
Abstract
Of the nearly 6,800 extant frog species, most have weak jaws that play only a minor role in prey capture. South American horned frogs (Ceratophrys) are a notable exception. Aggressive and able to consume vertebrates their own size, these “hopping heads” use a vice-like grip of their jaws to restrain and immobilize prey. Using a longitudinal experimental design, we quantified the ontogenetic profile of bite-force performance in post-metamorphic Ceratophrys cranwelli. Regression slopes indicate positive allometric scaling of bite force with reference to head and body size, results that concur with scaling patterns across a diversity of taxa, including fish and amniotes (lizards, tuatara, turtles, crocodylians, rodents). Our recovered scaling relationship suggests that exceptionally large individuals of a congener (C. aurita) and extinct giant frogs (Beelzebufo ampinga, Late Cretaceous of Madagascar) probably could bite with forces of 500 to 2200 N, comparable to medium to large-sized mammalian carnivores.
Similar content being viewed by others

Introduction
The evolution of jaws and their use in prey capture has played a key role in the radiation and evolutionary success of vertebrates1,2,3,4,5. However, in most non-larval frogs and salamanders, derived tongue projection mechanisms serve as the primary means of capturing prey, and the jaws tend to be weak and play an ancillary role in predation6,7. Horned frogs (Ceratophrys) are a notable exception.
Although Ceratophrys frogs possess large and highly adhesive tongues8, they also have strong jaws and, quite atypical of frogs, readily bite. The strength of their bite is reflected by their extremely wide and short heads, which contain large jaw-adductor muscles and provide a high mechanical advantage, even at the tips of the jaws. Using their disproportionately large head and the forceful bite it affords, Ceratophrys frogs are capable of capturing and subduing prey that can be large, strong, and/or potentially dangerous (e.g., frogs, lizards, snakes, birds, rodents; ref.9).
In vivo bite-force performance has been studied in a diversity of vertebrates10. Interspecific and intraspecific studies of bite force, including some analyses of ontogenetic variation, have been conducted in various amniotes and some fish, with the results consistently indicating a pattern of positive allometric scaling of bite force with respect to body size and cranial dimensions11,12,13,14,15,16,17,18,19,20,21,22,23,24 (Table 1). An understanding of the ontogenetic scaling of biomechanical performance provides a rare opportunity to gain insight into the ecology and behavior of extinct taxa, including those beyond the size range of extant analogues22,24.
Here, we present an analysis of the ontogenetic scaling of in vivo bite force in Ceratophrys cranwelli 25 (Cranwell’s Horned Frog26). Using a longitudinal experimental design, we measured bite-force performance in a sample of eight post-metamorphic individuals to test the hypothesis that, typical of fish and amniotes examined to date, this non-amniote tetrapod exhibits a positive allometric increase in bite force during ontogeny. We then use our data to extrapolate the maximum bite-force performance of two other taxa, an exceptionally large museum specimen of an extant congener (C. aurita), as well as a giant ceratophryid-like frog from the Late Cretaceous of Madagascar (Beelzebufo 27,28) that lived contemporaneously with small crocodylians and non-avian dinosaurs that were potential prey. In light of our results, we discuss the role of a forceful bite and associated specializations in shaping the unusual dietary behavioral ecology of these megalophagous predators.
Results
Scaling of bite-force performance in Ceratophrys cranwelli
Our sample of eight frogs spanned a nearly 17-fold range in body mass (8.9–147.8 g) and a 1.9 to 2.4-fold range in linear morphometrics of the body (length: 39.8–95.6 mm) and head (length: 14.7–31.7 mm; width: 22.4–46.1 mm; depth: 10.8–20.6 mm; Supplementary Table S1). Direct measurement of bite force (Fig. 1) produced voluntary performance that varied 12-fold (2.7–32.9 N).

An individual Ceratophrys cranwelli biting a force transducer. Leather strips glued to ends of bite bars provide a natural surface that encourages high-effort biting and avoids damage to teeth and bones. The strips also indicate a bite point for standardization of bite-force performance (see Methods).
Scaling coefficients of bite force on the linear morphometric variables (log-transformed) ranged from 2.68–4.07, in all cases significantly exceeding the predicted value of 2.0 based on isometric scaling (Table 2). Similarly, the logarithmic scaling coefficient of bite force on body mass (0.91) exceeded that predicted by isometry (i.e., 0.\(\mathop{6}\limits^{\bar{} }\)). The relationship between bite force and body size (body mass and length), as well as between bite force and head width, did not differ among the individual frogs (ANCOVA with non-significant individual by covariate interactions, Table 3, Fig. 2).

Head width vs. bite force for specimens of Ceratophrys cranwelli. (a) Plot of bite force on head width in which each point represents maximum bite force of three trials (standardized here for bite position at posterior teeth, see Methods) measured during an experimental session with each specimen represented by a different symbol (n = 8). The line represents a power function fitted to the pooled data (y = 0.0004x2.9608, R2 = 0.9209). (b) Log-log plot in which each point represents maximum bite force (standardized here for bite position at jaw tips, see Methods) measured during an experimental session with each specimen represented by a different symbol (n = 8). Thin linear regression lines are given for each specimen. The thick dotted regression line was calculated from the pooled data (slope = 3.3269). Estimated bite forces for Beelzebufo were calculated using reduced major axis regression (see Methods).
Full model and stepwise multiple regressions with bite force as the dependent variable and all of the morphometrics as the independent variables indicate that head width is by far the best predictor of bite force in our sample (full model — F5, 57 = 135.56, p < 0.0001, head width: p = 0.0005, all other variables: p ≥ 0.232; forward and backward stepwise models retain only head width and mass — F2, 57 = 348.19, p < 0.0001, head width: p < 0.0001, mass: p = 0.0394; partial correlations — head width = 0.457, all other variables ≤ 0.165).
Estimates of jaw-adductor muscle size in Ceratophrys frogs and Beelzebufo
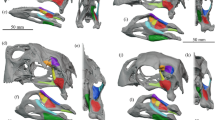
The ventral aspect of the adductor chambers of frogs is bound by bony elements that form a conduit for the primary jaw-adductor musculature. Because the muscle fibers are near vertically oriented as they pass through the conduit, the area of the pair of conduits in palatal view serves as a gauge of the cross-sectional area of the primary jaw muscles that generate bite force. We found that the average area of the pair of palatal conduits, relative to head width (the external morphometric that best predicts bite force in live specimens), is comparable among C. cranwelli, other Ceratophrys spp. including the exceptional C. aurita specimen, and Beelzebufo (Fig. 3; Supplementary Table S2).

Relationship between head width and size of palatal conduits of major jaw-adductor musculature of ceratophryid frogs. Log-log plot of head width vs. average area of right and left palatal conduits of the major jaw-adductor musculature of Ceratophrys spp. based on dissections and skeletal specimens. Solid linear regression line represents all Ceratophrys spp. (open squares □, y = 2.4902x − 2.4094, R2 = 0.9524). Dotted linear regression line represents only C. cranwelli (filled squares ■, y = 2.0263x − 1.6627, R2 = 0.8784). Estimate for the exceptional C. aurita specimen (LACM 163430) is represented by a triangle (△). Three estimates for the Beelzebufo composite reconstruction with a head width of 111 mm (Fig. 3D in ref.28) are represented by diamonds (◊), based on conservative, likely, and maximum conduit size. Silhouettes scaled to the same head width, from smallest to largest, are provided for C. cranwelli (collection of A.K.L.), C. ornata (UMCZ R1529), C. ornata (LDUCZ W186), C. aurita, and the Beelzebufo reconstruction. Scale bars = 40 mm. Note that our estimate of maximum bite force for Beelzebufo is based on larger individuals with a head width of 154 mm (estimate of maximum size provided in ref.28 based on recovered material).
Predictions for other frogs
A pooled RMA regression of log bite force on log head width produced a positively allometric linear scaling coefficient of 3.3269 ± 0.1524. The intercept calculated for a bite position at the jaw tips was −4.2336. At the jaw antero-posterior midpoint (i.e., midpoint between jaw tips and jaw joint measured in parallel to the antero-posterior axis of the skull), approximately the position of the posterior teeth, the intercept was −3.9326. Using regression means for log head width and log bite force at each jaw position, the linear relationship described by the model (log bite force = 3.3269 *log head width – intercept) predicts that Ceratophrys aurita specimen LACM 163430 (head width = 98.3 mm) would have had a bite force of 248.6 N at the jaw tips and 497.1 N at the jaw midpoint (Table 4). The same model predicts that a Beelzebufo with head width of 111 mm (FMNH PR 2512) would have bite forces at the jaw tips/midpoint of 372.4/744.8 N, and large individuals (head width = 154 mm, UA 9269; ref.28) would have bite forces of 1106.8/2213.7 N.
Discussion
Our results demonstrate that extant Ceratophrys frogs bite with forces comparable to those of many predatory amniotes (~5 to 500 N at the jaw midpoint). This capacity for forceful biting is associated with prey capture using a large and highly adhesive tongue8. Nonetheless, following tongue adhesion large prey must be trapped and restrained with a vice-like clamp of the jaws. In contrast to the gracile skull and flexible mandible typical of most frogs29, the skull of ceratophryids is rigid and heavily built, the mandibular symphysis is fused, and the mentomeckelian joints are immovable30,31. These characteristics should more effectively transmit jaw-adductor muscle forces to the mandible during a bite, including an enhanced transmission of muscle forces from the balancing to the working side of the jaws during a unilateral bite. Furthermore, extant ceratophryids have extremely short jaws (i.e., short jaw out-lever) thus providing great mechanical advantage for biting. Greater bite forces permit access to a broader range of prey in predators that use their jaws to subdue prey32,33.
Complementing the musculoskeletal robustness of the jaws, the unicuspid and non-pedicellate teeth of Ceratophrys 31,34,35 are unusual for frogs, and exhibit structural characteristics that reflect their function during prey dispatch (Fig. 4). Having sharp tips and being strongly attached to the premaxillae and maxillae27,31, they are well-suited to produce high pressures to effectively penetrate and engage the integument of prey. They also are recurved, labiolingually expanded, and have robust bases, and thus are ideally shaped to resist forces produced by struggling prey and prevent it from escaping. In addition, Ceratophrys has a pair of large and fully ossified odontoids (fangs) on either side of the mandibular symphysis, which enhance the ability to secure large, strong prey28,31. In contrast to theoretical predictions, we found that in C. cranwelli bite-force performance scales with strong positive allometry with respect to both body size and head dimensions. Given that the physiological cross-sectional area of a muscle is a major determinant of its capacity to generate force36, and assuming isometric growth and maximum voluntary performance throughout ontogeny, it follows that bite force should scale isometrically with the sum of the cross-sectional areas of all of the jaw-adductor muscles contributing to a bite. Because areas scale to the square of linear measurements, bite force also is predicted to scale to the square of linear measurements (slope = 2.0)14. Moreover, as areas scale to the two-thirds power of volumes, and volume scales isometrically with mass (approximately for most live vertebrates)36, bite force is predicted to scale to the two-thirds power of body mass (slope = 0.\(\mathop{6}\limits^{\bar{} }\))14. In C. cranwelli, the scaling exponents between bite force and linear morphometrics of body and head size, as well as between bite force and body mass, are significantly greater than theoretical predictions.

Teeth of Ceratophrys cranwelli. As with almost all other extant frogs bearing teeth, Ceratophrys has teeth only on the upper jaw. Unusual among frogs, the teeth of Ceratophrys exhibit a derived non-pedicellate morphology and have sharp recurved tips situated upon robust, labiolingually expanded bases. (a) view of teeth with jaws closed; (b) view of teeth with mouth slightly open; (c) close-up of single tooth. Scale bars = 1 mm.
The positive allometric relationship in Ceratophrys between bite force and body and head size is consistent with results found for other taxa examined to date. For example, positive intraspecific allometry between in vivo voluntary bite force and body and head morphometrics is observed in fish22, lizards13,17, tuatara37, alligators14, turtles17,21, and rodents38,39 (Table 1). Published intraspecific scaling analyses of voluntary bite force in mammals other than rodents are rare, but see analyses of scaling of bite force with age in hyenas (including scaling coefficients)40 and in coyotes41.
Our predictions of bite-force performance at two different jaw positions for C. aurita specimen LACM 163430 (~250–500 N) and Beelzebufo (~370–740 N [FMNH PR 2512, HW = 111 mm] and ~1100–2200 N [UA 9269, HW = 154 mm]) are consistent with previously published studies of bite force in other medium to large-sized vertebrates. Direct empirical in vivo measurements of voluntary bite force for vertebrates with heads of comparable size to that of C. aurita (LACM 163430) and Beelzebufo are uncommon. However, useful comparisons are afforded by several in vivo scaling analyses on turtles and crocodylians, as well as measurements and estimates of bite force for carnivorous mammals. A specimen of the large predatory turtle Chelydra serpentina (common snapping turtle), with a mass of 16.65 kg and a head width of 101 mm, was measured to bite with a force of 657 N17. This exceeds the estimated maximum bite force of C. aurita (LACM 163430) with a head of slight lesser width (98.3 mm, 497.1 N) by ~32%. Although the position in the jaw at which bite forces were measured was not reported (i.e., bite out-lever10), the scaling relationship provided by the authors predicts that a C. serpentina with a head 154 mm wide (~73.5 kg body mass) should bite with a force of 2042 N, similar to that of a Beelzebufo with a head of equal width (2213.7 N).
For a given head width, maximum bite-force performance in crocodylians is greater than calculated for C. aurita but less than calculated for Beelzebufo. For crocodylians with a head width of 98.3 mm, corresponding to C. aurita (LACM 163430), bite forces estimated at the position of the largest tooth at the posterior end of the tooth row are 689.3 N for Alligator mississippiensis (American alligator, interpolated from empirical data of bite force at 11th maxillary tooth; ref.14), 680.5 N for Crocodylus porosus (saltwater crocodile, interpolated), and 707.7 N for C. johnstoni (freshwater crocodile, extrapolated) (G. M. Erickson and AKL unpublished data, see ref.24 for relationships between bite force and body size). These crocodylians would have masses of 8.2 kg, 11.9 kg, and 9.4 kg, respectively. In all cases, our interpolation/extrapolation of bite force relative to head width for crocodylians exceed our bite force estimate for C. aurita (LACM 163430) by approximately 37–42%. For crocodylians with a head width of 154 mm, corresponding to that of the largest specimen of Beelzebufo, bite force estimates are 1659.6 N for Alligator mississippiensis, 1836.5 N for Crocodylus porosus, and 1863.6 N for C. johnstoni (G. M. Erickson and AKL unpublished data, see ref.24). These crocodylians would have masses of 24.2 kg, 34.0 kg, and 27.3 kg, respectively. In contrast to the comparison of bite force between crocodylians and C. aurita (LACM 163430), our estimate for the maximum bite force of Beelzebufo exceeds the bite force of crocodylians with a comparable head width by approximately 16–25%.
Our predictions for Beelzebufo also are compatible with available information on bite force in mammals. Published direct empirical measurements of voluntary bite forces in large mammals are surprisingly rare. Studies on captive colonies of two carnivorans, Crocuta crocuta (spotted hyaena)40 and Canis latrans (coyote)41, provide the only reliable empirical data. C. crocuta produced values of ~1000–4500 N for 6 + year old adults40. The greatest measurement for C. latrans, produced by an eight year-old male, was 704 N41. Bite forces estimated for Beelzebufo (UA 9269), both at the jaw tips and jaw midpoint, span the lower half of the range for C. crocuta and considerably exceed the maximum for C. latrans. Estimates of bite force derived from morphology-based models for Canis lupus (gray wolf), and Panthera leo (lion) and P. tigris (tiger) the size of adult females, are 774 N, 2024 N, and 2165 N, respectively32. Our estimates of bite force at the jaw tips for Beelzebufo (UA 9269) exceed the estimated bite force for C. lupus, the latter of which is unexpectedly similar to the in vivo measurement for the much smaller C. latrans. Bite force at the jaw midpoint for Beelzebufo (UA 9269) is similar to that estimated for the largest extant cats, P. leo and P. tigris.
Extant ceratophryid frogs are exceptional in having a disproportionately large head30, reflecting their megalophagous, ambush predatory tactics. They also have extremely wide skulls29, relative to skull length and depth30, which house large jaw adductor muscles. Although ceratophryids generally possess these characteristics, research on interspecific variation of the jaw-adductor mechanics of these frogs is warranted, including among species of Ceratophrys, which appear to exhibit appreciable interspecific variation of skull proportions.
Evaluating the biology of long-extinct animals, such as making estimates of their performance capacities, is challenging due to the incomplete nature of fossil material, as well as the desire to identify appropriate modern analogues. Although Beelzebufo is strikingly similar to Ceratophrys in many ways (e.g., unicuspid teeth, robust skull, cranial exostosis, absence of a palatine shelf on the premaxilla), available material suggests that its skull was relatively longer and shallower, which might also indicate differences in jaw muscle architecture28. Therefore, our estimates of its bite-force performance should be received with caution, as should be the case with any such estimate. Nevertheless, the head width we use for Beelzebufo (154 mm) may be less than the maximum possible as some fossil material corresponding to frogs of this size have unfused cranial sutures in contrast to the strongly fused cranial bones of adult Ceratophrys 27. Given that prey size is known to increase with body size in a variety of tetrapods42, and that Beelzebufo clearly had the ability to bite with considerable force, large individuals would have been able to prey upon a variety of contemporaneous taxa, including small/juvenile crocodiles and non-avian dinosaurs43.
Methods
Morphometrics
Body mass (BM) and body length (BL) were measured and recorded as indicators of body size. Head size was quantified by measuring three variables: head length from the jaw joint to the tip of the snout (HL), head width at the lateral extent of the jaw joints (HW), and head depth (HD) from the dorsal-most part of the skull just posterior of the orbits to the ventral extent of the lower jaw. All head measurements were made parallel or perpendicular to the antero-posterior or dorsal-ventral axis of the head to avoid the potentially confounding effects of angular measurements44. Linear measurements were made with digital calipers to the nearest 0.1 mm, and body mass was measured to the nearest 0.1 g with a digital scale.
Specimens and data sampling for in vivo bite-force experiments
Data were collected from eight individuals of the ceratophryid frog Ceratophrys cranwelli during post-metamorphic ontogeny. Specimens were purchased from commercial dealers and donated by private breeders. Data sampling followed a longitudinal experimental design. Over a period of one year of post-metamorphic growth, we conducted a series of experimental sessions (5–10 per animal, mean = 7.1) during which morphometric and bite-force data were collected. The pugnacious nature of Ceratophrys and its willingness to bite as a defensive response29,45, unusual among amphibians, makes this taxon an excellent subject for an analysis of bite force (Fig. 1). All procedures in this study were approved by the Animal Care and Use Committee of California State Polytechnic University, and the methods were carried out in accordance with the relevant guidelines and regulations.
Bite-force performance
Bite-force performance was measured using a piezoelectric force transducer (type 9203, Kistler, Switzerland) custom fitted with stainless steel bite plates and connected to a charge amplifier (type 5995, Kistler, Switzerland)11, but see refs10,46. The transducer was prepared for experiments by adhering leather strips at the ends of the plates where the frogs were induced to bite10 (Fig. 1). The leather served to protect the animals’ teeth and jaw bones from damage, provide a surface for the frog to grip with its jaws, and avoid the potential for reduced performance via sensory feedback if a non-naturalistic surface (e.g., steel) was used10. To calibrate amplifier output, a series of weights suspended with fishing line on the leather strip was used to reflect the force applied during bites to the same area. The distance between the plates and the thickness of the leather strips was adjusted as needed so that gape angle was approximately consistent among individuals (20–25 degrees). Three bite-force trials were performed on each frog during each experimental session, with one minute of rest between each trial. All frogs bit the transducer vigorously during all trials.
We standardized for variation, both among trials and among individuals, in the position along the jaw line that engaged the leather strips on the bite plates by using a simple lever calculation to compute bite force at two bite points10, at the tips of the jaws and at the antero-posterior midpoint of the lower jaw (i.e., midpoint between jaw tips and jaw joint), with the latter approximately corresponding to the location of the posterior teeth. These two bite points encompass the possible range of bite forces during a high-effort predatory episode.
Statistical analysis
All data were log-transformed and found to be normally distributed (Shapiro-Wilk W) for subsequent parametric statistical analyses47. Analyses were performed using JMP v8.0.2 and SAS v9.3.
Because our study was based on a longitudinal experimental design, we tested for individual variation in the relationship between bite force and each morphometric. To do this we used ANCOVA with bite force as the dependent variable, individual as the factor, morphometric as the covariate (separate model for each morphometric), and an individual by morphometric interaction. When the individual by morphometric (covariate) interaction was not significant, indicating that the slope of the relationship between the morphometric and bite force did not differ among the individual frogs, it was removed from the model. For these cases, the pooled data across all specimens is thus representative of the ontogenetic pattern for the individual frogs.
Tests of allometric scaling
To test the hypothesis that Ceratophrys exhibits a positive allometric increase in bite-force during ontogeny with respect to the measured morphological variables, we performed a reduced major axis (RMA) regression of log bite force on the log of each morphometric variable for each individual frog. We then pooled the slopes to produce a linear scaling coefficient for each bite force versus morphometric relationship. The standard error of the pooled RMA slope for each relationship was estimated by bootstrapping with 10000 iterations (i.e., 10000 bootstrap samples generated for each individual pooled to calculate 10000 slopes). Intercepts were calculated for bite positions at the jaw tips and at the jaw midpoint using regression means for the log of bite force on the morphometric at each bite position.
Estimates of jaw-adductor muscle size in Ceratophrys frogs and Beelzebufo
We examined the cranial musculoskeletal anatomy of Ceratophrys frogs to identify factors potentially significant in determining bite-force performance that also could be ascertained in skeletal or fossil material. The size of the adductor chamber of frogs is likely to have a strong association with potential bite-force performance. At the ventral end of the chamber on each side of the skull, bony elements form a conduit for the passage of the primary jaw-adductor musculature. This conduit can therefore be used as a proxy for the cross-sectional area of the adductor musculature (Fig. 3). The major jaw adductor muscles originate from the braincase and adjacent bones. From their origins, the muscle fibers run towards the palatal conduits, assume vertical or near vertical orientations via a pulley arrangement, and insert onto the dorsomedial and lateral aspects of the posterior end of the mandible. This pair of conduits through which the fibers of the major jaw-adductor muscles pass on their way to inserting onto the mandible provides an estimate of the cross-sectional area of the muscles that generate bite force, one that can be acquired from skeletal material alone and thus applied to comparable forms (i.e., C. aurita and Beelzebufo). We quantified and compared the area of the pair of palatal conduits among C. cranwelli, other Ceratophrys spp. including the exceptional C. aurita specimen, and Beelzebufo (Supplementary Figure S3), to assess relative similarity in the cross-sectional area of the major jaw-adductor musculature contributing to bite-force performance. Due to the incomplete nature of the existing Beelzebufo material, we made three estimates of palatal conduit area (conservative, likely, maximum; Fig. 3).
Extrapolation of bite forces for exceptional Ceratophrys and extinct Beelzebufo
Using the results of our scaling analysis, we extrapolated bite force for two larger frogs. First, we estimated the maximum bite force of an exceptionally large Ceratophrys specimen housed at the Natural History Museum of Los Angeles County (LACM 163430) (see ref.28; Supplementary Figure S4). This is a no data skeletal specimen indicated as C. varia, which is an invalid junior synonym for C. aurita (Brazilian Horned Frog)48,49. C. aurita is considered the largest species in the genus50, and we found that LACM 163430 has a body length of 178 mm (estimated by the length of the skull plus the articulated vertebral column and pelvic girdle) and a head width of 98.3 mm. We also make a prediction for the bite force of the extinct Beelzebufo ampinga from the Late Cretaceous of Madagascar. This is a large anuran (body length = 232 + mm, skull width = 111–154 + mm; ref.28; Supplementary Figure S5), which extensive phylogenetic analyses recovered as more closely related to ceratophryids than any other extant lineage27,28.
To estimate the bite force of LACM 163430 and of Beelzebufo, we determined the best morphometric predictor of bite force (i.e., head width – see Results) by running full model and stepwise multiple regressions with bite force as the dependent variable and all of the morphometrics as the independent variables. We then performed a reduced major axis (RMA) regression of log bite force on log head width, calculated the standard error of the pooled RMA, and used a bootstrapping method to calculate the intercepts for bite positions at the jaw tips and at the posterior extent of the tooth row, as described above. The linear relationship described by the model was used to calculate the potential bite-force performance of a Ceratophrys with a head width of 98.3 mm (LACM 163430), as well as to make two estimates of the bite force of Beelzebufo, one with a head width of 111 mm (FMNH PR 2512) and one with a head width of 154 mm (UA 9269; see ref.28).
Data availability
The data that support the findings of this study are available from the authors on reasonable request.
References
Romer, A. S. Major steps in vertebrate evolution. Science 158, 1629–1637 (1967).
de Beer, G. R. The Development of the Vertebrate Skull. (University of Chicago Press, 1985).
Mallatt, J. Crossing a major morphological boundary: the origin of jaws in vertebrates. Zoology 100, 128–140 (1997).
Depew, M. J. & Simpson, C. A. 21st Century neontology and the comparative development of the vertebrate skull. Dev Dyn 235, 1256–1291 (2006).
Anderson, P. S. L., Friedman, M., Brazae, M. D. & Rayfield, E. J. Initial radiation of jaws demonstrated stability despite faunal and environmental change. Nature 476, 206–209 (2001).
Nishikawa, K. C. Feeding in frogs. In Feeding Form, Function, and Evolution in Tetrapod Vertebrates (ed. Schwenk, K.) 117–147 (Academic Press, 2000).
Wake, D. B. & Deban, S. M. Terrestrial feeding in salamanders. In Feeding Form, Function, and Evolution in Tetrapod Vertebrates (ed. Schwenk, K.) 95–116 (Academic Press, 2000).
Kleinteich, T. & Gorb, S. N. Tongue adhesion in the horned frog Ceratophrys sp. Sci Rep 4, 5225 (2014).
Schalk, C. M., Montaña, C. G., Klemish, J. L. & Wild, E. R. On the diet of the frogs of the Ceratophryidae: synopsis and new contributions. S Amer J Herpetol 9, 90–105 (2014).
Lappin, A. K. & Jones, M. E. H. Reliable quantification of bite-force performance requires use of appropriate biting substrate and standardization of bite out-lever. J Exp Biol 217, 4303–4312 (2014).
Herrel, A., Spithoven, L., Van Damme, R. & De Vree, F. Sexual dimorphism of head size in Gallotia galloti: testing the niche divergence hypothesis by functional analyses. Func Ecol 13, 289–297 (1999).
Herrel, A., Joachim, R., Vanhooydonck, B. & Irschick, D. J. Ecological consequences of ontogenetic changes in head shape and bite performance in the Jamaican lizard Anolis lineatopus. Biol J Linn Soc 89, 443–454 (2006).
Meyers, J. J., Herrel, A. & Birch, J. Scaling of morphology, bite force, and feeding kinematics in an iguanian and a scleroglossan lizard. in Topics in Functional and Ecological Vertebrate Morphology (ed. Aerts, P., D’aout, K., Herrel, A. & Van Damme, R.) 47–62 (Shaker Publishing, 2002).
Erickson, G. M., Lappin, A. K. & Vliet, K. A. The ontogeny of bite-force performance in American alligator (Alligator mississippiensis). J Zool 260, 317–327 (2003).
Herrel, A. & Gibb, A. C. Ontogeny of performance in vertebrates. Physiol Biochem Zool 79, 1–6 (2006).
Hernandez, L. P. & Motta, P. L. Trophic consequences of differential performance in the sheepshead, Archsargus probatocephalus (Teleostei, Sparidae). J Zool 243, 737–756 (1997).
Herrel, A. & O’Reilly, J. C. Ontogenetic scaling of bite force in lizards and turtles. Physiol Biochem Zool 79, 31–42 (2006).
Huber, D. R., Weggelaar, C. L. & Motta, P. J. Scaling of bite force in the blacktip shark Carcharhinus limbatus. Zoology 109, 109–119 (2006).
Anderson, R. A., McBrayer, L. D. & Herrel, A. Bite force in vertebrates: opportunities and caveats for use of a nonpareil whole-animal performance measure. Biol J Linn Soc 93, 709–720 (2008).
van der Meij, M. A. A. & Bout, R. G. Scaling of jaw muscle size and maximal bite force in finches. J Exp Biol 207, 2745–2753 (2004).
Pfaller, J. B., Herrera, N. D., Gignac, P. M. & Erickson, G. M. Ontogenetic scaling of cranial morphology and bite-force generation in the loggerhead musk turtle. J Zool 280, 280–289 (2010).
Grubich, J. R., Huskey, S., Crofts, S., Orti, G. & Porto, J. Mega-bites: extreme jaw forces of living and extinct piranhas (Serrasalmidae). Sci Rep 2, 1009 (2012).
Marshall, C. D. et al. The ontogenetic scaling of bite force and head size in loggerhead sea turtles (Caretta caretta): implications for durophagy in neritic, benthic habitats. J Exp Biol 215, 4166–4174 (2012).
Erickson, G. M. et al. A comparative analysis of ontogenetic bite-force scaling among Crocodylia. J Zool 292, 48–55 (2014).
Barrio, A. Una nueva especie de Ceratophrys (Anura, Ceratophryidae) del dominio Chaqueno. Physis. Sección C: los continentes y los organismos terrestres 39, 21–30 (1980).
Frank, N. & Ramus, E. Complete Guide to Scientific and Common Names of Amphibians and Reptiles of the World. (N. G. Publishing Inc., 1995).
Evans, S. E., Jones, M. E. H. & Krause, D. W. A giant frog with South American affinities in the Late Cretaceous of Madagascar. Proc Nat Acad Sci 105, 2951–2956 (2008).
Evans, S. E., Groenke, J. R., Jones, M. E. H., Turner, A. H. & Krause, D. W. New material of Beelzebufo, a hyperossified frog (Amphibia: Anura) from the Late Cretaceous of Madagascar. PLoS ONE 9(1), e87236 (2014).
Emerson, S. B. Skull shape in frogs-correlations with diet. Herpetologica 41, 177–188 (1985).
Wild, E. R. Description of the adult skeleton and developmental osteology of the hyperossified horned frog, Ceratophrys cornuta (Anura: Leptodactylidae). J Morphol 232, 169–206 (1997).
Fabrezi, M. et al. Developmental changes and novelties in ceratophryid frogs. Evo Devo 7, 5 (2016).
Christiansen, P. & Wroe, S. Bite forces and evolutionary adaptations to feeding ecology in carnivores. Ecology 88, 347–358 (2007).
Wroe, S., McHenry, C. & Thomason, J. Bite club: comparative bite force in big biting mammals and the prediction of predatory behaviour in fossil taxa. Proc R Soc B 272, 619–625 (2005).
Lynch, J. D. Relationships of the frogs of the genus Ceratophrys (Leptodactylidae) and their bearing on hypotheses of Pleistocene forest refugia in South America and punctuated equilibria. Syst Zool 31, 166–179 (1982).
Smirnov, S. V. & Vasil’eva, A. B. Anuran dentition: development and evolution. Russian J Herpetol 2, 120–128 (1995).
Schmidt-Nielsen, K. Scaling: Why is Animal Size so Important? (Cambridge University Press, 1984).
Jones, M. E. H. & Lappin, A. K. Bite-force performance of the last rhynchocephalian (Lepidosauria: Sphenodon). J Royal Soc New Zealand 39, 71–83 (2009).
Becerra, F., Echeverría, A., Vassallo, A. I. & Casinos, A. Bite force and jaw biomechanics in the subterranean rodent Talas tuco-tuco (Ctenomys talarum) (Caviomorpha: Octodontoidea). Can J Zool 89, 334–342 (2011).
Becerra, F., Casinos, A. & Vassallo, A. I. Biting performance and skull biomechanics of a chisel tooth digging rodent (Ctenomys tuconax; Caviomorpha; Octodontoidea). J Exp Zool 319A, 74–85 (2013).
Binder, W. J. & Van Valkenburgh, B. Development of bite strength and feeding behaviour of juvenile spotted hyenas. J Zool 252, 273–283 (2000).
La Croix, S., Zelditch, M. L., Shivik, J. A., Lundrigan, B. L. & Holekamp, K. E. Ontogeny of feeding performance and biomechanics in coyotes. J Zool 285, 301–315 (2011).
Vézina, A. F. Empirical relationships between predator and prey size among terrestrial vertebrate predators. Oecologia 67, 555–565 (1985).
Krause, D. W. et al. Late Cretaceous terrestrial vertebrates from Madagascar: implications for Latin American biogeography. Ann Missouri Bot Gard 93, 178–208 (2006).
Lappin, A. K. & Swinney, E. J. Sexual dimorphism as it relates to the natural history of leopard lizards (Crotaphytidae: Gambelia). Copeia 1999, 649–660 (1999).
Duellman, W. E. & Lizana, M. Biology of a sit-and-wait predator, the leptodactylid frog Ceratophrys cornuta. Herpetologica 50, 51–64 (1994).
Lappin, A. K. & Husak, J. F. Weapon performance, not size, determines mating success and potential reproductive output in the collared lizard (Crotaphytus collaris). Am Nat 166, 426–436 (2005).
Sokal, R. R. & Rohlf, F. J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed. (W. H. Freeman and Co., 1995).
Bokermann, W. C. A. Lista Anotada das Localidades Tipo de Anfíbios Brasileiros. (Servicio de Documentacão, 1966).
Frost, D. R. Amphibian Species of the World: A Taxonomic and Geographic Reference. (Allen Press, 1997).
de Vosjoli, P. The General Care and Maintenance of Horned Frogs. (California: Advanced Vivarium Systems, 1989).
Acknowledgements
We thank Brigitte Demes, David Krause, Marci Hollenshead, Roy Heng, Molly Peters, Par Singhaseni, Katherine Belessiotis, and Cassandra Stepp-Bolling for discussion and comments on earlier versions of this manuscript, and Philippe de Vosjolli for the donation of specimens used in this research. For access to skeletal specimens, we thank Gregory Pauly and Neftali Camacho (LACM), Matthew Lowe (Cambridge University Museum of Zoology), Mark Carnall (UCL Grant Museum), and Barry Clarke (Natural History Museum London). We thank Joseph Groenke (Ohio University) for use of CT scans of skeletal specimen LACM 163430, which was originally loaned to David Krause (Stony Brook University, NY). This research was supported by the HHMI Undergraduate Research-Apprentice Program, the College of Science and Biological Sciences Department at California State Polytechnic University, and a Discovery Early Career Researcher Award DE130101567 (MEHJ) (Australian Research Council).
Author information
Authors and Affiliations
Contributions
A.K.L., S.C.W., S.E.E., and M.E.H.J. designed the study. A.K.L., S.C.W., and S.A.R.S. collected the data. A.K.L. and D.J.M. analyzed the data and ran the statistical analyses. A.K.L. drafted the paper. All authors contributed to interpreting results, developing content, and (except S.A.R.S. in memoriam) editing the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lappin, A.K., Wilcox, S.C., Moriarty, D.J. et al. Bite force in the horned frog (Ceratophrys cranwelli) with implications for extinct giant frogs. Sci Rep 7, 11963 (2017). https://doi.org/10.1038/s41598-017-11968-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-11968-6
This article is cited by
-
A bite force database of 654 insect species
Scientific Data (2024)
-
The radiation continuum and the evolution of frog diversity
Nature Communications (2023)
-
Interspecific size- and sex-related variation in the cranium of European brown frogs (Genus Rana)
Zoomorphology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
