
Abstract
At the global level, maize is the third most important crop on the basis of harvested area. Given its importance, an assessment of the variation in regional climatic suitability under climate change is critical. CliMond 10′ data were used to model the potential current and future climate distribution of maize at the global level using the CLIMEX distribution model with climate data from two general circulation models, CSIRO-Mk3.0 and MIROC-H, assuming an A2 emissions scenario for 2050 and 2100. The change in area under future climate was analysed at continental level and for major maize-producing countries of the world. Regions between the tropics of Cancer and Capricorn indicate the highest loss of climatic suitability, contrary to poleward regions that exhibit an increase of suitability. South America shows the highest loss of climatic suitability, followed by Africa and Oceania. Asia, Europe and North America exhibit an increase in climatic suitability. This study indicates that globally, large areas that are currently suitable for maize cultivation will suffer from heat and dry stresses that may constrain production. For the first time, a model was applied worldwide, allowing for a better understanding of areas that are suitable and that may remain suitable for maize.
Similar content being viewed by others

Introduction
Elevated atmospheric CO2 concentrations, global warming and extreme weather events will impact food production, altering the current level of suitability of regions for specific crops. Changes in rainfall patterns and increases in temperature and carbon dioxide levels are likely to have major implications for agricultural productivity, with positive impacts in some regions and negative impacts in others1,2,3. Elevated CO2 can improve photosynthetic efficiency, thus increasing the yield of C3 crops and decreasing water consumption through decreases in stomatal conductance in C3 and C4 crops4. Conversely, variations in temperature, precipitation and ozone concentrations may affect plant growth and development through increases in abiotic stress5,6,7,8. Such changes will have important impacts in the quantity and quality of agricultural production, in terms of food security and the welfare of a growing global population9.
Maize (Zea mays L.) is a major food source for the world and is a high-yield commodity crop, with an average harvested area of 157 million hectares and production of 781 mega tonnes from 2000 to 2014; it is a vital source of food security in many developing countries in Latin America and Sub-Saharan Africa10, 11. Furthermore, it serves as forage for the production of biogas12. Maize originated in the Mexican Highlands and spread around the world after the colonization of America. Mexico remains one of the main producers, with an average yearly production of 14 mega tonnes from 1961 to 2014, ranked fourth in the world13,14,15,16. Maize can be produced in an extended range of conditions, from 0 to over 3800 m.a.s.l., and under precipitation levels from 200 mm to 2000 mm13, 17,18,19. Though a variety of abiotic (soil, climate) and biotic (diseases, plagues) stresses affect maize, its main constraints are currently climatic factors and physical characteristics related to soil fertility12, 20.
To date, several studies have addressed the possible impacts of climate change on maize, mostly at the regional level and focusing on changes in productivity. Some studies have indicated that temperature increases have a negative effect on maize yield, whereas CO2 increases could be beneficial for changes in water availability. However, the level of uncertainty in the CO2 results has been consistently high in all research studies11, 21,22,23. This crop is extremely susceptible to drought during the flowering stage, during which the quality of the seed is reduced6. Maize drought stress could result in yield losses of nearly 50% in southern Africa24. In general, studies have reported a negative impact on maize production that is attributable to increasing temperatures and reduced precipitation9. Several institutions around the world have released maize varieties resistant to drought or heat stress to reduce vulnerability6, 24.
To assess the potential changes in global maize distribution due to climate change, an appropriate modelling technique should be applied. There are close to one hundred mechanistic and crop niche models that can simulate the potential consequences of climate change on crop production and species distributions. These models differ in their input parameters, protocols and methods11, 25. Some widely used distribution models include the following: EcoCrop, which is a mechanistic model that integrates the FAO-EcoCrop database and uses temperature, rainfall and length of the growing season as inputs26, 27; MaxEnt, which is an empirical approach that models the potential distributions of species based on presence information of the species of interest28, 29; and CLIMEX, which is a hybrid statistical-mechanistic model that is used to estimate the potential abundance and geographic distribution of an organism using climatic data and biological parameters30. CLIMEX has been widely used to model the suitability of a variety of organisms, from weeds to insects, at global and regional levels, providing important insights into the ecology of a species31,32,33,34. An important insight available in CLIMEX is the daily or weekly species’ response to climate variables. Furthermore, the model allows us to explore abiotic constraints, such as heat, cold, dry and moisture stresses34.
The objectives of this research were as follows: (i) to employ CLIMEX as a mechanistic species distribution model to assess potential changes in the global distribution of agricultural land for maize cultivation based on shifts in climatic suitability for current versus two time periods, 2050 and 2100 according to projections from two general circulation models (GCMs) CSIRO Mk3.0 and MIROC-H and assuming the A2 emissions scenario; (ii) to identify current maize cultivation regions that are likely to be severely impacted as a consequence of climate change and to identify future stresses; and (iii) to perform a sensitivity analysis to quantify species response to temperature, soil moisture and cold stress changes and to identify the parameters of functional importance to provide a greater understanding of the climatic factors that most impact species distribution.
Materials and Methods
CLIMEX description
CLIMEX exemplifies an eco-physiological growth-modelling approach, forecasting shifts in the abundance and distributions of species. The model employs data that are based on climatic information and biological parameters. The locations of climatic suitability are obtained from the species-response functions in CLIMEX, and these functions are based on ecological studies that provide a foundation for the successful modelling of potential species distribution. CLIMEX models the mechanisms affecting species and matches the geographic occurrences with meteorological data. The required monthly climatic data variables include average minimum temperature (Tmin), average maximum temperature (Tmax), average precipitation (Ptotal), and relative humidity at 09:00 and 15:00 hours (RH09:00 and RH15:00) for the specific research locations. The required biological parameters include minimal, maximal and optimal temperatures, as well as soil moisture data for the species. CLIMEX is based on the assumption that a population experiences two seasons. A favourable season produces the temperature and moisture requirements for growth, as described by the annual growth index (GIA) that is calculated from the temperature (TI) and moisture indices (MI). An unfavourable season supports no population growth and is represented by four stress indices (cold (CS), dry (DS), hot (HS) and wet (WS)) and their interactions. An annual index of climatic suitability was obtained with the weekly integration of the growth (favourable season) and stress (unfavourable season) indices. This integrated index describes the level of climatic suitability and is known as the ecoclimatic index (EI), which is calculated as follows (E.g. 1):
The EI range is from 0 to 100. An EI of 0 describes an unsuitable region for the species to persist and an EI of 100 represents a region with ideal conditions for the species. Such ideal conditions could only exist in a greenhouse or laboratory setting. In CLIMEX, the parameters that describe the response of a species to climate are deduced from its geographic range and phenological observations. Later, the inferred parameters are used to project the potential range in new areas or under new climate scenarios30, 35, 36. For a detailed description of the mechanisms of the CLIMEX model, see Sutherst et al., (2007) and Kriticos et al., (2015). In this study, CLIMEX version 4 was used. The following categories were used for the EI: EI of 0 represents unsuitable regions, an EI from 1 to 10 represents marginal regions, an EI from 10 to 20 indicates regions where large populations can persist (medium suitability) and an EI with values greater than 20 represents a highly favourable region for the species (optimal suitability)30, 35, 36.
Current maize distribution, climatology and climate change scenarios
Current maize distribution data were gathered from the Global Biodiversity Information Facility37 and literature resources11, 17, 38. The data were verified for biological reasonability; furthermore, duplicate records and those without geographic coordinates were eliminated from the database. Nearly 15,000 records were gathered and used for parameter fitting, with more than 50% of the records representing Mexico. These records geographically represent the current known global distribution of maize (Fig. 1). The use of native and planted (agricultural) distribution records to adjust CLIMEX parameters could generate a better model that more accurately reproduces the potential distribution of the species of study, allowing for the expansion of the fundamental niche36, 39, 40. Thus, both native and planted distributions were used to fit the CLIMEX parameters.

(a) The current global distribution of maize (Zea mays L.). (b) The ecoclimatic index (EI) for the current climate scenario of maize (Zea mays L.). ArcMap 10.2 (http://desktop.arcgis.com/en/arcmap).
CliMond provides 10′ (18.55 km) and 30′ (55 km) global high-resolution data for bioclimatic modelling for CLIMEX and Bioclim. CliMond data are based on WorldClim 10′ (minimum and maximum monthly temperature, monthly total precipitation) and CRU CL2·0 10′ (mean relative humidity). For this study, the climatology of 10′ gridded resolution data was used to model the potential current and future distribution of maize. The data were downloaded in CLIMEX format, containing average monthly maximum and minimum temperatures, average monthly precipitation and relative humidity at 9:00 hours and 15:00 hours41. The climate variables were averaged first, after which the species distribution was determined. Historical suitability was modelled with CliMond baseline data, averaging a period from 1961–199041. The future potential distribution was modelled using A2 SRES (Special Report on Emission Scenarios) scenario (business as usual) for 2050 and 2100 based on two GCMs: CSIRO Mk3·0 from Australia42 and MIROC-H, developed in Tokyo, Japan43. The CLIMEX CliMond climatology data are based on CSIRO and MIROC because these two GCMs have the climate data required for CLIMEX model and have superior performance in areas with a range of climates compared with other GCMs44. Data for both models is available on the CliMond website41. No other scenarios of the SRES family were included in this investigation, as the A2 SRES scenario incorporates actual CO2 emissions levels and population growth trends45, 46. The projected global average surface warming by the end of the present century for the A2 scenario is 3.4 °C, with a likely range from 2–5.4 °C47, global carbon dioxide emissions are estimated at approximately 30 GtC/yr.48 and carbon dioxide concentrations of 846 ppm are expected by 210049. A new report from the IPCC, the AR5 (Fifth Assessment Report) describes four greenhouse gas trajectories, as representative concentration pathways (RCPs) to replace the SRES scenarios49. The best equivalent of A2 is the RCP 8.5, representing a higher greenhouse gas emissions scenario50. The temperature increase in the period from 2090–2099 is relative to the pre-industrial era for A2 of approximately 6 °C, and the RCP 8.5 is 7 °C51. Furthermore, the CO2 concentrations by the end of the century for RCP 8.5 are 936 ppm, just above A2. The radiative forcing (W/m2) by the end of the century is relatively similar for A2 and RCP 8.550.
Fitting CLIMEX parameters
Distribution data for India, China and neighbouring countries was not used for parameter fitting, but was reserved for model validation. The reasons for the choices of parameter values are described in detail below. Each parameter was manually and iteratively adjusted until an acceptable visual level of agreement was obtained between the potential and current distribution of maize. The stress indices were first adjusted to maintain the population within the observed distribution limits and to model the core distribution. After adjusting the stresses, the temperature and moisture indices were fixed. All of the CLIMEX parameters were compared with experimental biological data to ensure their validity (Table 1).
Stress parameters
Cold stress
The cold stress temperature threshold (TTCS) sets the extreme low temperatures, below which the species cannot survive. TTCS is a weekly average of minimum temperatures that are accumulated at a specific rate, known as the cold stress temperature rate (THCS). Maize does not tolerate low temperatures and dies just below the freezing point5, 13, 52, 53. Low temperatures affect germination, emergence and vegetative growth. The intensity of the damage will depend on the air temperature and duration of the stress exposure. Maize plants are sensitive to cold temperatures, and frost is detrimental at all stages of plant development, except as dry seed (0 to 6 °C); young plants can be killed at 1 °C53, 54 or from the severe cold stress that occurs at 0 °C5, 55. A group of cold-induced maize genes (ZmCOI) that may affect abiotic stresses was isolated from maize exposed to 6 °C56. A review by Garcia and Lopez7 found that maize can tolerate temperatures as low as −2 and −3.5 °C, but they did not mention the acceptable duration of low temperatures. Thus, the TTCS was adjusted to 7 °C with a THCS of −0.00007 week−1, to allow for the existence of the coldest current locations, such as Norway, Sweden, Finland and northeastern China, but to avoid survival scenarios in Northern Russia.
Heat stress
Extreme high temperatures also terminate survival. This factor is modelled through a weekly heat stress temperature threshold (TTHS) and its accumulation rate (THHS). Maize is a thermophilic plant species that can tolerate high temperatures10, 13, 53. An extended literature review indicated that extremely high temperatures can cause sterility and reduce yield, with 46 °C being the lethal temperature for maize5, 57. Maize can tolerate temperatures below 45 °C, if not, drought stress occurs13, 54. Temperatures above 40 °C stop crop development, and no further heat units are accumulated for crop development57. Thus, TTHS was adjusted to 40 °C, and the THHS was set to 0.01 week−1.
Dry stress
This stress starts to accumulate when the conditions are too dry for the species. The weekly dry stress threshold (SMDS) accumulates at a given rate (HDS). Maize is susceptible to drought, especially during flowering, tasselling, silking and pollination, as well as during the grain-filling stage during which a lack of water can cause losses of nearly 90%, with little or no grain yield6, 12, 13, 57. Water shortages can restrict cell division and growth in maize53. Doorenbos and Kassam (1979) mentioned that with sufficient water availability, maize can tolerate dry atmospheric conditions and, during the vegetative and ripening periods, may be relatively tolerant of water deficits13, 54. Since maize can tolerate water shortages, the SMDS was set at 0.1 (the permanent wilting point), at a stress accumulation rate (HDS) of −0.009 week−1.
Wet stress
Extreme wet conditions can produce wet stress. This stress accumulates weekly once the soil moisture has passed a wet stress threshold (SMWS) at a given rate (HWS). Waterlogging may reduce growth, photosynthesis and cause a loss of biomass production or even death. Older maize plants are more tolerant to waterlogging than younger ones6, 58. The phenological stages that are more susceptible to waterlogging precede those of tasselling and flowering. Damage to the roots, due to the accumulation of toxic bioproducts, affect yields6, 13, 54. Maize prefers aerated and well-drained soils13, 54, although some varieties can produce adventitious roots and tolerate waterlogging20. Therefore, a SMWS of 1.3 was used with an accumulation rate (HWS) of 0.001 week−1.
Growth-related parameters
Temperature index
This weekly index describes the response of the crop to the daily temperature cycle, ranging between a lower temperature threshold (DV0) and an upper temperature threshold (DV3); optimal temperatures occur between the lower optimum temperature (DV1) and the upper optimum temperature (DV2). The most common base temperature for maize growth is 10 °C5, 7, 10, 53, 59,60,61. Thus, the DV0 was set at 10 °C. Maize requires high optimal temperatures for germination and growth53. The influence of temperature on the germination and elongation of maize radicles has been tested62. The elongation rate peaked at 30 °C and stopped at the extremities of 9 °C and 40 °C62. At 20 °C, early and medium grain varieties take between 80 and 140 days to mature. Optimal temperatures for germination are 18 to 20 °C13, 54. The optimal range during daylight is from 25 to 33 °C, and at night it is 17 to 23 °C. The optimal temperatures for the whole crop season are between 20 and 22 °C63. The DV1 was set at 18 °C and DV2 was set at 30 °C as a compromise between the varied values in the literature5, 57, 59,60,61, 64. Research has established temperatures ranging from 36 to 40.2 °C as maximal for growth development in maize5. Temperatures higher than 35 °C reduce dry matter accumulation during grain filling11, 63, 65. Thus DV3 was set at 35 °C.
Moisture index
The hydrological model integrated in CLIMEX represents the effects of rainfall and evaporation within the species, termed the ‘moisture index’36. Maize is most susceptible to water stress at flowering66. A study to determine the sensitivity of maize to water stress found that the threshold of the soil moisture stress index was between 0.20 and 0.3067. The limiting low soil moisture (SM0) was set to 0.1 to represent the permanent wilting point68, a value that is also consistent with the dry stress threshold (SMDS). The lower (SM1) and upper (SM2) soil moisture values were adjusted to 0.7 and 0.9, respectively. The limiting high soil moisture (SM3) was set at 1.3 to maintain consistency with the wet stress threshold (SMWS). Excessive moisture is a main constraint in regions such as Southeast Asia and India20. These values provided the best fit for current maize distribution. The CLIMEX parameters are summarized in Table 1.
Once the best fit was obtained, the final model was run for historical and future scenarios and maps were generated. Maps for the four stress indices were also obtained for current and future scenarios.
Parameter sensitivity of CLIMEX values
The sensitivity analysis determines how the accuracy of the parameters affect the model output when applied on an individual basis. A parameter sensitivity analysis runs the model repeatedly, altering individual parameters successively to exceed their fitted values. This analysis was carried out with the global CliMond database. Variables with a greater effect on the model output are described as ‘sensitive’, whereas those that have no impact on the model output are termed ‘insensitive’34.
Methods of validation
Occurrence data from India, China and neighbouring countries was not used in the fitting process, but was reserved for the validation process. Once the parameter fitting was satisfactory, the validation area was verified to check the performance of the model in this region. Furthermore, a cross validation that is incorporated in CLIMEX was performed, using two maps of current maize distribution based on productivity69, 70. The agreement between the modelled historical potential distribution and the seasonal phenology of the modelled species in different areas provided the cross validation for the model34.
Estimating land areas for various suitability classes
The Food and Agriculture Organization (FAO) has an agricultural statistical database that is available for download. Information on the average global maize yields from 1983 to 2013 was obtained from this database71, which was also used to identify the major maize-producing countries.
Category levels of suitability (marginal, medium, and optimal) and unsuitable areas, as reflected by the EI values, were determined according to continent, and for each of the five major maize producing countries. The raster images were re-projected using Behrmann projection to obtain the estimated sizes of the regions for the spatial analysis41.
Results
Validation methods, parameter sensitivity
The modelled distribution (Fig. 1b) shows an acceptable match with the current known distribution of maize (Fig. 1a). Approximately 96% of the occurrence records are classified in the marginal to optimal categories, while 88% of the occurrence records for the validation area (India, China and neighbouring countries) are located within the modelled marginal to optimal areas. The global distribution of You et al., (2014) and Leff et al., (2004), used for cross validation, indicates broad agreement with our maize model under historical conditions. These validations confirm the optimal performance of the present model and the validity of the parameter values selected.
The parameter sensitivity indicates that a limiting low moisture (SM0) of 5.3% and cold stress temperature rate (TCHS) of 3.5% impact are the most sensitive parameters in the modelled potential range (Table S1). It is important to note that the range change is at the global level.
Current scenario
The modelled results indicate medium to optimal climatic suitability for the eastern United States, Mexico, Brazil, Argentina, some other South American countries, Central and Southern Africa, most of Europe, Southern India, Eastern China and the Australian coasts. The Sahara Desert, central Australia, most of Canada, Mongolia and a large part of Russia are among the countries or territories with unsuitable conditions. The current modelled projection results show that over 60% of the African continent has some degree of climatic suitability for maize, compared with 77% of South America. Europe was projected to have the largest area of climatic suitability for maize cultivation, with almost 82% coverage (Table 2).
Future climate projections
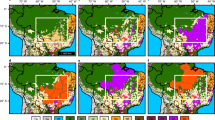
The future scenarios project a loss of climatic suitability area for maize in Sub-Saharan Africa and Latin America, but an expansion in the northern hemisphere, particularly in Europe (Fig. 2). The following section describes the results for 2050. Both GCMs project similar trends for America, Africa, Asia and Oceania. For South America, a shift in climatic suitability can be observed, with previously medium areas changing to marginal suitability. In North America, an increase in areas of marginal suitability have also been projected (Fig. 2a,c). Both GCMs project a loss of approximately 5% of general suitability in South America and an increase of approximately 10% in general suitability for North America (Table 2). In Africa, areas of climatic suitability for maize cultivation are projected to contract, with shifts from medium to marginal climate suitability in Angola, Zambia, Mozambique and Congo (Fig. 2a,c). The CSIRO model indicates a reduction of 11% in areas with climatic suitability, with this figure at 7% in the MIROC model (Table 2). In Asia, including the Middle East, climatic suitability for maize is projected to remain similar to current conditions. In India, marginal areas will become unsuitable, whereas in South East Asia, most medium suitability areas will change to marginal suitability by 2050. Bangladesh, Burma, Thailand, Laos and Cambodia will see a decrease in medium suitability areas and an increase in marginal suitability areas. An increase of 4% in climatically suitable areas for maize cultivation is projected under both GCMs for Asia, especially China (Fig. 2a,c) (Table 2) (Fig. S1).

The Ecoclimatic Index (EI) of future climate conditions for maize (Zea mays L.) under the SRES A2 (resembling RCP 8.5) in CSIRO: (a) Historical (b) in 2050 (c) in 2100; and in MIROC (d) in 2050 (e) in 2100. ArcMap 10.2 (http://desktop.arcgis.com/en/arcmap).
In Australia, areas of marginal suitability are projected to decrease in New South Wales, Queensland and Victoria under CSIRO (Fig. 2a). Larger decreases in areas with a medium climate for maize are projected under MIROC (5%) (Table 2). For Europe, the modelled projections under the two GCMs indicated slight differences. MIROC projected larger increases in the medium category compared to CSIRO, with medium areas extending to Russia and an increase in optimal suitability for England, France, Germany, Denmark, Netherlands, Poland, Slovakia and the Czech Republic. Both GCMs projected a change from unsuitable to marginal suitability in the Nordic Countries (Fig. 2a,c). The projected percentage of change under CSIRO and MIROC is similar, with increases of 10%. However, CSIRO projected an increase in the marginal suitability category, whereas under MIROC the increase corresponds mostly to optimal areas (Table 2).
Interestingly, Africa and Asia exhibited similar trends under both GCMs for 2100. For Africa, there is a considerable reduction in climatic suitability for maize. The Democratic Republic of Congo, Angola, Zambia, Mozambique Central African Republic, Cote d’Ivoire, Ghana, Togo and Nigeria may have small areas with marginal suitability remaining by the end of this century. Ethiopia, Kenya, Uganda, Tanzania and Madagascar could be some of the few remaining African countries with medium and optimal suitability by 2100 (Fig. 2b,d). Unsuitable areas will increase under CSIRO, with a 36% increment, compared to the 29% indicated under MIROC (Table 2). In Asia, reductions in climatic suitability for maize cultivation are projected for India, Malaysia, Singapore, Philippines and Indonesia. Mongolia and Southern Russia may become marginally suitable by 2100. Eastern China is projected to increase in optimal suitability, particularly under the MIROC scenario (Fig. 2b,d). The increase in suitability is low for Asia, at 7% under CSIRO and 11% under MIROC (Table 2).
By 2100, America, Europe and Oceania indicate some differences may occur between the two GCMs. Canada and the USA are projected to become more suitable for maize, with MIROC exhibiting larger increases in the south-eastern parts of Canada compared to CSIRO. Most Latin American countries are projected to experience a reduction in maize suitability, mainly in the optimal areas. Mexico, Brazil, Argentina, Paraguay and Peru may preserve some optimal and medium suitability areas. Interestingly, Uruguay may remain as optimally suitable until 2100 (Fig. 2b,d). Climatic suitability in North America is projected to increase by 19% and 20% under CSIRO and MIROC, respectively. South America is projected to decrease in suitability by 43% under both GCMs (Table 2). All of the European countries are projected to be suitable for maize to some extent. The MIROC model projects an important increase in optimal suitability in Scotland, England, France, Germany, Poland, Italy, Belarus and Russia. An increase in marginal suitability is expected for the Nordic countries and western Russia. CSIRO indicates a shift from marginal to medium suitability in northwestern Russia and in the Nordic countries, and an increase in the medium category in England and Italy (Fig. 2b,d). The percentage change in these areas is similar under both GCMs, with 16% under CSIRO and 14% under MIROC (Table 2). Finally, in Oceania, CSIRO predicts a three-times greater increase in unsuitable areas in Australia (30%) compared with MIROC (10%). New Zealand remains similar to current conditions (Fig. 2b,d) (Table 2).
The stresses were mapped to visualise the changes from the current scenario to the future projections. Cold stress is projected to decrease poleward at a global scale. Figure 3 shows the reduction in Europe and Asia. Heat stress may increase as a result of climate change, mainly in South America, Africa, parts of Asia and Australia. An illustration of the increase in heat stress is shown for Africa and some Asian countries in Fig. 3. Regarding dry stress, a worldwide increase is projected in accordance with a decrease in wet stress (Fig. 3). See also Fig. S3.

Projections of cold stress in Europe and Asia; (a) current scenario, (b) 2050, and (c) 2100; heat stress in Africa: (d) current scenario, (e) 2050, and (f) 2100; dry stress in South America: (g) current scenario, (h) 2050, and (i) 2100; and wet stress in Southeast Asia: (j) current scenario, (k) 2050, and (l) 2100. ArcMap 10.2 (http://desktop.arcgis.com/en/arcmap).
Current and future climate situation in the major maize-producing countries
According to the FAO, during the period from 1983 to 201371, the USA, China, Brazil, Mexico and Argentina were listed as the major maize producers. These countries are responsible for approximately 70% of the global maize production, with the USA and China together producing more than half of the total.
Under current climate conditions, approximately 50% of the global area, with some degree of suitability for maize, falls into the territories of these five major producers. Argentina has a large area of optimal suitability, whereas Brazil has larger areas in the medium category. The three remaining countries have mainly marginal suitable areas for maize (Table 3). Under projected future climate conditions, a reduction in optimal areas in Argentina is indicated. In the 2050 scenario, Argentina will see a reduction in climate suitability for maize, but may recover some by 2100 (Table 3). It is projected that cold and dry stresses may decrease in Argentina by 2100, and heat stress is not projected to occur in Argentina. Brazil, between 2050 and 2100, may see a radical reduction in climate suitability, principally in the medium and optimal categories due to an increase of dry and heat stresses, even when wet stress is expected to decrease and no changes are expected in cold stress. South of the Rio Grande do Sul, Parana and Sao Paulo remain as areas of optimal or medium suitability. A decrease of 64% in suitable areas is projected for Brazil (Table 3). China remains similar to current conditions in the 2050 scenario under both GCMs, with an approximately 5% increase in suitable areas. Under CSIRO for 2100, suitability in eastern China could increase by 13%, whereas under MIROC, this figure is as high as 20% (Table 3). For China, cold and wet stresses may decrease in future scenarios, whereas dry stress is projected to increase slightly. For Mexico, an increase of greater than 60% in unsuitable areas is projected by 2100 under both GCMs. Areas with medium suitability may be considerably reduced under future climate scenarios (Table 3). The USA exhibits a reduction in optimal areas, with an increase in areas in the medium category. Under CSIRO, a 16% increase in suitable climatic areas is projected for the USA, with MIROC indicating a more conservative increase of 9% (Table 3) (Fig. S2) In Mexico and the USA, cold and wet stresses are projected to decrease, contrary to heat and dry stresses, which will increase. Dry stress will cover almost the entire territory of Mexico by 2100, but only the western part of the USA. Figure 4 indicates the percentage of suitable areas for maize cultivation under the current scenario for the major five maize producers. The future scenarios indicate increases or decreases in this area for the major groups in relation to the current scenario.

Changes in areas of climatic suitability for maize among the current five major maize producers. For example, under current conditions China is modelled to have 48% of its territory with some climatic suitability for maize cultivation, with an increase of 4% expected by 2050 and 5% by 2100. ArcMap 10.2 (http://desktop.arcgis.com/en/arcmap).
Changes in areas of climatic suitability for maize among the current five major maize producers. For example, under current conditions China is modelled to have 48% of its territory with some climatic suitability for maize cultivation, with an increase of 4% expected by 2050 and 5% by 2100. ArcMap 10.2 (http://desktop.arcgis.com/en/arcmap).
Discussion
Validation and parameter sensitivity
This study was based on the biological and climatic requirements that cover all of the maize mega-environments. Taking into account these requirements, the model provides a good fit with the current global distribution of maize, including the validation area, which provided data that was not used to determine the bioclimatic requirements (India, China and neighbouring countries) (Fig. 1). In this study, the model represented approximately 96% of the occurrence records worldwide during the calibration phase and 88% of the occurrence records within the validation area matching the suitable areas for maize cultivation estimated using CLIMEX modelling.
The cross-validation performed with maps from MapSPAM (Spatial Production Allocation Model)70 and research into the geographic distribution of major crops69 generally matched with the modelled current global distribution of this study. The areas with high maize production from the MapSPAM map for rainfed conditions agree with the areas of optimal suitability in our model, such as the Maize belt of the USA70. This cross-validation provides further verification of our model, indicating consistency between the potential geographical distribution of the CLIMEX model and the global seasonal phenology of maize provided by You et al., (2014) and Leff et al. (2004).
The low percentage in the range change in the sensitivity analysis provides confidence about the chosen parameters34, 72. In summary, all of these tools support a satisfactory level of confidence in the maize model performance for current and future climate scenarios.
Current scenario
Even where the areas of climatic suitability estimated for the current conditions in this research match the known distribution, the CLIMEX maize model differs from other projections. The EcoCrop mechanistic model projects higher maize suitability in Africa, Australia and India and lower suitability in the Maize Belt of the USA, Europe and China under current conditions27, 73. This may be because the EcoCrop temperature thresholds differ from those used in this research. For example, the EcoCrop maximum temperature was set at 47 °C, whereas in this study, a value of 35 °C was used. This difference may be a result of the high level of biological diversity and climatic requirements of maize18 and of the model choice; for example, the Maize Belt of the USA has an average temperature between 26 to 34 °C, whereas in Ethiopia, Mexico and the Andean zone, the range is between 18 to 24 °C18. Furthermore, the EcoCrop niche projection for the USA does not predict high suitability in the Maize Belt of the USA; however, this region produces more than 30% of the world’s maize yield, mainly under rainfed conditions18. In contrast, the present model projects from medium to high suitability in this region.
Future scenarios
Regions with low temperatures will become warmer and more suitable for crop cultivation under predicted climate changes in the future as a consequence of new areas becoming available for agricultural production27, 47. These findings agree with the results of the present study, which indicate the expansion of climatic suitability for maize in cooler regions (northern latitudes) such as North America and Northern Europe, in agreement with a reduction in cold stress in future scenarios, under both GCMs for these regions. Currently, maize does not have optimal conditions in Northern Europe, but despite climate constraints, it has become established as a competitive crop in countries such as Denmark74. The Nordic countries started to cultivate maize mainly for silage, due to the short growing season and the low temperature limits for grain production. These countries are projected to increase maize cultivation into more northern latitudes as a result of the use of new hybrids and/or the climate change impacts75, 76. This change in suitability may increase the competitiveness of maize in cooler regions and may create conditions for grain production, not only silage.
The CERES (Crop Environment Resource Synthesis) crop model predicts a reduction in production of almost 12 million tons per year; representing 10% of the current production of Latin American and African countries by 2055, using data based on four maize varieties in rainfed conditions10. Such a prediction bears a direct relationship to the results of the present study in terms of the projected reduction of suitability in Latin America and Africa. Though production yields were not estimated in the present study, the level of climatic suitability is directly related to changes in maize production. Another study, using the SRES and HadCM3 GCM, with production functions that incorporate temperature, precipitation and the response to CO2, projects a large reduction (30%) in climatically suitable areas for maize in Africa1, bearing a similar relationship to the reduction of 32.5% projected in this study on this continent.
The A2 scenario (resembling RCP 8.5) describes increased rainfall and moderately increased temperature in most developed countries2, 47. Conversely, developing countries, mainly in the Southern Hemisphere, face decreased rainfall and greatly increased temperatures1. Such is the case for Honduras and El Salvador, which may suffer maize yield losses greater than 10% by 2020 due to projected global-warming factors77. It is crucial in that the availability of precipitation is rated as the most important constraint in maize productivity12. At the global level, the results of this research indicate a significant reduction in climatic suitability for maize cultivation in developing countries and expanded or greater suitability for some developed countries (USA, Canada, Europe and Australia). These previous results indicate comparative agreement with increases in heat stress over Africa and dry stress in Latin America, as noted in the projections in this study. Moderately higher temperatures, shortening of the growing cycle resulting in reduced dry matter in grain maize, and the increase in heat stress and dry stress are likely to be responsible for reduced yields and degraded climate suitability11, 12, 63.
As mentioned earlier, the top maize producers are the USA, China, Brazil, Argentina and Mexico71. The present model projects an increase of up to 16% in climatic suitability for maize in the USA, and China is also projected to have increased climate suitability. In China, maize is one of the major crops and is grown over an extended range of climatic conditions, from cold temperate to sub-tropical. During recent decades, there has been a tendency to increase the sown area and consequently the production yield in China38, 78. The more productive regions in China and the USA are those projected to increase in suitability, as shown by the present results. In contrast, our results indicate a reduction in areas of climatic suitability for maize in Brazil and Mexico. Some researchers have suggested that the availability of agricultural areas will be reduced due to climate change in northeastern Brazil and, as a consequence, productivity could be reduced79. By 2050, a 30% reduction in maize production is predicted for Brazil80; the present study projects a reduction of climate suitability of 5% by 2050 and a dramatic increase by the end of the century to 64%. Similarly, reports from Mexico suggest a reduction in rain-fed maize production and areas suitable for its cultivation81, 82. Finally, the present modelling predicted no significant change in Argentina for maize suitability; whereas some authors have predicted an increase in production due to climate change effects in Argentina83, 84. Travasso et al., (2009) modelled maize production in Argentina with the regional model MM5/CIMA and predicted increases in yield with the inclusion of CO2 effects, whereas without the inclusion of CO2 effects the production would reduce by 9% under SRES A2.
This future global scenario has important implications for food security in developing countries. However, development of new technologies and management adaptation could mitigate the impact of climate change on maize cultivation. For example, the development of stress tolerant varieties can reduce losses due to heat, drought, frost and hail in areas where conditions become unfavourable11, 78, and through adaptation measures, such as changes in planting date and water saving techniques84. Currently, varieties with elevated levels of drought tolerance have been introduced successfully in Asia and could be used in regions with similar characteristics6. Without investment and research into adaptation, developing countries are likely to suffer the negative impacts of climate change, with massive implications for food security among the populations of these countries85. It is important to mention that regardless of the differences between GCMs, the model outputs follow a similar trend of future climate predictions. This is because the fundamental basis is the same, despite some differences47.
Constraints of the study
Recently, in the AR5 report, the IPCC adopted representative concentration pathways (RCP) to replace the SRES. Moreover, new general circulation models (GCMs), such as the Coupled Global Climate Model (CGCM3), MIROC3.2, CNRM-CM3, CSIRO Mark 3.0, CM2.0-AOGCM, FGOALS1.0_g, INMCM3.0, Parallel Climate Model (PCM) and HadCM3 are becoming available. The new data from the RCPs can be used in various correlative species distribution models, such as MaxEnt, the Generalized Linear Model, Random Forest and Boosted Regression Trees; however, the available data for CLIMEX are only from two SRES (A1B and A2) and two GCMs (CSIRO Mk3.0 and MIROC-H). Shabani et al.86 explains the differences between performance of different correlative and mechanistic species distribution models.
We would also like to note that the CLIMEX model does not incorporate certain limiting abiotic and biotic factors, such as soil type, pests, pest interactions and weed impacts. Thus, one criticism of this study is that estimates of technical feasibility fail to identify the areas where it would be economically desirable to cultivate the species. Inclusion of non-climatic factors, such as topography, soil taxonomy, physicochemical properties of soil, and land usage on the national level, are required and could be included in future studies of maize suitability projections, either at global or regional levels. Furthermore, in the present study, the potential genetic progress of the species was not taken into account.
Conclusion
This study assessed the potential impacts of increased greenhouse gas emissions on climate suitability for maize cultivation, and consequently its distribution around the world. The use of CLIMEX allowed for a deeper understanding of the stress factors reducing the climate suitability of a species under future scenarios. Moreover, the high percentage of records in the validation area, as well as the sensitivity analysis, reduced the uncertainty in the CLIMEX model. Our main findings indicate that areas of climatic suitability for maize production are projected to increase in many developed countries (poleward areas), such as the USA, which is a major production area, due to a reduction in cold stress, whereas reductions in suitability are projected mainly for developing countries, such as Mexico and Brazil, which are two important production areas, and in many African countries, due to an increase in heat and dry stresses. Millions of people in Africa and Latin America depend on maize as their staple crop. Smallholders in these countries, as well as a portion of the population they feed, may be affected by the reduction in maize suitability, jeopardizing food security. The USA and China, two of the main maize producers (60%), are likely to see an increase in areas suitable for maize cultivation. This study indicate that heat and dry stresses may limit maize suitability in the future for countries between the tropics of Capricorn and Cancer; however, agricultural research and the genetics of plant cultivars could mitigate some of the negative effects of climate change for maize with new varieties that are tolerant to heat and drought conditions.
References
Parry, M. L., Rosenzweig, C., Iglesias, A., Livermore, M. & Fischer, G. Effects of climate change on global food production under SRES emissions and socio-economic scenarios. Global Environmental Change 14, 53–67, doi:10.1016/j.gloenvcha.2003.10.008 (2004).
IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 1132 (Cambridge, United Kingdom and New York, NY, USA, 2014).
Wheeler, T. & von Braun, J. Climate change impacts on global food security. Science 341, 508–513 (2013).
Ainsworth, E. A. & Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: mechanisms and environmental interactions. Plant, Cell Environ 30, 258–270 (2007).
Sánchez, B., Rasmussen, A. & Porter, J. R. Temperatures and the growth and development of maize and rice: a review. Global Change Biology 20, 408–417, doi:10.1111/gcb.12389 (2014).
Srinivasan, G., Zaidi, P. H., Singh, N. & Sanchez, C. Increasing productivity through genetic improvement for tolerance to drought and excess-moisture stress in maize (Zea mays L.). Water in Agriculture 116, 227–239 (2004).
Garcia, P. A. D. & Lopez, C. C. Temperatura base y tasa de extension foliar del maiz. Revista fitotecnia mexicana 25, 381–386 (2002).
Jaggard, K. W., Qi, A. & Ober, E. S. Possible changes to arable crop yields by 2050. Philosophical Transactions of the Royal Society of London B: Biological Sciences 365, 2835–2851 (2010).
Tripathi, A., Tripathi, D. K., Chauhan, D., Kumar, N. & Singh, G. Paradigms of climate change impacts on some major food sources of the world: A review on current knowledge and future prospects. Agric., Ecosyst. Environ 216, 356–373 (2016).
Jones, P. G. & Thornton, P. K. The potential impacts of climate change on maize production in Africa and Latin America in 2055. Global Environ. Change 13, 51–59, doi:10.1016/S0959-3780(02)00090-0 (2003).
Bassu, S. et al. How do various maize crop models vary in their responses to climate change factors? Global Change Biol 20, 2301–2320, doi:10.1111/gcb.12520 (2014).
Ammani, A., Ja’afaru, A., Aliyu, J. & Arab, A. Climate Change and Maize Production: Empirical Evidence from Kaduna State, Nigeria. Journal of Agricultural Extension 16, 1–8, doi:10.4314/jae.v16i1.1 (2013).
Doorenbos, J. & Kassam, A. H. Yield response to water. Vol. 33 (FAO, 1979).
Matsuoka, Y. et al. A single domestication for maize shown by multilocus microsatellite genotyping. Proceedings of the National Academy of Sciences 99, 6080–6084, doi:10.1073pnas.052125199 (2002).
Rebourg, C. et al. Maize introduction into Europe: the history reviewed in the light of molecular data. Theor. Appl. Genet. 106, 895–903, doi:10.1007/s00122-002-1140-9 (2003).
Reeves, T., Thomas, G. & Ramsay, G. Save and grow in practice: maize, rice, wheat. A Guide to Sustainable Cereal Production (FAO UN, 2016) (2016).
Hartkamp, A. et al. Maize production environments revisited: a GIS-based approach. (CIMMYT. The Maize Program, 2001).
Fischer, T., Byerlee, D. & Edmeades, G. Crop yields and global food security: Will yield increase continue to feed the world?, Vol. 158 (Australian Centre for International Agricultural Research (ACIAR), 2014).
Ortega, A. C. Insect pests of Maize. A guide for field identification. (CIMMYT, 1987).
Lone, A. A. & Warsi, M. Z. K. Response of maize (Zea mays L.) to excess soil moisture (ESM) tolerance at different stages of life cycle. Botany Research International 2, 211–217 (2009).
Abraha, M. & Savage, M. Potential impacts of climate change on the grain yield of maize for the midlands of KwaZulu-Natal, South Africa. Agric., Ecosyst. Environ 115, 150–160 (2006).
Byjesh, K., Kumar, S. N. & Aggarwal, P. K. Simulating impacts, potential adaptation and vulnerability of maize to climate change in India. Mitigation and Adaptation Strategies for Global Change 15, 413–431 (2010).
Conde, C. et al. Vulnerability of rainfed maize crops in Mexico to climate change. Clim. Res 9, 17–23 (1997).
Fisher, M. et al. Drought tolerant maize for farmer adaptation to drought in sub-Saharan Africa: Determinants of adoption in eastern and southern Africa. Clim. Change 133, 283–299 (2015).
Vanuytrecht, E. & Thorburn, P. J. Responses to atmospheric CO2 concentrations in crop simulation models: a review of current simple and semicomplex representations and options for model development. Global Change Biol (2017).
Hijmans, R., Guarino, L., Cruz, M. & Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genetic Resources Newsletter 127, 15–19 (2001).
Ramirez-Villegas, J., Jarvis, A. & Läderach, P. Empirical approaches for assessing impacts of climate change on agriculture: the EcoCrop model and a case study with grain sorghum. Agricultural and Forest Meteorology 170, 67–78, doi:10.1016/j.agrformet.2011.09.005 (2013).
Elith, J. et al. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17, 43–57, doi:10.1111/j.1472-4642.2010.00725.x (2011).
Merow, C., Smith, M. J. & Silander, J. A. A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography 36, 1058–1069 (2013).
Sutherst, R. & Maywald, G. A computerised system for matching climates in ecology. Agric., Ecosyst. Environ 13, 281–299 (1985).
Kriticos, D. J., Reynaud, P., Baker, R. H. A. & Eyre, D. Estimating the global area of potential establishment for the western corn rootworm (Diabrotica virgifera virgifera) under rain-fed and irrigated agriculture. OEPP/EPPO Bulletin 42, 56–64, doi:10.1111/epp.2540 (2012).
Kriticos, D., Sutherst, R., Brown, J., Adkins, S. & Maywald, G. Climate change and the potential distribution of an invasive alien plant: Acacia nilotica ssp. indica in Australia. J. Appl. Ecol. 40, 111–124 (2003).
Shabani, F., Kumar, L. & Taylor, S. Projecting date palm distribution in Iran under climate change using topography, physicochemical soil properties, soil taxonomy, land use, and climate data. Theoretical and Applied Climatology. 1–15, doi:10.1007/s00704-013-1064-0 (2014).
Kriticos, D. et al. CLIMEX Version 4: Exploring the Effects of Climate on Plants, Animals and Diseases. Canberra, ACT: CSIRO (2015).
Sutherst, R., Maywald, G. & Kriticos, D. CLIMEX version 3: user’s guide. Hearne Scientific Software Pty Ltd (2007).
Sutherst, R. W. Prediction of species geographical ranges. J. Biogeogr 30, 805–816 (2003).
GBIF. The Global Biodiversity Information Facility, http://www.gbif.org/ (2014). (Date of access: 28/07/2014).
Bai, J. et al. Evaluation of NASA satellite-and model-derived weather data for simulation of maize yield potential in China. Agron. J. 102, 9–16, doi:10.2134/agronj2009.0085 (2010).
Taylor, S., Kumar, L., Reid, N. & Kriticos, D. J. Climate change and the potential distribution of an invasive shrub, Lantana camara L. PloS one 7, e35565 (2012).
Shabani, F., Kumar, L. & Taylor, S. Climate change impacts on the future distribution of date palms: a modeling exercise using CLIMEX. PloS one 7, 1–12, doi:10.1371/journal.pone.0048021 (2012).
Kriticos, D. et al. Managing invasive weeds under climate change: considering the current and potential future distribution of Buddleja davidii. Weed Res. 51, 85–96, doi:10.1111/j.1365-3180.2010.00827.x (2011).
Gordon, H. et al. In CAWCR Technical Report No. 021 Vol. 021 74 (Centre for Australian Weather and Climate Research, 2010).
Shiogama, H. et al. Emission scenario dependency of precipitation on global warming in the MIROC3. 2 model. J. Clim. 23, 2404–2417, doi:10.1175/2009JCLI3428.1 (2010).
Kriticos, D. J. et al. CliMond: global high‐resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods in Ecology and Evolution 3, 53–64 (2011).
Raupach, M. R. et al. Global and regional drivers of accelerating CO2 emissions. Proceedings of the National Academy of Sciences 104, 10288–10293 (2007).
UN. World population to 2300. Vol. 236 (United Nations Publications, 2004).
IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. 996 (Cambridge University Press, Cambridge, 2007).
IPCC. Summary for Policymakers. Climate Change 2013. The Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change-Abstract for decision-makers. 28 (World Meteorological Organization, 2013).
van Vuuren, D. P. & Carter, T. R. Climate and socio-economic scenarios for climate change research and assessment: reconciling the new with the old. Clim. Change 122, 415–429 (2014).
Van Vuuren, D. P. et al. The representative concentration pathways: an overview. Clim. Change 109, 5–31 (2011).
Rogelj, J., Meinshausen, M. & Knutti, R. Global warming under old and new scenarios using IPCC climate sensitivity range estimates. Nature Climate Change 2, 248–253 (2012).
Basra, A. S. Crop responses and adaptations to temperature stress. (Food Products Press, 2001).
Miedema, P. The effects of low temperature on Zea mays. Advances in agronomy 35, 93–128 (1982).
FAO. Crop Water Information: Maize, http://www.fao.org/nr/water/cropinfo_maize.html (2014) (Date of access: 5/08/2014).
Hetherington, S., Smillie, R., Hardacre, A. & Eagles, H. Using chlorophyll fluorescence in vivo to measure the chilling tolerances of different populations of maize. Funct. Plant Biol. 10, 247–256 (1983).
Nguyen, H. T., Leipner, J., Stamp, P. & Guerra-Peraza, O. Low temperature stress in maize (Zea mays L.) induces genes involved in photosynthesis and signal transduction as studied by suppression subtractive hybridization. Plant Physiology and Biochemistry 47, 116–122, doi:10.1016/j.plaphy.2008.10.010 (2009).
Birch, C., Robertson, M., Humphreys, E. & Hutchins, N. Agronomy of maize in Australia: in review and prospect. Versatile Maize-Golden Opportunities: 5th Australian Maize Conference, 45–57 (2003).
Zaidi, P. H., Rafique, S. & Singh, N. Response of maize (Zea mays L.) genotypes to excess soil moisture stress: morpho-physiological effects and basis of tolerance. European Journal of Agronomy 19, 383–399, doi:10.1016/S1161-0301(02)00090-4 (2003).
Warrington, I. & Kanemasu, E. Corn growth response to temperature and photoperiod I. Seedling emergence, tassel initiation, and anthesis. Agron. J. 75, 749–754 (1983).
Smith, P., Bootsma, A. & Gates, A. Heat units in relation to corn maturity in the Atlantic region of Canada. Agricultural Meteorology 26, 201–213 (1982).
Bowen, C. R. & Hollinger, S. E. Geographic screening of potential alternative crops. Renew. Agric. Food Syst 19, 141–151, doi:10.1079/RAFS200473 (2004).
Blacklow, W. Influence of temperature on germination and elongation of the radicle and shoot of corn (Zea mays L.). Crop Sci. 12, 647–650, doi:10.2135/cropsci1972.0011183X001200050028x (1972).
Neild, R. E. & Newman, J. E. In National Corn Handbook. Vol. 40 Ch. 40, 10 (2014).
Lee, C. Corn Growth Stages and Growing Degree Days: A quick reference guide. University of Kentucky. http://www2.ca.uky.edu/agcomm/pubs/agr/agr202/agr202.pdf (2011) (Date of access: 05/08/2014).
Badu-Apraku, B., Hunter, R. & Tollenaar, M. Effect of temperature during grain filling on whole plant and grain yield in maize (Zea mays L.). Canadian Journal of Plant Science 63, 357–363 (1983).
Campos, H., Cooper, M., Habben, J., Edmeades, G. & Schussler, J. Improving drought tolerance in maize: a view from industry. Field Crops Res 90, 19–34, doi:10.1016/j.fcr.2004.07.003 (2004).
Grant, R., Jackson, B., Kiniry, J. & Arkin, G. Water deficit timing effects on yield components in maize. Agron. J. 81, 61–65, doi:10.2134/agronj1989.00021962008100010011x (1989).
Kriticos, D. et al. Balancing bioenergy and biosecurity policies: estimating current and future climate suitability patterns for a bioenergy crop. Global Change Biology, Bioenergy, 1–12, doi:10.1111/gcbb.12068 (2013).
Leff, B., Ramankutty, N. & Foley, J. A. Geographic distribution of major crops across the world. Global Biogeochemical Cycles 18 (2004).
You, L. et al. Spatial Production Allocation Model (SPAM) 2005 v2.0, http://mapspam.info (2014) (Date of access: 06/15/2016).
FAOSTAT, F. Agriculture Organization of the United Nations, http://faostat3.fao.org/home/E (2014) (Date of access: 14/10/2014).
Yonow, T., Kriticos, D. J., Ota, N., Van Den Berg, J. & Hutchison, W. D. The potential global distribution of Chilo partellus, including consideration of irrigation and cropping patterns. J. Pest Sci. 1–19 (2016).
EcoCrop. Forecasting potential niches for crop production, http://gisweb.ciat.cgiar.org/ClimateChange/EcoCropFB/ (2014) (Date of access: 8/11/2014).
Mikkelsen, M. & Halling, M. A. Cultivation of maize in cooler climates., 35–44 (Nordic Association of Agricultural Scientists, Kristianstad, Sweden, 2014).
Swensson, C. Use of maize in the Nordic and Baltic countries. 9–15 (Nordic Association of Agricultural Scientists, Kristianstad, Sweden, 2014).
Manevski, K., Børgesen, C. D., Andersen, M. N. & Kristensen, I. S. Parameterization of maize phenology, canopy development and dry matter partitioning in cool versus warm climate. 47–48 (Nordic Association of Agricultural Scientists., Kristianstad, Sweden, 2014).
Schmidt, A., Eitzinger, A., Sonder, K. & Sain, G. Tortillas on the Roaster (ToR): Central American Maize-Bean Systems and the Changing Climate. 123 (CIAT, CRS, CIMMYT, 2012).
Meng, E. C. H., Hu, R., Shi, X. & Zhang, S. Maize in China: Production systems, constraints, and research priorities., (IFAD - CIMMYT, 2006).
da Silva, Vd. P. R., Campos, J. Hd. C. & Silva, M. T. Climate risk in maize crop in the northeastern of Brazil based on climate change scenarios. Engenharia Agrícola 32, 1176–1183 (2012).
Costa, L. et al. Potential forcing of CO2, technology and climate changes in maize (Zea mays) and bean (Phaseolus vulgaris) yield in southeast Brazil. Environmental Research Letters 4, 014013, doi:10.1088/1748-9326/4/1/014013 (2009).
Tinoco-Rueda, J., Gómez-Díaz, J. & Monterroso-Rivas, A. Efectos del cambio climático en la distribución potencial del maíz en el estado de Jalisco, México. Terra Latinoamericana 29, 161–168 (2011).
Olivera, V. S. M. La productividad del maíz de temporal en México: repercusiones del cambio climático. 24 (División de Desarrollo Sostenible y Asentamientos Humanos CEPAL., Santiago de Chile., 2013).
Magrin, G. O., Travasso, M. I. & Rodríguez, G. R. Changes in climate and crop production during the 20th century in Argentina. Clim. Change 72, 229–249, doi:10.1007/s10584-005-5374-9 (2005).
Travasso, M. I., Magrin, G. O., Rodriguez, G. R., Solman, S. & Nunez, M. Climate change impacts on regional maize yields and possible adaptation measures in Argentina. International Journal of Global Warming 1, 201–213, doi:10.1504/IJGW.2009.027089 (2009).
Lobell, D. B. et al. Prioritizing climate change adaptation needs for food security in 2030. Science 319, 607–610 (2008).
Shabani, F., Kumar, L., Nojoumian, A. H., Esmaeili, A. & Toghyani, M. Projected future distribution of date palm and its potential use in alleviating micronutrient deficiency. J. Sci. Food Agric 96, 1132–1140 (2016).
Acknowledgements
This research was supported by the National Research Institute for Forestry, Agriculture and Livestock (INIFAP), the School of Environmental and Rural Science of the University of New England, Armidale, Australia, and AusAID Scholarships. The simulations were carried out using computational facilities at UNE.
Author information
Authors and Affiliations
Contributions
N.Y.Z.R.C. designed experiments, run models, analysed results, wrote and reviewed manuscript. F.S. and L.K. wrote and reviewed the manuscript. All authors reviewed the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ramirez-Cabral, N.Y.Z., Kumar, L. & Shabani, F. Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX). Sci Rep 7, 5910 (2017). https://doi.org/10.1038/s41598-017-05804-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-05804-0
This article is cited by
-
Capturing the Variability for Heat and Drought Tolerance in Wheat Using Multiple Selection Indices
Journal of Crop Health (2024)
-
Modelling the potential distribution of the Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae) using CLIMEX
International Journal of Tropical Insect Science (2024)
-
Climate change impacts on evapotranspiration in Brazil: a multi-model assessment
Theoretical and Applied Climatology (2024)
-
Tamarixia radiata global distribution to current and future climate using the climate change experiment (CLIMEX) model
Scientific Reports (2023)
-
Forecasting Brassica napus production under climate change with a mechanistic species distribution model
Scientific Reports (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
