
Abstract
Adipogenesis plays an important role in the regulation of whole-body energy homeostasis and is inextricably related to obesity. Several studies have highlighted the relevance of microRNAs in adipocyte differentiation, but the contributions of long non-coding RNAs (lncRNAs) are still largely uncharacterized. Here, we determined that lncRNA MIR31HG is related to adipocyte lineage commitment. We demonstrated that knockdown of MIR31HG inhibited adipocyte differentiation, whereas overexpression of MIR31HG promoted adipogenesis in vitro and in vivo. Furthermore, inhibition of MIR31HG reduced the enrichment of active histone markers, histone H3 lysine 4 trimethylation (H3K4me3) and acetylation (AcH3), in the promoter of the adipogenic-related gene, fatty acid binding protein 4 (FABP4), leading to suppression of its expression and adipogenesis. These results provide new insights into the molecular mechanisms of MIR31HG in terms of adipogenesis and may have implications for obesity and associated disorders.
Similar content being viewed by others


Introduction
Obesity is an important factor in various diseases, particularly heart disease, diabetes, hypertension, and cancer1, 2. The global prevalence of obesity has greatly increased and constitutes a public health crisis3. Adipogenesis plays key roles in the regulation of whole-body energy homeostasis and is inextricably related to obesity. It is a complicated process, involving the proliferation of precursor cells, their commitment to the adipogenic lineage, and terminal differentiation4. A clearer understanding of the molecular mechanisms that initiate differentiation of stem cells into adipocytes would facilitate the development of methods for the treatment of obesity and other adipogenic-differentiation-related disorders. Adipose-derived stem cells (ASCs) are a ready and ideal cell model for human adipogenesis. The ability of ASCs to differentiate into mature adipocytes has been demonstrated5, 6.
Recently, emerging studies of non-protein-coding RNAs (ncRNAs) in adipocyte commitment provide new insights into the molecular basis of adiposity. A class of small ncRNAs, including miR-25 7, miR-29b 8, miR-93 9, miR-148a 10, and miR-1275 11, have been identified as important regulators of adipogenic differentiation by targeting various genes involved in cell self-renewal and differentiation. In contrast, the functional contributions of long non-coding RNAs (lncRNAs), tentatively defined as ncRNAs > 200 nucleotides in length12, 13, in ASC adipogenic differentiation remain largely unknown. A recent study has profiled the transcriptome of mature adipocytes and pre-adipocytes and identified 175 lncRNAs that are differentially expressed during adipogenesis14. However, only a few lncRNAs, such as ADNCR 15, ADINR 16, PU.1-as 17, and NEAT1 18, have been shown to participate in the genetic control of adipogenic differentiation of ASCs or pre-adipocytes. The importance and potential roles of most lncRNAs in adipocyte lineage commitment remain uncharacterized.
The miR-31 host gene (MIR31HG, GenBank accession number, NR_027954), located on chromosome 9 (9p21.3), is transcribed into an lncRNA19. MIR31HG has been reported to affect cell proliferation in various cancers19,20,21,22, and our previous study shows that it can affect osteoblast differentiation23, indicating an important biological function of MIR31HG in cellular growth and differentiation. In this study, we determined the role of MIR31HG in adipocyte differentiation. We found that knockdown of MIR31HG inhibited adipogenesis of human ASCs (hASCs) in vitro and in vivo, whereas overexpression of MIR31HG promoted this process. In mechanism, MIR31HG participated in histone methylation and acetylation of fatty acid binding protein 4 (FABP4). These results reveal new insights into the mechanisms of adipogenic differentiation and provide a potential molecular target for the treatment of obesity and related diseases.
Results
LncRNA MIR31HG is involved in adipocyte differentiation of hASCs
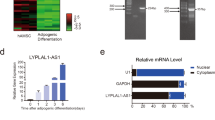
To gain an understanding of MIR31HG, we firstly studied the expression pattern of MIR31HG in human tissues. RNA-seq data from NONCODE (http://www.noncode.org) indicated that MIR31HG is expressed in a number of tissues or cell lines, such as foreskin, kidney, prostate, testes, hela cells, etc (Fig. 1A). Conservation information showed that MIR31HG is expressed in several mammals, including human, orangutan, and cow, but absent in other chordates (Fig. 1B). Single-molecule RNA fluorescence in situ hybridization of MIR31HG showed that the transcript was localized both in the nucleus and cytoplasm of hASCs, predominantly in the nucleus (Fig. 1C). Its localization was also confirmed by nuclear/cytoplasm fractionation (Fig. 1D).

Characterization of lncRNA MIR31HG. (A) Expression profile of MIR31HG in human tissues. FPKM: fragments per kilobase of transcript per million fragments mapped. (B) Conservation information of lncRNA MIR31HG. Chr: chromosome. (C) Confocal FISH images showing localization of MIR31HG in hASCs. Scale bars: 20 μm. (D) The expression of MIR31HG, GAPDH, and MALAT-1 in nuclear or cytoplasmic fraction of hASCs. GAPDH and MALAT-1 were set as controls. Results are presented as mean ± SD (**P < 0.01).
To study the functional role of MIR31HG during adipogenesis, we analyzed MIR31HG expression profile during adipocyte differentiation of hASCs. MIR31HG exhibited significantly increased expression after 2 days compared to undifferentiated hASCs and returned to normal levels after 4 days (Fig. 2A). The expression of the genes associated with adipogenic differentiation, peroxisome proliferator-activated receptors -γ (PPARγ), CCAAT/enhancer-binding protein α (C/EBPα), and FABP4, was substantially upregulated after adipogenic differentiation induction (Fig. 2B and C).

Dynamic Expression profile of MIR31HG during adipogenic differentiation of hASCs. (A) Relative expression of MIR31HG on day 2, 4, and 6 during differentiation as determined by qRT-PCR analysis normalized to the undifferentiation group cultured in growth medium (GM). (B) Relative mRNA expression levels of the adipogenic markers PPARγ, C/EBPα, and FABP4 at the indicated time points, as in (A). Gene expression plotted as fold-change relative to the GM group. (C) Western blot analysis (left) and quantification (right) of protein expression of PPARγ, C/EBPα, FABP4, and the internal control GAPDH at the indicated time points, as in (A). Results are presented as mean ± SD (*P < 0.05, **P < 0.01 compared to the undifferentiated hASCs).
MIR31HG promotes cell proliferation
To further study the biological function of MIR31HG, we stably overexpressed and knocked down MIR31HG in hASCs using lentivirus transfection. Two different shRNA sequences targeting MIR31HG were designed to control for potential off-target shRNA effects. The transduced hASCs did not exhibit morphological changes compared with the control. The efficiency of lentiviral transduction was >90% (Fig. 3A), and the expression of MIR31HG was significantly reduced by ~70% in the knockdown group and increased more than 50-fold in the overexpressed group (Fig. 3B).

MIR31HG promoted cell proliferation of hASCs. (A) The hASCs were transfected with lentivirus expressing shMIR31HG-1, shMIR31HG-2, the scrambled nontargeting vector (shNC), MIR31HG, or the scrambled vector (NC). Fluorescent photomicrographs show the efficiency of lentivirus transduction (>90%). Scale bar, 500 μm. (B) Relative expression of MIR31HG in the shMIR31HG groups (up) or the MIR31HG group (down). (C) CCK-8 assay showing cell proliferation in the shMIR31HG groups (up) or the MIR31HG group (down). Results are presented as mean ± SD (*P < 0.05, **P < 0.01).
Since MIR31HG expression was induced at an early stage of adipogenic differentiation, we determined the biological function of MIR31HG in cell proliferation. According to the CCK-8 assay results, we found that MIR31HG overexpression slightly promoted cell proliferation, while MIR31HG knockdown decreased cell growth by ~30% relative to negative control at 48 h and 72 h in hASCs (Fig. 3C).
Knockdown of MIR31HG inhibits adipocyte differentiation
We then investigated the impact of MIR31HG on adipogenic differentiation. We stably knocked down endogenous MIR31HG in hASCs. After induction to the adipogenic lineage, MIR31HG knockdown significantly reduced adipocyte numbers, as indicated by Oil red O staining on day 8 of differentiation (Fig. 4A). Accordingly, the mRNA levels of the adipocyte-specific markers, PPARγ, C/EBPα, and FABP4, were substantially inhibited by the depletion of MIR31HG (Fig. 4B). Consistently, the protein levels of PPARγ, C/EBPα, and FABP4 were downregulated in hASCs with MIR31HG knockdown on day 4 of adipogenic induction (Fig. 4C).

Knockdown of MIR31HG inhibited adipogenic differentiation. The hASCs were transfected with lentivirus expressing shMIR31HG-1, shMIR31HG-2, or the scrambled nontargeting vector (shNC). (A) Images of Oil red O staining on day 8 of adipocyte differentiation. Scale bar, 100 μm. Histograms show quantification of Oil red O staining by spectrophotometry (normalized to shNC group). (B) Relative mRNA expression of the adipogenic factors PPARγ, C/EBPα, and FABP4 measured by qRT-PCR on day 4 of adipogenic induction. (C) Western blot analysis of PPARγ, C/EBPα, FABP4, and GAPDH on day 4 of adipogenic induction. Histograms show quantification of the band intensities. Results are presented as mean ± SD (*P < 0.05, **P < 0.01).
Overexpression of MIR31HG promotes adipocyte differentiation
We stably overexpressed MIR31HG in hASCs. In the presence of adipogenic agents, MIR31HG transfection into hASCs led to a significant increase in adipocyte formation, as shown by Oil red O staining on day 8 of differentiation (Fig. 5A). Consistently, MIR31HG substantially increased the mRNA (Fig. 5B) and protein (Fig. 5C) levels of the adipogenic genes, PPARγ, C/EBPα, and FABP4 on day 4 of adipogenic differentiation. Interestingly, we found that even without adipogenic treatment, supplementation with MIR31HG alone increased the expression of adipogenic factors, PPARγ, C/EBPα, and FABP4, as examined by qRT-PCR (Fig. 5D) and Western blot (Fig. 5E). However, MIR31HG alone was not sufficient to induce intracellular lipid accumulation without adipogenic supplements as indicated by Oil red O staining (Supplementary Figure 1).

MIR31HG promoted adipogenic differentiation. The hASCs were transfected with lentivirus expressing MIR31HG or the scrambled vector (NC). (A) Images of Oil red O staining on day 8 of adipocyte differentiation. Scale bar, 100 μm. Histograms show quantification of Oil red O staining by spectrophotometry (normalized to NC group). (B) Relative mRNA expression of the adipogenic genes PPARγ, C/EBPα, and FABP4 on day 4 of differentiation. (C) Western blot analysis (left) and quantification (right) of protein expression of PPARγ, C/EBPα, FABP4, and GAPDH on day 4 of differentiation. (D) Relative mRNA expression of the adipogenic genes PPARγ, C/EBPα, and FABP4 without adipogenic differentiation. (E) Western blot analysis (left) and quantification (right) of protein expression of PPARγ, C/EBPα, FABP4, and GAPDH without adipogenic differentiation. Data are presented as mean ± SD (*P < 0.05, **P < 0.01).
Histone modification of the FABP4 promoter region by MIR31HG
One way that lncRNAs acquire functionality is by interacting with heteronuclear proteins or chromatin modification complexes24. We determined the effect of MIR31HG on histone modification of adipogenic transcription factors. Combined the results of qRT-PCR and Western blot analysis, we found that MIR31HG had significant effect on FABP4 (Figs 4 and 5), we thus examined local histone H3 acetylation and methylation in chromatin associated with the FABP4 promoter region using chromatin immunoprecipitation (ChIP) assay. The histone-associated DNAs, immunoprecipitated with antibodies against AcH3 and H3K4me3, were individually amplified with five primer sets covering the FABP4 promoter region (Fig. 6A). The results showed the marked differences in the levels of histone H3 acetylation and methylation in MIR31HG knockdown or overexpressed cells compared to the control group. The level of AcH3 was decreased in MIR31HG knockdown cells and increased in MIR31HG overexpression cells in the region containing primers 1–4 (Fig. 6B), while the level of H3K4me3 was decreased in MIR31HG knockdown cells and increased in MIR31HG overexpression cells in the fragment containing primers 2–5 (Fig. 6C).

Inhibition of MIR31HG reduced the levels of AcH3 and H3K4me3 in the FABP4 promoter. (A) Diagram of the FABP4 promoter and location of the primers. Positions marked are relative to the transcriptional start site (TSS). (B) ChIP assay of AcH3 in the FABP4 locus in hASCs with MIR31HG knockdown (shMIR31HG-1), MIR31HG overexpression (MIR31HG), or the scrambled vector (NC). Quantitative PCR data of each group were normalized to its input as % input. IgG served as negative control. (C) ChIP assay of H3K4me3 in the FABP4 locus in hASCs with MIR31HG knockdown (shMIR31HG-1), MIR31HG overexpression (MIR31HG), or the scrambled vector (NC). Quantitative PCR data of each group were normalized to its input as % input. IgG served as negative control. (D) Images of Oil red O staining on day 8 of adipogenic differentiation in hASCs with MIR31HG overexpression (MIR31HG) with or without FABP4 knockdown (shFABP4). Histograms show quantification of Oil red O staining by spectrophotometry (normalized to NC group). Data are presented as mean ± SD (*P < 0.05).
Knockdown of FABP4 abolishes the pro-adipogenic effect of MIR31HG
To determine whether MIR31HG regulates adipogenesis via histone modification of FABP4 gene, we knocked down FABP4 expression in hASCs with MIR31HG stably overexpression. After adipogenic induction, knockdown of FABP4 abolished the promotion of adipogenesis induced by MIR31HG, as indicated by Oil red O staining on day 8 of differentiation (Fig. 6D).
MIR31HG promotes newly formed adipose tissue in vivo
To determine the role of MIR31HG in vivo, we subcutaneously implanted hASCs stably expressing MIR31HG, shMIR31HG-1, or control mixed with a collagen scaffold in the dorsum of nude mice (Fig. 7A). The neo-generated tissue was histologically composed of numerous lobule-like structures separated from each other by fibrous tissue 8 weeks after implantation. The progressive development of adipose tissue was further characterized by Oil red O staining, showing intracellular lipid accumulation. The green fluorescent protein (GFP) -positive stem cells were tracked using fluorescence microscope. The MIR31HG group had more adipose-tissue-like constructs stained positively for Oil red O, while the shMIR31HG group had fewer lipid droplets stained for Oil red O (Fig. 7B). Quantification of the newly generated adipose tissue consistently showed a greater ratio of adipogenesis in the MIR31HG group (63.5%) and a lower ratio in the shMIR31HG group (9.2%) compared to control group (28.6%) (Fig. 7C). The qRT-PCR analysis of total RNA from the newly formed tissues showed that the FABP4 expression was reduced in the shMIR31HG group, while increased in the MIR31HG group (Fig. 7D).

MIR31HG knockdown inhibited adipose tissue formation in vivo. (A) Schematic diagram illustrating the experimental setup. (B) Fluorescent images, H&E staining, and Oil red O staining of the specimens in hASC/Collagen group (implanted with non-induced hASC/Collagen complex), Adi-hASC/Collagen group (implanted with adipogenic-induced hASC/Collagen complex), shMIR31HG-hASC/Collagen group (implanted with adipogenic-induced shMIR31HG-hASC/Collagen complex), and MIR31HG-hASC/Collagen group (implanted with adipogenic-induced MIR31HG-hASC/Collagen complex). Scale bar, 25 μm. (C) Quantitative analysis of the Oil red O-stained areas of newly generated tissue at 8 weeks after implantation. The areas were expressed as percentages of the total areas. (D) Relative mRNA expression of FABP4 in the newly generated tissue from indicated groups (normalized to Adi-hASC/Collagen group). Data are presented as mean ± SD (*P < 0.05, **P < 0.01).
Discussion
In this study, we demonstrated that knockdown of MIR31HG inhibited the adipocyte differentiation of hASCs, whereas overexpression of MIR31HG promoted the commitment of adipocytes. MIR31HG can also affect cell proliferation of hASCs, consistent with several studies of MIR31HG in carcinogenesis19,20,21,22. Moreover, our previous study shows that inhibition of MIR31HG expression promotes osteogenesis23. Taken together, MIR31HG may be involved in balancing self-renewal and stem cell differentiation. Interestingly, a previous study has subcutaneously injected recombinant adenovirus expressing Adamts1 shRNA adjacent to the inguinal fat pad in a preclinical mouse model, and found that the mice show enlargement of inguinal fat pads, greater weight, and impaired insulin sensitivity and glucose tolerance compared to control mice25. Our study indicating the role of MIR31HG in adipocyte commitment may also provide a potential target for the treatment of obesity through subcutaneous injection of shMIR31HG. On the other hand, engineering adipose tissue is an alternative to plastic and reconstructive surgery for restoring body contours in patients who have lost soft tissue because of external injury, congenital malformation, or surgical resection5, 6. Supplementation of MIR31HG in hASCs combined with a degradable scaffold may serve as a promising approach to improve adipogenesis in adipose tissue engineering.
MIR31HG contributed to histone H3 methylation and acetylation of the FABP4 gene. Histone modifications are significant reversible mechanisms of epigenetic regulation of gene expression. Dramatic changes in epigenetic signatures have been observed during adipogenesis26,27,28. A previous study has profiled the dynamic histone modifications of key adipogenesis regulatory genes during adipocyte commitment, and found that both H3 acetylation and K4 trimethylation levels increase in all the induced adipogenic genes and correlate positively with gene activity27. The most obvious increase is observed in the FABP4 gene27. This study supports our results showing that knockdown of MIR31HG reduced the enrichment of AcH3 and H3K4me3 in the FABP4 promoter, leading to suppression of FABP4 expression. And, knockdown of FABP4 expression abolished the effect of MIR31HG in adipogenesis. Adipocyte differentiation is a highly regulated process governed by PPARγ 29, 30, which cooperates with C/EBP to induce the expression of many genes important for terminal differentiation, such as FABP4 and CD36 31. FABP4 is highly expressed in adipose tissue and is able to bind and transport fatty acids and other lipophilic compounds32. In mouse models, FABP4 deficiency partially protects mice against the development of insulin resistance associated with genetic or diet-induced obesity33, 34. Previous studies suggest that targeting FABP4 with inhibitors is possible to prevent and treat metabolic diseases, such as type II diabetes and atherosclerosis35. Thus, MIR31HG may serve as a molecular target for metabolic diseases at least partially through suppression of FABP4. However, during the adipocyte differentiation, the expression of MIR31HG was transiently upregulated in the early stage (day 2) and returned to normal levels in the late stage (day 6). This expression pattern did not correlate with the expression pattern of adipogenic markers. Many different events contribute to adipocyte determination and differentiation including the coordination of a complex network of transcription factors, cofactors and signaling intermediates4. We cannot exclude the possibility that other transcription factors are simultaneously involved in regulation of PPARγ, C/EBPα, and FABP4, resulting in the inconsistent expression pattern with MIR31HG. Further study is needed to identify the possible mechanism.
LncRNAs can act as regulatory devices through combinations of modular functional components that include RNA-, DNA-, and protein-binding domains. They can create a scaffold for chromatin-modifying enzymes to epigenetically affect genomic loci24. For example, the lncRNAs ANRIL 36 and HOTAIR 37, 38 physically associate with a subunit of the polycomb repressive complex 2 to trimethylate H3K27 and specifically repress target genes. Similarly, the lncRNA HOTTIP binds WD repeat containing protein 5, a cofactor to histone methyltransferase MLL1 that mediates H3K4 trimethylation associated with gene activation39. A recent study has found the lncRNA ADINR regulates adipogenesis by transcriptional activation of C/EBPα. This lncRNA binds PA1 and recruits MLL3/4 complexes in the C/EBPα promoter to maintain H3K4me3 and remove H3K27me3 in these regions during adipogenic differentiation16. MIR31HG may exert its function by a similar mechanism. We have previously indicated several stable stem-loop structures in MIR31HG 23, which may provide the necessary spatial conformation for interaction with chromatin-modifying proteins. However, the modification complex recruited by MIR31HG to chromatin needs further investigation.
In conclusion, our study highlights the role of MIR31HG in adipocyte differentiation of hASCs. Knockdown of MIR31HG inhibits the adipocyte lineage commitment of stem cells. In mechanism, MIR31HG contributes to the enrichment of AcH3 and H3K4me3 in FABP4 gene and promotes its transcription, leading to the promotion of adipogenesis (Fig. 8). Further research is needed to elucidate the potential use of MIR31HG as a molecular target for treating adiposity.

Schematics showing the regulation of adipogenesis by MIR31HG. MIR31HG promoted adipogenic differentiation of stem cells. In mechanism, MIR31HG contributed to histone H3 lysine 4 trimethylation and H3 acetylation at the FABP4 gene locus and promoted its transcription, leading to the promotion of adipogenesis.
Methods
Cell culture
Primary hASCs from three different healthy human donors were obtained from ScienCell (San Diego, CA, USA) and their lot number were 2447, 8278, and 11537, respectively. The cells were cultured in growth medium (GM) consisting of DMEM with 10% fetal bovine serum and 1% antibiotics. All cell-based in vitro experiments were repeated in triplicate. For the adipocyte differentiation experiment, cells were allowed to become confluent for 1 day, and then cultured in standard growth medium supplemented with 10 μg/ml insulin (Sigma-Aldrich, St Louis, MO, USA), 100 nM dexamethasone (Sigma-Aldrich), 0.5 mM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), and 200 μM indomethacin (Sigma-Aldrich). The adipogenic medium was changed every 2 days, and cells were harvested at the indicated times.
Lentivirus infection
Recombinant lentiviruses containing full-length MIR31HG or the scrambled control (NC) were obtained from Integrated Biotech Solutions Co. (Ibsbio Co., Shanghai, China). Recombinant lentiviruses targeting MIR31HG (shMIR31HG-1 and shMIR31HG-2) or the scrambled nontargeting vector (shNC) were obtained from GenePharma Co. (Shanghai, China). Recombinant lentiviruses targeting FABP4 (shFABP4) was obtained from Integrated Biotech Solutions Co. Transfection of the hASCs was performed by exposing them to dilutions of the viral supernatant in the presence of polybrene (5 μg/mL) for 72 h, as described previously23. The recombinant lentivirus vectors contain the puromycin-resistance and GFP gene. 48 hours post transfection, puromycin was added to the cells at concentration of 10 µg/ml. The drug-resistant stable transfectants that express the gene of interest were selected. The transfection efficiency was then verified by fluorescence microscope tracking GFP, and the sorted cells were used as the stably expressing model in subsequent experiments.
Cell viability analysis
The cell viability detection was performed using Cell Counting Kit-8 (CCK-8, Dojindo, Kumamoto, Japan) according to manufacturer’s instructions. Briefly, cells were seeded in 96-well plate (5 × 103 cells/well). 10 ml of CCK-8 solution was added into each well at 24, 48, and 72 h after culture. Subsequently, the plates were further incubated for 1 h, then the absorbance was measured at 450 nm using a microplate spectrophotometer (Bio-Tek Instruments Inc., Winosski, VT).
Oil red O staining
Oil red O staining was performed as described previously40. Cells were washed with phosphate-buffered saline (PBS) and fixed in 10% formalin for 30 min. The cells were subsequently rinsed with 60% isopropanol. Oil red O (0.3%, Sigma-Aldrich) was then added and incubated for 10 min with gentle agitation. After staining, the cells were washed with distilled water to eliminate unbound dye, visualized by light microscopy, and photographed. For quantitative assessment, Oil red O was eluted by 100% isopropanol and quantified by spectrophotometric absorbance at 520 nm against a blank (100% isopropanol).
RNA isolation and qRT-PCR
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions and then reverse-transcribed into cDNA using the cDNA Reverse Transcription Kit from Applied Biosystems (Foster City, CA, USA). qRT-PCR was conducted with 1 μg of total RNA using the SYBR Green Master Mix on an ABI Prism 7500 real-time PCR System (Applied Biosystems) as described41. The following thermal settings were used: 95 °C for 10 min followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. The primers used for MIR31HG, PPARγ, C/EBPα, FABP4, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH, internal control for mRNAs and lncRNAs) are listed in Supplementary Table S1. The data were analyzed using the 2−ΔΔCt relative expression method as described previously41.
Western blot analysis
Western blot analysis was performed as described previously41. Briefly, cells were harvested, washed with PBS, and lysed in RIPA buffer. Proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane by electroblotting. After blocking, proteins were detected by overnight incubation with primary antibodies against PPARγ (Cell Signaling Technology, Beverly, MA, USA), C/EBPα (HuaxingBio Science, Beijing, China), FABP4 (HuaxingBio Science), and GAPDH (Abcam, Cambridge, UK) at dilutions of 1:1,000. After washing, the membranes were incubated with secondary antibodies (Zhongshan Goldenbridge, Beijing, China, 1:10,000 dilution) at room temperature for 1 hour. The specific complexes were visualized using the ECL Kit (Applygen, Beijing, China). The intensities of the bands obtained by Western blot analysis were quantified using ImageJ software (http://rsb.info.nih.gov/ij/). The background was subtracted, and the signal of each target band was normalized to that of the GAPDH band.
Fluorescent in situ hybridization (FISH)
In situ hybridization was performed with a Fluorescent In Situ Hybridization Kit (RiboBio, Guangzhou, China), as described previously23. Briefly, cells were fixed in 4% formaldehyde, permeabilized with 0.5% Triton X-100 for 5 min, and prehybridizated at 37 °C for 30 min before hybridization. Then an anti-MIR31HG oligodeoxynucleotide probe was used in the hybridization solution at 37 °C overnight in the dark. The next day, the cells were counterstained with DAPI and imaged using a confocal laser-scanning microscope (Carl Zeiss, Oberkochen, Germany).
Cell fractionation
For the fractionation assay, cytoplasmic and nuclear RNAs were performed using a Nuclei Isolation Kit (KeyGEN, Nanjing, China), as described previously23. Briefly, cells were harvested and resuspended in lysis buffer. Then, the lysate was treated with Reagent A, incubated on ice for 15 min, followed by centrifugation at 4 °C. The pellet was then resuspended in lysis buffer followed by centrifugation. The supernatant was transferred to a new tube as the cytoplasmic fraction; the pellet was resuspended in Medium Buffer A and then added to a new tube with Medium Buffer B, followed by centrifugation at 4 °C. The supernatant was saved as the cytoplasmic fraction. The pellet was used as the nuclear fraction. RNA was extracted from both fractions using TRIzol.
ChIP assay
ChIP assays were performed using the EZ-Magna ChIP assay kit (Merck Millipore, Darmstadt, Germany) according to the manufacturer’s instructions. Briefly, cells were washed with PBS and cross-linked with 1% formaldehyde for 10 min. Chromatin was sonicated on ice to generate chromatin fragments of 200–1000 bp, and then immunoprecipitated with each antibody. Antibodies for acetylated histone H3 (AcH3, Merck Millipore), tri-methylated histone H3K4 (H3K4me3, Merck Millipore), and isotype IgG (negative control, Cell Signaling Technology) were used in the immunoprecipitations. The immunoprecipitated materials were then washed extensively, and cross-linking was reversed. DNA from the eluted chromatin was purified by phenol extraction and ethanol precipitation. Input control DNA or immunoprecipitated DNA was determined by quantitative real-time PCR. Standard curves were constructed using a pool of input samples, and each ChIP sample was normalized to its respective input. We designed primers to separately amplify five regions in the FABP4 promoter region. The primer pairs used for ChIP assays are shown in Supplementary Table S1.
In vivo adipose tissue formation assay
The cells were adipogenically induced for 1 week before the in vivo study. Cell suspensions containing 5 × 106 hASCs were incubated with Collagen Sponge (8 mm × 8 mm × 2 mm) for 2 h at 37 °C to allow the cells to attach to the scaffold. A total of 40 5-week-old BALB/c homozygous nude (nu/nu) mice were obtained from Charles River Laboratories (Beijing, China). The mice were maintained under specific pathogen-free conditions and randomized into four groups of 10 mice each: hASC/Collagen group (implanted with non-induced hASC/Collagen complex), Adi-hASC/Collagen group (implanted with adipogenic-induced hASC/Collagen complex), shMIR31HG-hASC/Collagen group (implanted with adipogenic-induced shMIR31HG-hASC/Collagen complex), and MIR31HG-hASC/Collagen group (implanted with adipogenic-induced MIR31HG-hASC/Collagen complex). The complex was implanted subcutaneously in the dorsum of the nude mice. All animal experiments were approved by the Peking University Animal Care and Use Committee (No. LA2014233). All experiments were performed in accordance with relevant guidelines and regulations.
Histological observation
Neo-generated tissues were carefully dissected from the surrounding tissue 8 weeks after implantation. The harvested specimens were fixed in 4% paraformaldehyde and cut in half. One half was embedded in paraffin, and sectioned at 5 μm thickness for hematoxylin and eosin (H&E) staining, while the other half was frozen in Tissue-Tek OCT freezing medium (Sakura Finetek Inc., Torrance, CA, USA) and sectioned at 8 μm thickness for Oil red O staining.
Quantification of newly generated adipose tissue
The proportion of the positive area for Oil red O was calculated by Lumina Visionâ (Mitani Corp., Tokyo, Japan) for quantification. Twenty fields at 40 × magnification in each microscopic photograph were chosen in a random fashion and the positive area was measured by the software. The percent square of the positive area was used as an indication of the degree of adipogenesis.
Statistical analysis
Statistical analyses were performed using SPSS version 16.0 (SPSS, Chicago, IL, USA). All data are expressed as mean ± standard deviation (SD). Differences between groups were analyzed using Student’s t-test. In cases of multiple-group testing, one-way analysis of variance and post hoc Bonferroni test were adopted. A two-tailed value of P < 0.05 was considered statistically significant.
References
Friedman, J. M. Obesity in the new millennium. Nature 404, 632–634 (2000).
Klein, S. et al. Waist circumference and cardiometabolic risk: a consensus statement from Shaping America’s Health: Association for Weight Management and Obesity Prevention; NAASO, The Obesity Society; the American Society for Nutrition; and the American Diabetes Association. The American journal of clinical nutrition 85, 1197–1202 (2007).
Wise, J. Obesity rates rise substantially worldwide. Bmj 348, g3582 (2014).
Rosen, E. D. & MacDougald, O. A. Adipocyte differentiation from the inside out. Nature reviews. Molecular cell biology 7, 885–896 (2006).
Locke, M., Feisst, V. & Dunbar, P. R. Concise review: human adipose-derived stem cells: separating promise from clinical need. Stem cells 29, 404–411 (2011).
Zuk, P. A. et al. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue engineering 7, 211–228 (2001).
Liang, W. C. et al. MiR-25 Suppresses 3T3-L1 Adipogenesis by Directly Targeting KLF4 and C/EBPalpha. Journal of cellular biochemistry 116, 2658–2666 (2015).
Zhang, X. M. et al. MicroRNA-29b promotes the adipogenic differentiation of human adipose tissue-derived stromal cells. Obesity 24, 1097–1105 (2016).
Cioffi, M. et al. MiR-93 Controls Adiposity via Inhibition of Sirt7 and Tbx3. Cell reports 12, 1594–1605 (2015).
Shi, C. et al. miR-148a is Associated with Obesity and Modulates Adipocyte Differentiation of Mesenchymal Stem Cells through Wnt Signaling. Scientific reports 5, 9930 (2015).
Pang, L. et al. miR-1275 inhibits adipogenesis via ELK1 and its expression decreases in obese subjects. Journal of molecular endocrinology 57, 33–43 (2016).
Ulitsky, I. & Bartel, D. P. lincRNAs: genomics, evolution, and mechanisms. Cell 154, 26–46 (2013).
Batista, P. J. & Chang, H. Y. Long noncoding RNAs: cellular address codes in development and disease. Cell 152, 1298–1307 (2013).
Sun, L. et al. Long noncoding RNAs regulate adipogenesis. Proceedings of the National Academy of Sciences of the United States of America 110, 3387–3392 (2013).
Li, M. et al. Long non-coding RNA ADNCR suppresses adipogenic differentiation by targeting miR-204. Biochimica et biophysica acta 1859, 871–882 (2016).
Xiao, T. et al. Long Noncoding RNA ADINR Regulates Adipogenesis by Transcriptionally Activating C/EBPalpha. Stem cell reports 5, 856–865 (2015).
Pang, W. J. et al. Knockdown of PU.1 AS lncRNA inhibits adipogenesis through enhancing PU.1 mRNA translation. Journal of cellular biochemistry 114, 2500–2512 (2013).
Cooper, D. R. et al. Long Non-Coding RNA NEAT1 Associates with SRp40 to Temporally Regulate PPARgamma2 Splicing during Adipogenesis in 3T3-L1 Cells. Genes 5, 1050–1063 (2014).
Augoff, K., McCue, B., Plow, E. F. & Sossey-Alaoui, K. miR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Molecular cancer 11, 5 (2012).
Yang, H. et al. Long noncoding RNA MIR31HG exhibits oncogenic property in pancreatic ductal adenocarcinoma and is negatively regulated by miR-193b. Oncogene 35, 3647–3657 (2016).
Nie, F. Q. et al. Decreased long noncoding RNA MIR31HG is correlated with poor prognosis and contributes to cell proliferation in gastric cancer. Tumour biology 37, 7693–7701 (2016).
Montes, M. et al. The lncRNA MIR31HG regulates p16(INK4A) expression to modulate senescence. Nature communications 6, 6967 (2015).
Jin, C. et al. Inhibition of lncRNA MIR31HG promotes osteogenic differentiation of human adipose-derived stem cells. Stem cells 34, 2707–2720 (2016).
Mercer, T. R. & Mattick, J. S. Structure and function of long noncoding RNAs in epigenetic regulation. Nature structural & molecular biology 20, 300–307 (2013).
Chen, S. Z. et al. The miR-181d-regulated metalloproteinase Adamts1 enzymatically impairs adipogenesis via ECM remodeling. Cell death and differentiation 23, 1778–1791 (2016).
Mikkelsen, T. S. et al. Comparative epigenomic analysis of murine and human adipogenesis. Cell 143, 156–169 (2010).
Zhang, Q. et al. Dynamic and distinct histone modifications modulate the expression of key adipogenesis regulatory genes. Cell cycle 11, 4310–4322 (2012).
Siersbaek, R., Nielsen, R. & Mandrup, S. Transcriptional networks and chromatin remodeling controlling adipogenesis. Trends in endocrinology and metabolism 23, 56–64 (2012).
Tontonoz, P., Hu, E. & Spiegelman, B. M. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell 79, 1147–1156 (1994).
Rosen, E. D., Walkey, C. J., Puigserver, P. & Spiegelman, B. M. Transcriptional regulation of adipogenesis. Genes & development 14, 1293–1307 (2000).
Lefterova, M. I. et al. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes & development 22, 2941–2952 (2008).
Coe, N. R. & Bernlohr, D. A. Physiological properties and functions of intracellular fatty acid-binding proteins. Biochimica et biophysica acta 1391, 287–306 (1998).
Hotamisligil, G. S. et al. Uncoupling of obesity from insulin resistance through a targeted mutation in aP2, the adipocyte fatty acid binding protein. Science 274, 1377–1379 (1996).
Uysal, K. T., Scheja, L., Wiesbrock, S. M., Bonner-Weir, S. & Hotamisligil, G. S. Improved glucose and lipid metabolism in genetically obese mice lacking aP2. Endocrinology 141, 3388–3396 (2000).
Furuhashi, M. et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature 447, 959–965 (2007).
Yap, K. L. et al. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Molecular cell 38, 662–674 (2010).
Tsai, M. C. et al. Long noncoding RNA as modular scaffold of histone modification complexes. Science 329, 689–693 (2010).
Rinn, J. L. et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 129, 1311–1323 (2007).
Wang, K. C. et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature 472, 120–124 (2011).
Huang, Y. et al. Long Non-coding RNA H19 Inhibits Adipocyte Differentiation of Bone Marrow Mesenchymal Stem Cells through Epigenetic Modulation of Histone Deacetylases. Scientific reports 6, 28897 (2016).
Jia, L. F. et al. Expression, regulation and roles of miR-26a and MEG3 in tongue squamous cell carcinoma. International journal of cancer 135, 2282–2293 (2014).
Acknowledgements
This study was supported by grants from the National Natural Science Foundation of China (81670957 and 81402235) and from the Peking University School of Stomatology Fund for Talented Young Investigators (PKUSS20140104).
Author information
Authors and Affiliations
Contributions
Y.H. performed the in vitro experiments, analyzed the data, prepared the figures and tables, and wrote the main manuscript text. C.J. performed experiments, including lentivirus transfection and in vivo assay. Y.Z. performed the cell proliferation experiments, assisted in data analysis. X.L. assisted in data collection and analysis. S.Z contributed to the ChIP assay. Y.Z. contributed to the Oil red O staining experiments. L.J. and W.L. conceived the experiments, supervised the work, and edited the manuscript. All authors approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Huang, Y., Jin, C., Zheng, Y. et al. Knockdown of lncRNA MIR31HG inhibits adipocyte differentiation of human adipose-derived stem cells via histone modification of FABP4 . Sci Rep 7, 8080 (2017). https://doi.org/10.1038/s41598-017-08131-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-08131-6
This article is cited by
-
The interaction between MALAT1 and TUG1 with dietary fatty acid quality indices on visceral adiposity index and body adiposity index
Scientific Reports (2024)
-
Stroma AReactive Invasion Front Areas (SARIFA) improves prognostic risk stratification of perioperative chemotherapy treated oesophagogastric cancer patients from the MAGIC and the ST03 trial
British Journal of Cancer (2024)
-
Stroma AReactive Invasion Front Areas (SARIFA) proves prognostic relevance in gastric carcinoma and is based on a tumor–adipocyte interaction indicating an altered immune response
Gastric Cancer (2024)
-
Pulling the trigger: Noncoding RNAs in white adipose tissue browning
Reviews in Endocrine and Metabolic Disorders (2024)
-
LncRNA MIR31HG fosters stemness malignant features of non-small cell lung cancer via H3K4me1- and H3K27Ace-mediated GLI2 expression
Oncogene (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
