–374 (2016).
Caronni, F. E. Il caso del ciliegio tardivo (Prunus serotina Ehrh.) al Parco lombardo della Valle del Ticino. Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano XXXVI (2008).
Cristóbal, J., Limleamthong, P., Manfredi, S. & Guillén-Gosálbez, G. Methodology for combined use of data envelopment analysis and life cycle assessment applied to food waste management. J. Clean. Prod. 135, 158–168 (2016).
Bernardini, S., Tiezzi, A., Laghezza Masci, V. & Ovidi, E. Natural products for human health: an historical overview of the drug discovery approaches. Nat. Prod. Res. 1–25 doi:https://doi.org/10.1080/14786419.2017.1356838 (2017).
Conforti, F. et al. Bioactive phytonutrients (omega fatty acids, tocopherols, polyphenols), in vitro inhibition of nitric oxide production and free radical scavenging activity of non-cultivated Mediterranean vegetables. Food Chem. 129, 1413–1419 (2011).
Carvalho, A. M. & Barata, A. M. The Consumption of Wild Edible Plants. in Wild Plants, Mushrooms and Nuts: Functional Food Properties and Applications (eds I. C. F. R. Ferreira, P., Morales & L. Barros) Ch. 6, 159–198 (John Wiley & Sons, Ltd, United Kingdom (2017).
Zoratti, L., Jaakola, L., Haggman, H. & Giongo, L. Anthocyanin profile in berries of wild and cultivated Vaccinium myrtillus along altitudinal gradients in the Alps. J. Agric. Food Chem. 63, 8641–8650 (2015).
Giovanelli, G. & Buratti, S. Comparison of polyphenolic composition and antioxidant activity of wild Italian blueberries and some cultivated varieties. Food Chem. 112, 903–908 (2009).
Cappuccino, N. & Arnason, J. T. Novel chemistry of invasive exotic plants. Biol. Lett. 22, 189–193 (2006).
Kim, Y. O. & Lee, E. J. Comparison of phenolic compounds and the effects of invasive and native species in East Asia: Support for the novel weapons hypothesis. Ecol. Res. 26, 87–94 (2011).
Li, Z., Wang, Q., Ruan, X., Pan, C. & Jiang, D. Phenolics and plant allelopathy. Molecules 15, 8933–8952 (2010).
Kong, D. et al. A comparative study oft he dynamic accumulation of polyphenol components and the changes in their antioxidant activities in diploid and tetraploid Lonicera japonica. Plant Physiology and Biochemistry 112, 87–96 (2017).
Kong, D. et al. Correlation between the dynamic accumulation of the main effective components and their associated regulatory enzyme activities at different growth stages in Lonicera japonica Thunb. Industrial Crops and Products 96, 16–22 (2017).
Luna-Vázquez, F. J. et al. Nutraceutical value of black cherry Prunus serotina Ehrh. Fruits: Antioxidant and antihypertensive properties. Molecules 18, 14597–14612 (2013).
Baenas, N., García-Viguera, C. & Moreno, D. A. Elicitation: a tool for enriching the bioactive composition of foods. Molecules 19, 13541–13563 (2014).
Acknowledgements
We are grateful to G. Bazzi and G. Assandri for their invaluable assistance in graphics and data analysis and to R. Garavaglia, R. Gentili, and A. Losa for their help during sampling activities. This work was partially supported by Regione Lombardia in the framework of the Project “Food Social Sensor Network - Call Accordi per la Ricerca e l’Innovazione” co-funded by the Italian Government action “POR FESR 2014–2020”.
Author information
Authors and Affiliations
Contributions
L.G. collected samples, performed some of the experiments, analyzed the data and contributed to writing the manuscript; A.G. and M.L. coordinated the study and contributed to the writing of the manuscript; I.B. surveyed plant extraction procedures; C.M. performed antioxidant activity tests; M.F., A.T., E.S., and M.D.A. performed biochemical characterizations of plant extracts, and all authors contributed substantially to revisions.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Guzzetti, L., Galimberti, A., Bruni, I. et al. Bioprospecting on invasive plant species to prevent seed dispersal. Sci Rep 7, 13799 (2017). https://doi.org/10.1038/s41598-017-14183-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-14183-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
