
Abstract
Anaerobic ammonium oxidizing (anammox) bacteria own a central position in the global N-cycle, as they have the ability to oxidize NH4 + to N2 under anoxic conditions using NO2 −. They are responsible for up to 50% of all N2 released from marine ecosystems into the atmosphere and are thus indispensible for balancing the activity of N-fixing bacteria and completing the marine N-cycle. The contribution, diversity, and impact of anammox bacteria in freshwater ecosystems, however, is largely unknown, confounding assessments of their role in the global N-cycle. Here we report the activity and diversity of anammox bacteria in the world’s largest freshwater lake—Lake Superior. We found that anammox performed by previously undiscovered bacteria is an important contributor to sediment N2 production. We observed striking differences in the anammox bacterial populations found at different locations within Lake Superior and those described from other locations. Our data thus reveal that novel anammox bacteria underpin N-loss from Lake Superior, and if more broadly distributed across inland waters would play an important role in continental N-cycling and mitigation of fixed nitrogen transfer from land to the sea.
Similar content being viewed by others

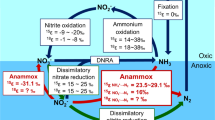

Introduction
As one of Earth’s great biogeochemical cycles, the N-cycle intimately couples to the carbon cycle, playing an important role in regulating global primary production1. The availability of N often limits productivity in many regions of the ocean, in most soils, and in some large freshwater environments2,3. Naturally, N is supplied to these environments through the microbial fixation of atmospheric N2 gas into biomass. In aerobic environments, this organic N is oxidized to nitrate, which can be mobilized from soils, and accumulate in groundwater, lakes, rivers, estuaries and the coastal ocean, leading to possible productivity increases and deleterious eutrophication with ensuing hypoxia or anoxia. Ultimate N removal from these environments back to the atmosphere depends on a combination of microbial denitrification and anammox. The anammox reaction is central to fixed nitrogen removal from the oceans where it operates in oxygen minimum zones and sediments playing an outsized role in closing the N-cycle4. Its role in the continental N-cycle and its possible activity in freshwater surface environments remains largely unknown.
Anammox bacteria have been detected in freshwater sediments5,6 and appear diverse and widely distributed in soils7,8,9,10. Potential anammox activity, on the other hand, has only been demonstrated in the water column of a single lake, Lake Tanganyika, a stratified, ancient lake situated in East Africa11 as well as in eutrophic lake sediments from Japan6 and China8. As a broader and quantitative test for the distribution and activity of anammox, we have thus combined molecular microbial techniques with geochemical analyses and process rate measurements to chart the diversity and biogeochemical significance of anammox bacteria in the sediments of Lake Superior—the world’s largest lake.
Holding 10% of the world’s surface freshwater, Lake Superior is an important natural resource. It is characterized by low rates of primary production12, and its sediments exhibit low rates of biogeochemical activity and very deep oxygen penetration (Fig. 1)13. Nitrate concentrations in Lake Superior are high (>20 µmol l−1) and have been increasing over the last century14. The high nitrate and low phosphate concentrations lead to extreme nutrient stoichiometry and N:P ratios of >10 00014. Nitrification in Lake Superior’s sediments15,16 leads to accumulation of nitrate (and nitrite) in the upper reaches of the sediment and penetration16 of nitrate well into the anoxic deeper sediments where ammonium accumulates (Fig. 1). The juxtaposition of oxidized forms of nitrogen with ammonium leads to an environment favorable to anammox. This pattern holds across multiple locations across the lake, and the nearly constant conditions at these great depths ensure minimal seasonal variability16.

Profiles illustrating the depth distributions of oxygen, NOx (combined nitrate and nitrite), and ammonium at stations IR (a) and SW (b).
We used whole core incubations to measure N2 production rates and pathways in Lake Superior’s sediments at two deep-water stations (IR and SW)17. The two stations exhibit similar biogeochemical properties, but the deeper sediments at station SW appear to be less active leading to deeper penetration of oxidized nitrogen, and lower ammonium concentrations (Fig. 1). Low total rates of N2 production were found at both stations (Table 1) and are consistent with the low overall biogeochemical activity of Lake Superior’s sediments, as indicated by oxygen uptake rates13. Higher overall rates of N2 production were observed at station IR than at SW, in line with shallower penetration of nitrate (and nitrite) at IR than SW. The anammox reaction was operative at both stations, and indeed dominated N2 production at SW, contributing more than 50% of the total N2 produced. At station IR, anammox was responsible for 24% of the total N2 production. Greater importance of anammox to N2 production at station SW compared to IR is in line with the general overall observation that anammox plays a proportionally greater role relative to denitrification in sediments with lower overall biogeochemical activity18,19,20, as for example qualified by oxygen respiration rates. Likewise, absolute rates of anammox at station IR are much higher than at SW despite the fact that its relative contribution to N2 production is lower. Similar observations have been made in marine sediments18, implying comparable controls on N loss processes in both marine and freshwater environments.
The difference in the rates and relative importance of anammox observed between sites was also reflected in the diversity of the hydrazine synthase gene (hzsA) (Fig. 2), a functional marker for anammox bacteria21. From all cultivated anammox bacteria, the hzsA from ‘Candidatus Brocadia fulgida’ was the closest relative to the hzsA sequences we recovered from both stations, which is consistent with the known affinity of Ca. Brocadia for non-saline environments. The hzsA sequences from the two Lake Superior stations clustered separate from each other, with those from IR forming a novel monophyletic group exclusively containing sequences from this station. The sequences obtained from SW also cluster together, but two sequences from the Ooijpolder eutrophic freshwater ditch sediment21 were closely related, indicating a likely wider distribution of these anammox bacteria in freshwater environments. This implies that the diversity of anammox bacteria is underexplored and may be much broader than currently known when freshwater taxa are fully described.

Neighbor joining phylogenetic reconstruction of HzsA sequences from Lake Superior based on 397 positions of an amino acid alignment. Bootstrap values (1000 replicates) above 50% are shown. Candidatus Scalindua sp. enrichment culture clone 15L (AEW50032) was used as outgroup.
Dissimilatory reduction of nitrate to ammonium (DNRA) can lead to the underestimation of anammox due to the conversion of some 15N-NO3 − to 15N-NH4 +, and its incorporation into the30N2 (15N15N) pool. We assayed for DNRA in our whole-core incubations, but did not detect it. Our measurements, however, may be blind to DNRA tightly coupled to anammox, such as that observed in sediments underlying marine oxygen minimum zones22. We thus take our whole-core measurements as minimum estimates for the contribution of anammox to total N2 production.
To verify the anammox activity observed in our whole-core incubations, we measured potential rates of denitrification and anammox in anoxic slurry incubations. In anoxic slurry incubations with added 15N-NO3 −, no 14N-NO3 − and in the absence of DNRA, anammox produces only 29N2 (14N15N), whereas the only known 30N2 production pathway is through denitrification. As with our whole core measurements, DNRA was not detected in any of our slurry incubations. Both 29N2 and 30N2 were produced in the slurry incubations confirming the activity of anammox in sediments from both stations. As in the whole core incubations, anammox contributed a large fraction (Table 2), up to 57%, to the total N2 produced. At IR, the highest rates of N2 production, and the highest contribution of anammox were both observed in the upper sediment layer, coinciding with the highest in situ nitrate concentrations of all layers. In contrast, the highest rates of N2 production, and the highest contribution of anammox were observed in the deeper sediment layer at station SW. This is likely related to a deeper in situ nitrate maximum at SW than at IR, which itself is the likely result of lower rates of oxygen consumption and deeper oxygen penetration at SW.
Anoxic slurry incubations were also conducted with 15N-NH4 + with addition of allylthiourea (ATU) to specifically inhibit of aerobic ammonium oxidation. Only anammox is known to produce 15N-labeled N2 (29N2) in incubations with 15N-NH4 +. Notably, without added nitrate, anammox in these experiments will be nitrite limited, owing to low rates of nitrite production from the low natural nitrate concentrations present. Again, anammox was observed at both stations, however, it was only observed at the shallowest depth at IR, and then at lower rates than observed in the incubations with added 15N-NO3 −. We attribute this to nitrite limitation in the shallow sediment and a complete lack of nitrate and therefore nitrite in the deeper sediment layer at IR. Both SW sediment layers yielded anammox from 15N-NH4 +. Natural nitrate concentrations are higher at SW, and indeed, 15N-NH4 + based rates are comparable to 15N-NO3 − based rates for the shallower depth. In contrast, 15N-NH4 + based rates are lower than 15N-NO3 − in the deeper layer, and again, we attribute this to nitrite limitation.
Our experiments also provide information on the anaerobic oxidation of NH4 + to N2 with alternative oxidants, which has been previously proposed23, but remains unsubstantiated by a robust set of nitrogen isotope labeling experiments. In our 15N-NH4 + based experiments, the direct oxidation of NH4 + to N2 would be revealed as 30N2 production, which was absent in all incubations except in the deeper sediment layer from station SW. Here 30N2 production rates are low, but significant, suggesting low rates of oxidation of NH4 + to N2 under anoxic conditions or nitrification by ATU-insensitive ammonium oxidizing archea. Possible oxidants include the (hydr)oxides of Fe and Mn, which are abundant in station SW sediments13. Further work is required to verify this process and the oxidant involved, but in Lake Superior, the rates appear to be very low and insignificant to N-cycling.
To gain a quantitative appreciation for how anammox contributes to Lake Superior’s N-cycle, we have calculated as mass-balance on N for both stations (Fig. 3). N is largely supplied to the sediment through organic matter deposition. Only a small fraction of this N is ultimately buried, however, and much of the N deposited is returned to the water column either as NO3 − or as N2. Mass balance calculations yield N2 production rates that are very similar to those measured in our whole-core incubations giving us confidence that we have accurately captured the in situ rates and pathways. Anammox therefore plays an important role in overall N2 production, quantitatively influencing the amount of fixed N buried, the amount recycled to the water column, and thus overall N cycling in Lake Superior.

Mass balance for nitrogen in stations IR (a) and SW (b). Shaded area represents anoxic sediments. Descriptions of calculations are included in the materials and methods (mmol m−2 d−1).
Oxygen and carbon cycling in Lake Superior compares well with trends observed in coastal marine environments13. The relative contribution of anammox to total N2 production also agrees with relative contributions determined in coastal marine sediments with similar water depths, maximum nitrate concentrations, and oxygen penetration depths18,20. Rates of anammox measured in Lake Superior, however, are lower than those typically measured in coastal marine sediments18. We hypothesize that this is related to the low rates of ammonium supplied through carbon degradation in Lake Superior sediments13.
Our experiments document appreciable anammox activity in Lake Superior’s sediments, showing that anammox can play an important role in the sedimentary N-cycle of freshwater environments. Our data also indicates that there are differences in the diversity of anammox bacteria between sites in Lake Superior, with a novel group of anammox bacteria likely present at the IR station. We find that the importance of anammox to total N2 production scales inversely with overall sediment biogeochemical activity in a fashion similar to that observed in marine sediments. We predict then, that anammox is most important in low productivity freshwater environments such as high-latitude lakes. As such, anammox would have global importance to continental N-cycling, and may be a key regulator of the productivity of inland ecosystems. This possibility should be tested through further studies to expand the geographical range of our observations and to more completely map the diversity of anammox bacteria on the continents.
Material and Methods
Sample collection
Sediment samples were collected from stations SW and IR using the R/V Blue Heron. Sediment cores of 94-mm inner diameter were recovered using an Ocean Instruments multi-corer. The landing sites were monitored using a Knudsen 320/R echo sounder with a 28-kHz transducer to select flat areas with laterally homogeneous sediment accumulation.
Process rate measurements
Whole-core measurements of N-transformation rates were measured as previously described17 and only key differences in methodology are noted here. 15N-NO3 − was added to both the overlying water and injected into sideports in core tubes at 1 cm intervals to depths of 7 cm at SW and 5 cm at IR and to nominal final concentrations of 25 µM 15N-NO3 −. The ratio of 15N/14N in the nitrate pool within the zone of nitrate reduction was estimated by measuring the isotopic composition of N2O, as previously described17. A total of 5 undisturbed intact sediment cores were incubated from each site and incubations terminated after 6, 16, and 38 hours. No relationship to incubation time was observed. The cores were incubated at 4 °C, which corresponds to the temperature (3–5 °C) of Lake Superior bottom waters from May to November. The cores were maintained at 4 °C in the dark and the overlying water was stirred at 60 revolutions min−1 using a magnetic stir bar suspended 3–4 cm above the sediment–water interface. Core tubes were sealed with thick rubber stoppers. Oxygen concentrations decreased over the course of the incubations, but not more than 10% from their in situ values and the measured decrease was linear throughout the incubation period.
Sediment slurries were prepared by mixing sediment from the top 0–3 and 3–6 cm of the core with an equivalent volume of bottom water that was previously purged with ultra-high-purity He gas to remove O2 and N2. The sediment slurry was subsequently incubated under He for an additional 24 h to remove residual N2 gas. Following this 24-h period, the sediment slurry was transferred, with no headspace, into ~12-ml gas tight vials (Exetainers, LabCo). Isotopic labels, and specific inhibitor (ATU, final concentration of 165 µmol L−1) were added. The sediment slurries were incubated at room temperature ~20 °C and mixed ~every 6 hours by inversion 3x. Upon sacrificing, 1 ml of slurry was removed from the Exetainer using a needle and syringe and replaced with He gas and 200 μl of a 37% formaldehyde solution to stop microbial activity. The withdrawn sediment slurry (1 ml) was filtered directly through a 0.2 μm pore size syringe filter and the filtrate was frozen for later analysis of NOx and NH4 + concentrations. The formaldehyde-fixed sediment slurry was stored upside down in the Exetainers until isotopic analysis.
The isotopic composition of N2 was determined by injecting 25–50 μl of headspace gas into an in-house built injection system. Following injection, CO2 was trapped using Ascarite, N2 and N2O separated using a Poropak R GC column, and the sample stream passed through a reduction reactor to reduce N2O to N2 and O2 to H2O. H2O was trapped on Mg perchlorate, and the sample stream was introduced using a Conflo III to a Thermo Electron DELTA V plus IR-MS operated in continuous-flow mode. N2 was measured at masses 28, 29, and 30. Similarly, the N isotopic composition of N2O was measured by injecting 200–1000 μl of headspace gas, but the reduction reactor was bypassed and isotopic measurements were made on masses 44, 45, and 46. Measurements of 15N-NH4 + were conducted by converting NH4 + to N2 following oxidation by hypobromite. In the case of the slurry incubations, NH4 + was extracted in a 2 mol l−1 KCl solution prior to hypobromite oxidation and isotopic analysis.
Mass balance calculations
The N sedimentation flux at the sediment water interface (SWI) and in the deep sediments (burial) was calculated from carbon flux (12C:1N for Lake Superior sediments)13. The C burial flux was calculated from organic carbon concentrations and sedimentation rates measured by 210Pb. The carbon sedimentation flux (at the SWI) was carbon burial plus carbon remineralization. The latter was calculated from total oxygen uptake minus the oxygen consumed by nitrification. Fluxes of nitrate and ammonium are diffusive fluxes calculated using Fick’s Law, from the porewater concentration gradients. Nitrification rates were calculated by integrating nitrification rates in the surface sediments. Rates are second derivatives of the NOx profile, and the integration is essentially the difference between nitrate flux at SWI and at the depth where nitrification goes to zero, i.e. the depth of oxygen penetration. This calculation though, is based on the assumption that zones of nitrification and nitrate reduction do not overlap16. Denitrification/Anammox rates were calculated by a mass balance approach. R(N2 removal) = Ammonium production by C remineralization − Flux out of ammonium + Flux out of nitrate. Ammonium production can be calculated from the rates of carbon mineralization and a 12C:1N ratio.
Molecular microbiology
Metagenomic DNA was extracted using the PowerLyzer™ PowerSoil® DNA Isolation Kit from MoBio following the manufacturers instructions. DNA was quantified spectrophotometrically (NanoDrop) and furthermore reviewed on a 1% agarose gel.
In order to analyze the hzsA genes present in our samples we conducted a nested PCR and prepared a clone library with the resulting products. In the first PCR round the hzsA gene was amplified with hzsA_526F (5′-TAYTTTGAAGGDGACTGG-3′) as forward and hzsA_1857R (5′-AAABGGYGAATCATARTGGC-3′) as reverse primer. The second round was performed with with hzsA_526F as forward and with hzsA_1829R (5′-TCATACCACCARTTGTA-3′) as reverse primer (for primer see Harhangi 2012). PCR reactions were performed in 50 µl end volume containing 5–10 ng template DNA respectively 1 µl of PCR product, 20 pmol of each primer, 1.25 Units Taq polymerase (Thermo), 40 nmol deoxynucleotides and 125 nmol MgCl2 in 1x Taq reaction buffer with KCl (Thermo). PCR reactions were performed as follows: 5 min at 96 °C followed by 35 resp. 32 cycles of 96 °C for 1 min, 53 °C resp. 51 °C for 1 min and 72 °C for 1.5 min. Final extension was performed at 72 °C for 5 min. Obtained PCR products were purified using the QIAEX® II Gel Extraction Kit from Qiagen and cloned into the pJET1.2/blunt cloning vector using the CloneJET PCR Cloning Kit from Thermo following the instructions of the manufacturers. Resulting clones were sequenced by Macrogen Europe (Netherlands) using the primers pJET1.2F (5′-CGA CTCAC TATAGGGAGA GCGGC-3′) and pJET2.1R (5′-AAGAACATCGATTTTCCATGG CAG-3′). In total 16 clones from each station were sequenced. In total 16 clones from each station were sequenced, of which 29 were hzsA genes without ambiguous bases. The sequences have been deposited in Genbank with the accession numbers MF565415 to MF565443.
References
Gruber, N. & Galloway, J. N. An Earth-system perspective of the global nitrogen cycle. Nature 451, 293–296, https://doi.org/10.1038/nature06592 (2008).
Vitousek, P. M. & Howarth, R. W. Nitrogen Limitation on Land and in the Sea - How Can It Occur. Biogeochemistry 13, 87–115 (1991).
Elser, J. J. et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10, 1135–1142 (2007).
Thamdrup, B. In Annual Review of Ecology, Evolution, and Systematics, Vol 43 Vol. 43 Annual Review of Ecology Evolution and Systematics (ed D. J. Futuyma) 407–428 (2012).
Penton, C. R., Devol, A. H. & Tiedje, J. M. Molecular evidence for the broad distribution of anaerobic ammonium-oxidizing bacteria in freshwater and marine sediments. Appl Environ Microb 72, 6829–6832 (2006).
Yoshinaga, I. et al. Distribution and Diversity of Anaerobic Ammonium Oxidation (Anammox) Bacteria in the Sediment of a Eutrophic Freshwater Lake, Lake Kitaura, Japan. Microbes Environ 26, 189–197 (2011).
Humbert, S. et al. Molecular detection of anammox bacteria in terrestrial ecosystems: distribution and diversity. Isme J 4, 450–454 (2010).
Zhu, G. B. et al. Hotspots of anaerobic ammonium oxidation at land-freshwater interfaces. Nat Geosci 6, 103–107 (2013).
Zhu, G. B. et al. Anaerobic ammonia oxidation in a fertilized paddy soil. Isme J 5, 1905–1912 (2011).
Hirsch, M. D., Long, Z. T. & Song, B. Anammox Bacterial Diversity in Various Aquatic Ecosystems Based on the Detection of Hydrazine Oxidase Genes (hzoA/hzoB). Microb Ecol 61, 264–276 (2011).
Schubert, C. J. et al. Anaerobic ammonium oxidation in a tropical freshwater system (Lake Tanganyika). Environ Microbiol 8, 1857–1863 (2006).
Sterner, R. W. In situ-measured primary production in Lake Superior. J Great Lakes Res 36, 139–149 (2010).
Li, J. Y. et al. Carbon mineralization and oxygen dynamics in sediments with deep oxygen penetration, Lake Superior. Limnol Oceanogr 57, 1634–1650 (2012).
Sterner, R. W. et al. Increasing stoichiometric imbalance in North America’s largest lake: Nitrification in Lake Superior. Geophys Res Lett 34 (2007).
Small, G. E. et al. Rates and controls of nitrification in a large oligotrophic lake. Limnol Oceanogr 58, 276–286, https://doi.org/10.4319/lo.2013.58.1.0276 (2013).
Li, J. & Katsev, S. Nitrogen cycling in deeply oxygenated sediments: Results in Lake Superior and implications for marine sediments. Limnol Oceanogr 59, 465–481, https://doi.org/10.4319/lo.2014.59.2.0465 (2014).
Trimmer, M. & Nicholls, J. C. Production of nitrogen gas via anammox and denitrification in intact sediment cores along a continental shelf to slope transect in the North Atlantic. Limnol Oceanogr 54, 577–589 (2009).
Dalsgaard, T., Thamdrup, B. & Canfield, D. E. Anaerobic ammonium oxidation (anammox) in the marine environment. Res Microbiol 156, 457–464 (2005).
Dalsgaard, T. & Thamdrup, B. Factors controlling anaerobic ammonium oxidation with nitrite in marine sediments. Appl Environ Microb 68, 3802–3808 (2002).
Trimmer, M. & Engström, P. In Nitrification (American Society of Microbiology, 2011).
Harhangi, H. R. et al. Hydrazine Synthase, a Unique Phylomarker with Which To Study the Presence and Biodiversity of Anammox Bacteria. Appl Environ Microb 78, 752–758 (2012).
Prokopenko, M. G. et al. Nitrogen losses in anoxic marine sediments driven by Thioploca–anammox bacterial consortia. Nature 500, 194–198 (2013).
Luther, G. W., Sundby, B., Lewis, B. L., Brendel, P. J. & Silverberg, N. Interactions of manganese with the nitrogen cycle: Alternative pathways to dinitrogen. Geochimica Et Cosmochimica Acta 61, 4043–4052, https://doi.org/10.1016/s0016-7037(97)00239-1 (1997).
Acknowledgements
We thank captain Mike King and the crew of R/V Blue Heron, as well as marine technician Jason Agnich. This work was supported through NSF OCE 0961720 grant to SK, Danish grant no. DNRF53 to DEC, and NSERC Discovery grant 0487 to SAC.
Author information
Authors and Affiliations
Contributions
S.A.C., A.H.T., and S.K. designed research. S.A.C., C.M., M.F., J.L. and A.H.T. conducted experiments and analyses. S.A.C. wrote the paper with input from A.H.T., M.F., B.T., S.K. and D.E.C.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Crowe, S.A., Treusch, A.H., Forth, M. et al. Novel anammox bacteria and nitrogen loss from Lake Superior. Sci Rep 7, 13757 (2017). https://doi.org/10.1038/s41598-017-12270-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-12270-1
This article is cited by
-
Structure and Functional Properties of Bacterial Communities in Surface Sediments of the Recently Declared Nutrient-Saturated Lake Villarrica in Southern Chile
Microbial Ecology (2023)
-
Nitrogen removal processes in lakes of different trophic states from on-site measurements and historic data
Aquatic Sciences (2021)
-
Extracellular electron transfer-dependent anaerobic oxidation of ammonium by anammox bacteria
Nature Communications (2020)
-
Removal Capacities and Environmental Constrains of Denitrification and Anammox Processes in Eutrophic Riverine Sediments
Water, Air, & Soil Pollution (2020)
-
Nitrous oxide in the Great Lakes: insights from two trophic extremes
Biogeochemistry (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
