
Abstract
It is widely accepted that modern pigs were domesticated independently at least twice, and Chinese native pigs are deemed as direct descendants of the first domesticated pigs in the corresponding domestication centers. By analyzing mitochondrial DNA sequences of an extensive sample set spanning 10,000 years, we find that the earliest pigs from the middle Yellow River region already carried the maternal lineages that are dominant in both younger archaeological populations and modern Chinese pigs. Our data set also supports early Neolithic pig utilization and a long-term in situ origin for northeastern Chinese pigs during 8,000–3,500 BP, suggesting a possibly independent domestication in northeast China. Additionally, we observe a genetic replacement in ancient northeast Chinese pigs since 3,500 BP. The results not only provide increasing evidence for pig origin in the middle Yellow River region but also depict an outline for the process of early pig domestication in northeast China.
Similar content being viewed by others


Introduction
It is widely accepted that pigs were domesticated independently in Near East and East Asia beginning ~10,000 years ago after Sus sp. emerged in Southeast Asia during the climatic fluctuations of the early Pliocene 5.3–3.5 My ago1,2,3. So far, at least six phylogeographically distinct wild boar lineages have been found to have contributed to the present domestic pig populations4. Over the past decade, regions in China, including the Mekong River basin5, the downstream region of the Yangtze River5, the upper stream region of the Yangtze River6, the Tibetan highlands7 and the lower region of the Yellow River8, 9 have been suggested as regions from which wild boar have contributed to the domestic pig gene pool and which may have represented independent centers for pig domestication.
Zoo-archaeological studies on the pig remains from the Early Neolithic assemblages in China back this idea of multiregional pig domestication in East Asia. The faunal assemblages of the ~9,000-y-old Zhenpiyan cave site10 in Guangxi province and the ~8,000-y-old Kuahuqiao site11 in Zhejiang Province both provide evidence for a commensal domestication phase12 in the Mekong River basin and the downstream region of Yangtze River. Furthermore, numerous pig remains excavated in the ~10,500-y-old Nanzhuangtou site13, the ~9,000-y-old Jiahu site14, and the ~7,500-y-old Cishan site15 as well as palaeogenetic studies8 presented the middle and lower branches of the Yellow River as one of the earliest regions for pig domestication. Extensive archaeological findings in these regions, such as broomcorn millet, foxtail millet, dog and chicken, are evidence for early origins of mixed agriculture16,17,18. Besides the archaeological findings, pigs seem to have had cultural significance for the communities in this region because two pig skeletons were buried with humans at Xinglongwa Site (8,200–7,400 BP), implying either that pigs may already have been domesticated in Northeast China around 8,000 years ago or that wild boar had cultural significance19, 20.
It has previously been shown that, at least with regard to maternal lineages, Chinese native pig breeds were direct descendants of the first domesticated pigs in the corresponding geographical regions8, 9. However, it is likely that roaming in a semi-managed status during the beginning of domestication allowed recurrent genetic exchange between domesticated pigs and indigenous wild boars. Thus, even during the beginning of domestication, genetic similarity between domestic pigs and wild boar from the same region may indicate introgression of wild boar haplotypes into the domestic gene pool rather than independent domestication events. Moreover, human migrations and cultural expansions repeatedly took place in East Asia during the past 10,000 years. Thus, human-mediated dispersal of and gene flow among domestic pig populations would not have been uncommon. Both for European and Near Eastern pigs, ancient DNA and dental geometric morphometric analyses have revealed nonlinear patterns of early domestication, dispersal of animals, genetic turnover and interaction among various geographic regions21,22,23.
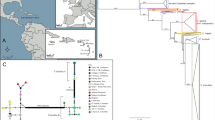
We have collected pig remains from 15 archaeological sites in China, including Jiahu and another two of the oldest archaeological sites in the middle Yellow River basin (Nanzhuangtou and Cishan), three sites in Northeast China (Xinglongwa, Wanfabozi and Dashanqian), and one site (Changning) in the upper region of the Yellow River. We investigated partial mitochondrial DNA control region (CR) sequences and a segment of the cytochrome b gene (Cytb) from both ancient and modern pigs, aiming to uncover the pattern of early maternal domestication and lineage dispersal in China. Locations and details for all samples can be found in Fig. 1 and Table S1.

Locations of samples. The orange dots depict all sites from which we retrieved ancient pig sequences. The inset shows the location of the region within China. The maps are modified from free map materials deposited in the public database of National Administration of Surveying, Mapping and Geoinformation (http://219.238.166.215/mcp/index.asp).
Results
Using loop-mediated PCR (L-PCR) followed by a specific singleplex PCR (for primers used see Table S2), we retrieved 49 Cytb fragments (Table S3) and 15 CR fragments (Table S4) of ancient pig samples from 15 archaeological sites. By combining our new results with previously reported sequences, we obtained a data set of 38 ancient pig CR sequences from China (Table S4) and combined Cytb/CR sequences from 37 samples, ranging geographically from northeast to northwest China and temporally from 10,000 BP to 2,500 BP (Table S1). We confirmed the Neolithic context of some of the early samples by determining the ages of the archaeological pig bones by direct radiocarbon dating of two bones from Cishan site, one bone from Nanzhuangtou site and one bone from Xinglongwa site. These samples yielded calibrated dates of ∼7,600, ∼7,800, ∼10,500 and ∼7,500 years, respectively (Tables S1 and S5), confirming the archaeologically determined ages.
We aligned the 37 combined sequences with 97 published homologous sequences of extant wild boar, native pig breeds and commercial pig breeds from all over the world (Table S6). Phylogenetic analyses resulted in two apparent clusters, which represent Asian and European origins, respectively (Fig. 2). All 10,000 BP to 2,500 BP Chinese pigs were found in the Asian clade (Fig. 2).

Bayesian consensus phylogenetic tree of the 37 combined (Cytb and control region) ancient pig sequences and 98 extant published homologous sequences. Ages (modern vs. ancient), status (wild vs. domestic) and geographical origin are depicted in different colours (see bottom left of the figure). The European and Asian sequence clusters are framed.
To investigate the source of the genetic diversity in Asian pigs in more detail, we compared ancient and modern Asian pigs based on the mitochondrial Cytb sequences alone. We aligned our 49 ancient sequences (Table S3) with 506 extant homologous sequences, which represent modern Asian pigs deposited in GenBank (Table S7). We found 20 SNPs among all Asian pig sequences, which constituted 21 haplotypes (Table S7). The 20 SNPs included 10 nonsynonymous substitutions, by which we classified Asian pigs into 11 groups defined by different Cytb amino acid sequences. The group AA1 contained 10 haplotypes representing 525 sequences (almost 95% of the sequences) and all geographically defined pig types from Asia (Fig. 3a and b), making this haplogroup the predominant haplogroup among Asian pigs. The groups AA2-AA11 contained the remaining 30 sequences, representing pigs from China, Japan, Korea and South East Asia Island, with one to maximally two AA substitutions to AA1 in this region of Cytb (Fig. 3a and b, Table S7) with various Grantham conservative scores24, 25.

Ancient Cytb gene analyses. Relationship of pig Cytb amino acid sequences. (a) Each haplogroup is represented by a circle, with the area of the circle proportional to the haplogroup’s frequency. Different colors indicate samples originating from different regions. The numbers alongside AA changes are the Grantham scores which help classify the conservation levels of AA changes. The number of asterisks highlights the level: non - considered conservative; one - moderately conservative; two - moderately radical and three - radical, respectively. (b) Alignment of AA changes among the 11 AA haplogroups and 21 DNA haplotypes. The AA haplogroups and DNA haplotypes containing ancient pigs are in red font.
The 38 ancient CR sequences obtained (Table S4) were aligned with 1,716 published homologous sequences from China, including 1,495 modern native domestic pigs and 221 wild boar (Table S8), representing 73 haplotypes (Table S8). Using the warthog (P. africanus) as outgroup, the consensus Bayesian tree resulted in three clades and a large polytomy representing the majority of the sequences (Fig. S1). The largest one (in red) of the three clades consisted of 8 haplotypes representing 12 Tibet Plateau pig samples and 1 haplotype representing 1 wild boar and 1 domestic pig each from southwest China. The other two clades (in green) were formed only by wild boars, in which one contained 2 haplotypes (representing 9 sequences) and the other one contained 3 haplotypes (25 sequences). The polytomy consisted of numerous branches, where three haplotypes were found only in ancient samples, 21 haplotypes were found only in wild boar, and 20 haplotypes only in modern domestic pigs, 6 of which were found only in Plateau pigs, while the remaining nine haplotypes were found in wild boars, ancient samples and modern domestic pigs (Table S8).
The 38 CR sequences obtained from ancient samples constitute 11 haplotypes (Table 1 and Fig. S2). Haplotypes H2 and H3 first appeared in Nanzhuangtou samples from the middle Yellow River valley in northern China at least 10,500 BP (~10,000 yrs for H2 from the indirectly dated dNZT1-3 sample and, ~10,500 yrs for H3 from the directly dated sample dNZT4). We also found haplotype H3 in sample dJH1 from Jiahu dated to ~9,000 BP. Haplotype H4 was first found in sample dJH2 also from Jiahu dating to at least ~8,500 BP. We also found it in sample dCS2 from Cishan (directly dated to ~7,800 yrs), and at Xinglongwa in Northeast China dating to 7,400–8,000 BP (sample dXLW1). The ancient sample dXLW2 from Xinglongwa, directly dated to ~7,500 BP, yielded haplotype H18, which also appeared in sample dWD2 from Wadian in the middle Yellow River area dating to ~4,000 BP. The ancient sample dCS1 from Cishan, directly dated to ~7,600 yrs, yielded haplotype H19 which also emerged in sample dWD3 from Wadian (~4,000 BP). We also found five rare haplotypes amongst our ancient samples. Two younger samples from Northeast China were identified as the rare haplotypes H31 (sample dWBP1 from Wanfabozi, 5,000–4,000 BP) and H70 (sample dDSQ2 from Dashanqian, 4,000–3,500 BP), while three ~4,000-y-old samples from the middle Yellow River region, dWD1, dGCJ1 and dGCJ3, yielded the rare haplotypes H71, H72 and H73, respectively. Except for those samples carrying rare haplotypes, ancient samples younger than 4,500 yrs carried haplotype H10 or haplotypes H2, H3 and H4 that were already found in the oldest samples (Table 1 and Fig. S2).
Together, the four haplotypes H2, H3, H4 and H10 accounted for 1,376 of the 1,754 investigated sequences (almost 80%), clearly representing the dominant haplotypes found in Chinese pigs (Table 1 and Fig. S3). Moreover, haplotypes that appeared before 7,000 BP in the middle Yellow River valley, including the dominant haplotypes H2, H3 and H4 as well as the less dominant haplotype H19, were repeatedly found in different historical sites, and represent a large part of the haplotypic diversity found in modern Chinese pigs (~55% of modern pigs carry these haplotypes; Fig. 4 and Table 1). In contrast, the fourth dominant haplotype, H10, which is found in ~25% of modern Chinese pigs, was detected only around 4,500 years ago in several sites across China within our data set (Fig. 4 and Table 1). Other contemporaneous haplotypes H70, H72 and H73 were generally detected almost exclusively in ancient pigs, except H70, which we found in one modern pig (Table 1). These three rare haplotypes are genetically close to the dominant haplotypes H2 or H10, separated by only one or two SNPs. A similar picture arises in Northeast China, where the ~4,000 y old haplotypes H31 and H71 were found, which are close to H2 and H10, respectively. While we did not find H71 in modern pigs, H31 interestingly appeared in two modern wild populations (Table S8). Moreover, the 8,000–7,400 years old remains in Northeast China carried not only the dominant haplotype H4 but also the less dominant haplotype H18. Even though H18 was never found in modern samples from Northeast China, it was detected in a younger sample (4,000 BP) from Wadian from the middle Yellow River region and at ~2.4% in modern pig populations in southern China (Fig. 4 and Table 1). Finally, for the ~4,000 year old samples from the upper region of the Yellow River and the middle region of the Yangtze River, we found that they shared similar haplotype compositions with ancient pigs in the middle Yellow River basin (Fig. 4). Overall, after their first appearance in the middle region of the Yellow River and Northeast China, most of the oldest haplotypes were inherited into younger times up to modern Chinese pigs (Fig. 4 and Table 1).

Temporal transition of mtDNA control region haplotypes of ancient Chinese pigs in different regions. The upper left map shows the location of the investigated region within China. From left to right and top to bottom: Time series of maps identifying the locations and haplotypes of ancient pig samples from which DNA sequences were obtained. Each symbol corresponds to a single sample. Different colours indicate different haplotypes (see lower right panel of the figure). The pie chart at the lower right shows the proportions of the different haplotypes in present day Chinese pigs. Maps are modified from free map materials deposited in the public database of National Administration of Surveying, Mapping and Geoinformation (http://219.238.166.215/mcp/index.asp).
Discussion
It is generally accepted that pigs were domesticated in at least two separate domestication centers, Europe and Asia26,27,28. In our data set of mitochondrial DNA sequences from ancient Chinese pigs, we found both considerable differences in haplotype composition over time and long-term continuity of some haplotypes.
The middle Yellow River basin has long been discussed as one center for early Chinese pig domestication based on both ancient DNA and morphometric evidence8, 14, 29. Already the earliest pig remains from this region revealed the haplotypes that represent the dominant maternal lineages in modern Chinese pigs (Table 1), supporting the hypothesis that the middle Yellow River basin indeed was one of the earliest centers for pig domestication from where domestic pigs spread to other regions in China. In a previous study, we demonstrated maternal lineage continuity in pigs from between ~9,000-y-old remains in the middle and ~4,000-y-old ones in the upper region of the Yellow River9. The present results from two samples from Changning in the upper region of the Yellow River further support this interpretation. Moreover, isotope data from Dadiwan in the upper Yellow River region provided evidence for a cultural replacement by central Chinese Neolithic cultures before 4,000 BP30, suggesting a possible reason why the ~4,000 BP pigs in the upper Yellow River region could have originated from progenitors from the middle Yellow River region. Similar to our previous study9, additional samples from Qinglongquan in the middle Yangtze River region also carry the dominant haplotypes that were already found in ~10,000 BP samples from the middle Yellow River drainage, suggesting a possible southward migration before (or around) ~4,000 year ago.
Due to close relationships between humans and pigs and evidence of early millet farming at Xinglongwa, Northeast China has been proposed as another region of early pig domestication20. However, only pig remains younger than 8,000 BP are assumed to represent domesticated ones19. In the present study, Xinglongwa specimens carry the dominant modern haplotype H4 and the less common haplotype H18, respectively, demonstrating maternal continuity between the earliest archaeological pig remains in this region and modern Chinese domestic pigs (Fig. 4 and Table 1). The haplotype H4 also appears in Jiahu and Cishan around the same time (Fig. 4), suggesting either early population migrations, trade of early domestic pigs or parallel domestication across the North China Plain and Northeast China. Suidae remains in Xinglongwa were found in all three cultural phases (8,200–8,000 BP, 8,000–7,400 BP and 7,400–7,000 BP)19, suggesting continuous utilization of pigs during the early Neolithic in this region. The younger haplotypes H31 and H71 discovered in Northeast China (Wanfabozi and Dashanqian) have so far not been found in the middle Yellow River region (Fig. 4 and Table 1), implying also long-term in situ continuity after initial pig domestication, or possibly, introgression of wild boar haplotypes into imported, early domestic pig herds, in Northeast China.
In addition to long-term continuity, we also observed genetic turnovers in ancient Northeast Chinese pigs. One example is haplotype H18, which we found in a ~7,500 year old sample from Xinglongwa, but which seems to have disappeared afterwards in Northeast China. However, we found it in younger samples from the middle Yellow River region ~4,000 BP (Fig. 4 and Table 1) and also in modern pigs from southern China. In contrast, the rare haplotype H71, which we found in a single ~3,500 year old sample from Northeast China has not yet been found in modern pigs (Table 1), indicating that genetic drift also played a role in the history of domestic pigs during the last 3,500 BP. This view is further supported by haplotypes H72 and H73, which are both from the ~4,000 BP Guchengzhai site in the middle Yellow River Region and have not been found in any younger sites or modern pigs so far.
The last pattern was found for haplotype H31 (Table S7), which we discovered only in one 4,000–5,000 year old sample from Northeast China and in two wild boar from southern China. In contrast to past pig populations, modern pigs in Northeast China show a similar mitochondrial haplotype composition as other regions (Fig. S3 and Table S7), suggesting extensive homogenization of pig populations over time, accompanied by a loss of at least some of the original, regional maternal lineages.
Conclusion
Our results support early independent domestication and long-term genetic continuity of pigs both in Europe and East Asia. The earliest pigs from the middle Yellow River region already carried the maternal lineages that are dominant in both younger archaeological populations and modern Chinese pigs. Moreover, there is strong evidence for early Neolithic pig utilization and possibly independent domestication, or alternatively, incorporation of local wild boar into introduced domestic pig populations in northeast China. However, this in situ origin for northeastern Chinese pigs ended around 3,500 BP due to genetic replacement, likely by pigs originating from the middle Yellow River region.
Materials and Methods
Sample Information
We used ninety-three ancient pig specimens (bones or teeth) for DNA analyses, which originate from 15 archaeological sites in northern and central China (Fig. 1), including 65 specimens from nine sites in the lower and middle area of the Yellow River drainage basin (Nanzhuangtou and Cishan sites in Hebei province; Jiahu, Nanwa, Guchengzhai, Wadian and Wangchenggang sites in Henan province; Taosi and Gaohong sites in Shanxi province), 6 specimens from two sites located in the upper region of the Yellow River (Lajia and Changning sites in Qinghai province), 12 specimens from one site (Qinglongquan site in Hubei province) located in the middle region of the Yangtze River, and 10 specimens from three sites in Northeast China (Xinglongwa and Dashanqian sites in eastern Inner Mongolia and Wangfabozi site in Jilin province). Dates of these archaeological sites ranged from 10,600 to 2,500 years before present. Details regarding the archaeological sites and the contexts from which the specimens were recovered can be found in Table S1.
Sample Preparation and Ancient DNA Extraction
All pre-PCR work was conducted in the Ancient DNA Laboratory at China Agricultural University. Samples were prepared by cautiously cleaning the adhering soils and other external contaminations using abrasive paper, and then washing them with 5% (vol/vol) sodium hypochlorite solution followed by double-distilled water and drying under UV-irradiation. After that, bones and teeth were ground to fine-grained powder.
DNA was extracted by using the QIAamp DNA Investigator kit (Qiagen) and Amicon Ultra-4 filters (Millipore). DNA extraction followed the QIAamp DNA Investigator handbook for purification of total DNA from bones or teeth. Amicon Ultra-4 (Millipore, 10 K) filters were used to concentrate ancient DNA to a final volume of ∼50 μL. Several mock extractions were carried out alongside the samples in the same manner to monitor for contamination.
Amplification and Sequencing of Ancient DNA
We used loop-mediated PCR (L-PCR) followed by a specific singleplex PCR amplification and Sanger sequencing to obtain the targeted ancient pig DNA sequences. As previously described18, L-PCR is designed to efficiently enrich the target copy number using loop-mediated isothermal amplification primer sets; subsequent singleplex PCR then allows generating a specific amplicon that can then be sequenced.
Primers were designed according to the published mitochondrial sequence of S. scrofa (NC_000845). All L-PCR primers were designed by online loop-mediated isothermal amplification primer designing software (primerexplorer.jp/e/). The primer set of the mitochondrial Cytb gene was newly designed for this study. The L-PCR primer pair of the mitochondrial control region was designed based on the control region PCR primer pair described by Larson et al.8. Amplicons of the Cytb gene and the control region were 101 bp and 138 bp in size, respectively, and contained polymorphic sites which were used to define haplogroups or haplotypes. Detailed primer sequences are available in Table S2.
L-PCR and the specific singleplex PCR amplification were setup as previously described18. Several blank controls were performed in all PCR assays. L-PCR used the following cycling conditions: 37 °C for 10 min, 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 65 °C for 40 s, 72 °C for 30 s, and a final extension of 10 min at 72 °C. Secondary PCR used the following cycling conditions: 37 °C for 10 min, 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 61 °C for 30 s, 72 °C for 30 s, and a final extension of 10 min at 72 °C. Amplifications of the extraction blank controls and PCR blank controls were performed in all experiments to monitor contaminations. Amplification success was controlled by electrophoresis on a 2% agarose gel. Subsequently, PCR products were purified using the QIAquick PCR purification kit (Qiagen). Sequencing was carried out on an ABI 3730XL automated DNA sequencer (Applied Biosystems) using the ABI Prism Big Dye Terminator v3.1 Cycle Sequencing kit.
Independent Replication Experiments
For all ancient DNA samples, we performed several replication experiments in the Ancient DNA Laboratory at China Agricultural University. Moreover, some of the positive samples were sent to the Ancient DNA Laboratory at the Research Center for Chinese Frontier Archeology at Jilin University for independent replication experiments (Table S1). DNA extraction, amplification and sequencing were carried out using the same methods and conditions in both laboratories.
Radiocarbon Dating Analysis
All samples used in the present study were from well-defined archaeological contexts. Nevertheless, four ancient bones from the three key archaeological sites of Nanzhuangtou (1 bone), Cishan (2 bones) and Xinglongwa sites (1 bone), which yielded DNA sequences were sent for radiocarbon dating using direct accelerator mass spectrometry at Beta Analytic Inc. (USA) to provide further support of their ages (Table S5).
Data Analyses
The reconstructed ancient DNA sequences were aligned with extant published homologous sequences from wild boars, local domestic pigs and commercial breeds from all over the world (Table S6). The ancient Cytb gene sequences (Table S3) and homologous sequences of Asian S. scrofa were aligned and investigated considering both DNA and amino acid (AA) phylogenetic relationship (Table S7). Particularly, to determine the origin and dispersal pattern of the dominant haplotypes in Chinese pig breeds, the ancient control region sequences (Table S4) were utilized in comparison with published sequences of ancient and modern Chinese domestic pigs and wild boars (Table S8). Sequences were aligned using MUSCLE31 or the online tool MAFFT32, and edited using MEGA 633; then the online tool FaBox (users-birc.au.dk/biopv/php/fabox/) was used to classify haplotypes34. MrBayes 3.2.335 was used for phylogenetic analysis with model parameters identified by jModelTest 2.1.136. The length of the MCMC was set to 10,000,000. Parameter estimates and consensus trees resulting from 10 MrBayes runs were recorded and compared. The best supported phylogenetic consensus tree was summarized with discarding the first 25% as burn-in. The tree was depicted using the software FigTree v1.4.2 (http://tree.bio.ed.ac.uk/software/figtree/). Median-joining networks37 were reconstructed using the software Network 4.6.1.0 (www.fluxus-engineering.com/index.htm). The aligned DNA sequence file format conversions were performed using BioEdit 7.038 and Forcon 1.039.
References
Groenen, M. A. et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 491, 393–398 (2012).
Bosse, M. et al. Genomic analysis reveals selection for Asian genes in European pigs following human-mediated introgression. Nat. Commun. 5, 4392 (2014).
Frantz, L. A. et al. Genome sequencing reveals fine scale diversification and reticulation history during speciation in Sus. Genome Biol. 14, R107 (2013).
Larson, G. et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science 307, 1618–1621 (2005).
Wu, G. S. et al. Population phylogenomic analysis of mitochondrial DNA in wild boars and domestic pigs revealed multiple domestication events in East Asia. Genome Biol. 8, R245 (2007).
Jin, L. et al. Mitochondrial DNA evidence indicates the local origin of domestic pigs in the upstream region of the Yangtze River. PloS one 7, e51649 (2012).
Yang, S. et al. The local origin of the Tibetan pig and additional insights into the origin of Asian pigs. PloS one 6, e28215 (2011).
Larson, G. et al. Patterns of East Asian pig domestication, migration, and turnover revealed by modern and ancient DNA. Proc. Natl Acad. Sci USA 107, 7686–7691 (2010).
Wang, Z., Xiang, H., Yuan, J., Luo, Y. & Zhao, X. Exploring the origin of domesticated pigs in the Yellow River area using information from ancient DNA. Chin. Sci. Bull. 57, 1011–1018 (2012).
Li, Y. H. & Han, D. F. The ruins fauna from Zhenpiyan at Guilin, Guangxi. Vertebrata PalAsiatica 16, 244–254 (1978).
Zhejiang Provincial Institute of Cultural Relic and Archaeology & Xiaoshan Museum. Kuahuqiao. (Cultural Relic Publishing House, 2004).
Zeder, M. A. The Domestication of Animals. J. Anthropol. Res. 68, 161–190 (2012).
Xu, H. S., Jin, J. G. & Yang, Y. H. Test excavation to Nanzhuangtou site in Xushui County, Hebei Province. Archaeology 11, 961–970 (1992).
Luo, Y. & Zhang, J. Restudy of the Pigs’ Bones from the Jiahu Site in Wuyang County, Henan. Archaeology 1, 90–96 (2008).
Sun, D. H., Liu, Y. & Chen, G. T. The Cishan site in Wu’ an, Hebei Province. Acta Archaeologica Sinica 3, 303–338 (1981).
Lu, H. et al. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago. Proc. Natl Acad. Sci USA 106, 7367–7372 (2009).
Yang, X. et al. Early millet use in northern China. Proc. Natl Acad. Sci USA 109, 3726–3730 (2012).
Xiang, H. et al. Early Holocene chicken domestication in northern China. Proc. Natl Acad. Sci USA 111, 17564–17569 (2014).
Yang, H. & Liu, G. X. Preliminary report on 1992 season excavation at the Xinglongwa site in Aohan, Inner Mongolia. Archaeology 1, 1–26 (1997).
Yuan, J., Han, J. & Blench, R. Livestock in ancient China: An archaeozoological perspective. in Past Human Migrations in East Asia: Matching Archaeology, Linguistics and Genetics (eds A. Sanchez-Mazas et al.) (Routledge Taylor & Francis Group, 2008).
Ottoni, C. et al. Pig domestication and human-mediated dispersal in western Eurasia revealed through ancient DNA and geometric morphometrics. Mol. Biol. Evol. 30, 824–832 (2012).
Krause-Kyora, B. et al. Use of domesticated pigs by Mesolithic hunter-gatherers in northwestern Europe. Nat. Commun. 4, 2348 (2013).
Larson, G. et al. Ancient DNA, pig domestication, and the spread of the Neolithic into Europe. Proc. Natl Acad. Sci USA 104, 15276–15281 (2007).
Grantham, R. Amino-Acid Difference Formula to Help Explain Protein Evolution. Science 185, 862–864 (1974).
Li, W. H., Wu, C. I. & Luo, C. C. Nonrandomness of Point Mutation as Reflected in Nucleotide Substitutions in Pseudogenes and Its Evolutionary Implications. J. Mol. Evol. 21, 58–71 (1984).
Giuffra, E. et al. The origin of the domestic pig: independent domestication and subsequent introgression. Genetics 154, 1785–1791 (2000).
Manunza, A. et al. A High Throughput Genotyping Approach Reveals Distinctive Autosomal Genetic Signatures for European and Near Eastern Wild Boar. PloS one 8, e55891 (2013).
Li, M. et al. Comprehensive variation discovery and recovery of missing sequence in the pig genome using multiple de novo assemblies. Genome Res. 17, 865–874 (2016).
Cucchi, T., Hulme-Beaman, A., Yuan, J. & Dobney, K. Early Neolithic pig domestication at Jiahu, Henan Province, China: clues from molar shape analyses using geometric morphometric approaches. J. Archaeol. Sci. 38, 11–22 (2011).
Barton, L. et al. Agricultural origins and the isotopic identity of domestication in northern China. Proc. Natl Acad. Sci USA 106, 5523–5528 (2009).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797 (2004).
Katoh, K. & Standley, D. M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780 (2013).
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729 (2013).
Villesen, P. FaBox: an online toolbox for FASTA sequences. Mol. Ecol. Notes 7, 965–968 (2007).
Ronquist, F. et al. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542 (2012).
Darriba, D., Taboada, G. L., Doallo, R. & Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9, 772–772 (2012).
Bandelt, H. J., Forster, P. & Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 16, 37–48 (1999).
Hall, T. A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. in Nucleic Acids Symposium Series. 95–98 (1999).
Raes, J. & Van de Peer, Y. ForCon: a software tool for the conversion of sequence alignments. EMBnet J. 6, 10–12 (1999).
Acknowledgements
We express great thanks to Prof. Jing Yuan (Institute of Archaeology, Chinese Academy of Social Science) for generously helping ancient sample collection, Prof. Dengyun Qiao (Handan Municipal Institute of Cultural Relics and Archaeology) and Associated Prof. Xue Ling (Northwest University) for discussions and comments, Dr. Jikun Wang and Dr. Tao Yin (China Agricultural University) for experimental assistance, and Dr. Ye Zhang (Jilin University) for replication experiments. This work was funded by the National Natural Science Foundation of China (31672379), the National Key Basic Research Program of China (2014CB138500) and the China Postdoctoral Science Foundation (2015M581201).
Author information
Authors and Affiliations
Contributions
X.Z. designed research; H.X., D.C., R.L., L.L., H.Z., X.C., W.D. and X.W. performed research; J.G., D.C., Y.L. and B.Y. contributed new reagents/analytic tools; H.X., L.L., M.H. and X.Z. analyzed data; and H.X., M.H., and X.Z. interpreted the data and wrote the paper.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xiang, H., Gao, J., Cai, D. et al. Origin and dispersal of early domestic pigs in northern China. Sci Rep 7, 5602 (2017). https://doi.org/10.1038/s41598-017-06056-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-06056-8
This article is cited by
-
Population genetics reveals new introgression in the nucleus herd of min pigs
Genes & Genomics (2024)
-
Ancient genomes reveal the genetic inheritance and recent introgression in Chinese indigenous pigs
Science China Life Sciences (2022)
-
SINE jumping contributes to large-scale polymorphisms in the pig genomes
Mobile DNA (2021)
-
The Archaeology of Pig Domestication in Eurasia
Journal of Archaeological Research (2020)
-
Comprehensive analysis of genetic and evolutionary features of the hepatitis E virus
BMC Genomics (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
