
Abstract
Queen health is crucial to colony survival of honeybees, since reproduction and colony growth rely solely on the queen. Queen failure is considered a relevant cause of colony losses, yet few data exist concerning effects of environmental stressors on queens. Here we demonstrate for the first time that exposure to field-realistic concentrations of neonicotinoid pesticides can severely affect the immunocompetence of queens of western honeybees (Apis mellifera L.). In young queens exposed to thiacloprid (200 µg/l or 2000 µg/l) or clothianidin (10 µg/l or 50 µg/l), the total hemocyte number and the proportion of active, differentiated hemocytes was significantly reduced. Moreover, functional aspects of the immune defence namely the wound healing/melanisation response, as well as the antimicrobial activity of the hemolymph were impaired. Our results demonstrate that neonicotinoid insecticides can negatively affect the immunocompetence of queens, possibly leading to an impaired disease resistance capacity.
Similar content being viewed by others


Introduction
Honeybees are highly eusocial insects that build colonies of several thousand individuals which contain only one fertile female, the queen1. This queen is responsible for all egg laying and brood production within the colony; consequently, her integrity and health is crucial for the colony’s performance and survival, and any impairment can result in adverse effects on colony fitness. In the worst case, if the workers are unable to replace a failing queen, the colony will perish2,3,4. Recently, queen failure has been proposed as an important driver of honeybee colony mortality2, 4,5,6. While the natural lifespan of a honeybee queen is two to four years1, recent reports from the U.S.A. show high rates of early queen failure, with 50% or more being replaced in colonies within the first six months2, 3. This extremely high rate of queen failure coincides with high mortality rates of colonies in the U.S.A., where in some years more than 50% of colonies are dying2, 7.
Several stress factors are suspected to negatively affect survival of honeybee colonies. Parasites and pathogens are among the main factors4, but diet quantity, quality, and diversity8,9,10 as well as exposure to pesticides may also affect colony survival5, 6, 11. In particular, the widespread application of neonicotinoid insecticides12,13,14,15,16,17,18, has been suspected to represent a major threat to honeybee survival11, 19,20,21,22. Neonicotinoids are neurotoxins that act as agonists of the nicotinic acetylcholine receptor. They disrupt the neuronal cholinergic signal transduction, leading to abnormal behaviour, immobility and death of target pests12, 23, 24. Frequently, non-target insects like honeybees also come into contact with these systemic insecticides21. Exposure of bees to neonicotinoids is mostly through ingestion of residues in the pollen and nectar of contaminated plants11, 19. These pesticide residues are taken by the forager bees to their colonies and remain stored in beebread or honey until they are fed to larvae, workers, drones, or the queen4, 25.
The study of lethal and sub-lethal effects of neonicotinoid pesticides on social bees has largely focused on worker bees and, to a lesser extent, on overall colony function18, 25,26,27,28. Although reports of colony losses due to high rates of queen failure exist, only few studies address direct or indirect physiological effects in queens caused by pesticide exposure2, 5, 29, 30. Most studies focused on only three neonicotinoid insecticides, clothianidin, imidacloprid, and thiamethoxam, which are currently subject to a moratorium in the European Union31. Here, we also include thiacloprid, a cyano-substituted neonicotinoid which is considered as non-harmful for bees due to its much lower acute toxicity compared to other neonicotinoids. Thiacloprid is widely used in agriculture, for instance as spray application in fruit trees or directly into flowering oil seed rape during bee flight. It is the most abundant insecticide in beebread samples and has been detected in more than 60% of the samples analysed in the German Bee Monitoring Project32.
A strong immune defence is vital for honeybee health and colony survival. The strength of the individual immune defence depends on internal factors such as the nutritional state, the age, and the caste affiliation of the exposed individuals10, 33,34,35,36,37. In addition, the invasive ectoparasite Varroa destructor 38 impairs the immune defence of honeybees by reducing expression of immune-relevant genes and boosting viral replication, thereby affecting lifespan and disease resistance11, 39,40,41. The individual immunocompetence can also be weakened by environmental factors like pesticides that may render honeybees more vulnerable to parasites and pathogens41,42,43,44.
The exposure to pesticides is often associated with an increased pathogenic load, including the prevalent gut-parasite Nosema spp. and viruses typically associated with V. destructor, such as deformed wing virus (DWV)8, 11, 43, 45, 46. Neonicotinoids have been shown to affect the individual immunocompetence of honeybee workers. They negatively modulate NF-κB immune signalling and promote the replication of DWV44. Neonicotinoids have been shown to reduce hemocyte density as well as functional aspects of insect immunity as the melanisation of foreign objects, and the antimicrobial activity of the hemolymph47 in worker bees. However, there is only little information available about the individual immunity of honeybee queens. In queens, phenoloxidase (PO) enzyme levels continuously increase with age and reach levels twice as high as those found in workers48. Gätschenberger and colleagues49 examined the antimicrobial defence systems of queens and found that reactions of young queens to bacterial immune challenges resemble those of worker bees. In this study we examine the effects of pesticide exposure on young queens. Our main question is whether general immune defence mechanisms are affected by sublethal concentrations of the neonicotinoids thiacloprid or clothianidin. Since disease resistance is difficult to measure directly33, we selected established functional parameters of immunity to analyse honeybee immunocompetence, namely total and differential hemocyte counts, melanisation response, and antimicrobial activity of the hemolymph.
Results
Sublethal effects and exposure
To date, acute toxicity tests are solely performed with worker bees5, 25. To determine whether the dosages of neonicotinoids used in the immune assays are indeed sublethal to queens, the number of dead queens was recorded after seven days of exposure. We found no significant difference between the treatment groups regarding the survival of the queens (Supplementary Fig. S1, Supplementary Table S2). Thiacloprid exposure had no lethal effect on worker bees. However, there was a significant effect of clothianidin on worker bee mortality (χ2-test; p = 0.009). Hence, the highest concentration of clothianidin (200 µg/l) was excluded from immune tests.
To estimate the exposure level to neonicotinoids, the food consumption was measured. We found no significant difference between the treatment groups regarding the consumption of sugar solution or pollen (Supplementary Fig. S2, Supplementary Table S2). Moreover, the behaviour of the bees was observed regularly. When the food was changed, the queens were surrounded by attending workers and fed via trophallaxis. During these short observations, we never saw a queen directly feeding at the syringe containing the spiked sugar solution.
The wellbeing of a queen depends on the care and the adequate supply with food (royal jelly and honey1) by the attending worker bees. However, neonicotinoids have been shown to reduce the size of the hypopharyngeal gland (HPG) of nurse bees. To examine, whether thiacloprid or clothianidin affected the HPG size of the attending worker bees in our experiments, the acinus diameter of age defined worker bees were measured. Interestingly, no significant difference was found in the HPG size of worker bees after seven days of exposure in a cage, attending to a queen (Supplementary Fig. S3, Supplementary Table S2; KWT, p > 0.05).
Total and differential hemocyte counts
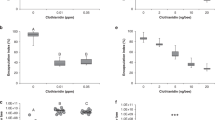
Exposure to relevant concentrations of both neonicotinoids significantly reduced the total hemocyte counts of young queens (Fig. 1a,b). The median total hemocyte counts of thiacloprid treated queens were lower than those of control queens (Fig. 1a, Kruskal–Wallis test (KWT), p = 0.011). Untreated control queens displayed a higher hemocyte density than queens treated with 200 µg/l thiacloprid (59% of control value; Supplementary Table S1; MWU, p = 0.022; control: median = 4250 hemocytes/µl (h/µl), n = 19; 200 µg/l thiacloprid: median = 2500 h/µl, n = 16), or treated with 2000 µg/l thiacloprid (47% of control value; MWU, p = 0.024, median = 2000 h/µl, n = 19).

Exposure to thiacloprid or clothianidin reduced total hemocyte counts. The seven day-treatment of young honeybee queen with thiacloprid ((a); control: n = 19, 200 µg/l: n = 16, 2000 µg/l: n = 19), or clothianidin ((b); control: n = 19; 10 µg/l: n = 15; 50 µg/l: n = 14) reduced the total hemocyte counts compared to control queens. Boxes show 1st and 3rd interquartile range with black lines denoting medians. Whiskers encompass 95% of the individuals, beyond which outliers (circles) reside. Treatments with different letters differ significantly from each other.
Total hemocyte counts of queens treated with clothianidin were lower than in control queens (Fig. 1b, KWT, p = 0.0005), with control queens displaying a higher hemocyte density than queens treated with 10 µg/l clothianidin (53% of control value; MWU, p = 0.015; control: median = 4250 h/µl, n = 19; 10 µg/l clothianidin: median = 2250 h/µl, n = 15), or treated with 50 µg/l clothianidin (47% of control value; median = 2000 h/µl, n = 14, MWU, p = 0.014).
To evaluate changes in the composition of subclasses in the hemocyte population, the cells were classified using the morphological characteristics described by Negri et al.36, 50, supported by phalloidin staining of the actin cytoskeleton and DAPI staining of the nuclear DNA (Fig. 2a–d). W1-like hemocytes50 contained a large oval or irregular shaped nucleus and prominent vesicle-like structures. These granulocyte-like hemocytes36, 50 had a well-developed actin cytoskeleton marked by intensive f-actin staining, and showed extreme cellular spreading as indicated by lammelipodia and numerous filopodia-like structures (Fig. 2a,b). The hemocytes of the subclass W2 had large, round or oval shaped nuclei with decondensed chromatin, but did not contain vesicle-like structures (Fig. 2a,c,d). Some W2-like hemocytes showed spreading with an intensively stained actin cytoskeleton, lammelipodia formation, and filopodia-like structures (Fig. 2c). Hemocytes of the W3-like subclass were the most abundant ones. They were small, of oval or round shape and contained small nuclei with highly condensed chromatin. In contrast to the other cell types, W3-like cells showed no spreading and had only very weak f-actin staining (Fig. 2a,b,d). Queen hemocytes classified as W4-like had a spindle-shaped morphology with well-developed actin cytoskeleton and small, elongated or irregular shaped nuclei (Fig. 2a,d).

Exposure to neonicotinoids differentially affected hemocyte subclasses. (a–e) Queen hemocytes showing staining of the filamentous actin cytoskeleton (red: phalloidin-staining) and nuclear DNA (blue: DAPI-staining). (a) Compound image. W1-like hemocytes contained vesicle-like structures and showed extreme cellular spreading, lammelipodia formation, and filopodia-like structures with a well developed cytoskeleton marked by intensive f-actin staining (a,b; W1). W2-like hemocytes contained no vesicle-like structures, but also showed spreading and strong f-actin staining (a,c; W2). The nuclei of W1- or W2-like hemocytes were large and contained decondensed chromatin. W3-like hemocytes were oval/round and had only very weak f-actin staining. Nuclear chromatin of W3 like cells was highly condensed (a,b,d; W3). W4-like hemocytes had a spindle-shaped morphology and showed intensive f-actin staining of the cytoskeleton and elongated nuclei (a,d; W4). (e) Thiacloprid (200 µg/l) or clothianidin (10 µg/l) treatment affected the composition of the hemocyte population (8 to 9 individuals per treatment group, at least 350 cells per individual counted). Thiacloprid or clothianidin exposure significantly reduced the number of W1-type hemocytes. Clothianidin treated queens had a lower percentage of W2-like cells and a higher percentage of W3-like cells compared to control queens (error bars = s.e.m; *p ≤ 0.05; **p ≤ 0.01; scale bar 10 µm).
Pesticide treatment differentially affected the relative abundance of hemocyte subclasses. Exposure to 200 µg/l thiacloprid and 10 µg/l clothianidin reduced the percentage of W1-like cells in treated queens compared to controls (Fig. 2e, KWT, p = 0.0005; MWU, control vs. 200 µg/l thiacloprid: p = 0.002, control vs. 10 µg/l clothianidin: p = 0.034; control: average = 3.356%, n = 9; 200 µg/l thiacloprid: average = 1.980%, n = 9; clothianidin: average = 0.394%, n = 8). Exposure to 10 µg/l clothianidin produced a detectable reduction of W2-like hemocytes (Fig. 2e, KWT, p = 0.0384; MWU, control vs. 200 µg/l thiacloprid: p > 0.05, control vs. clothianidin: p = 0.06; control: average = 14.17%; 200 µg/l thiacloprid: average = 7.97%; clothianidin: average = 6.67%). Clothianidin treated queens showed a relative increase of undifferentiated, W3-like hemocytes (Fig. 2e, KWT, p = 0.0408; MWU, control vs. 200 µg/l thiacloprid: p > 0.05, control vs. clothianidin: p = 0.077; control: average = 69.06%; 200 µg/l thiacloprid: average = 72.73%; clothianidin: average = 83.27%). There was no significant difference between the treatment groups regarding the spindle-shaped W4-like hemocytes.
Melanisation
Compared to control queens, the melanisation response of queens treated with neonicotinoids was significantly reduced at all concentrations tested (Fig. 3). The melanisation in thiacloprid treated queens was reduced to 41% (200 µg/l) or 23% (2000 µg/l) of the control value (Fig. 3a, KWT, p = 0.001; MWU, control vs. 200 µg/l thiacloprid: p = 0.016, control vs. 2000 µg/l thiacloprid: p = 0,003; control: median = 23.83% grey value (gv), n = 20; 200 µg/l thiacloprid: median = 10% gv, n = 17; 2000 µg/l thiacloprid: median = 5.5% gv, n = 15). The melanisation in clothianidin treated queens was reduced to 31% (10 µg/l) or 29% (50 µg/l) of the control value (Fig. 3b, KWT, p < 0.0001). Control queens showed a stronger melanisation than queens treated with clothianidin (Mann–Whitney U test (MWU), control vs. 10 µg/l clothianidin: p < 0.0001, control vs. 50 µg/l clothianidin: p = 0.002; control: median = 23.83% gv, n = 20; 10 µg/l clothianidin: median = 7.5% gv, n = 19; 50 µg/l clothianidin: median = 7.00% gv, n = 18).

Exposure to thiacloprid or clothianidin reduced melanisation response. The seven day-exposure of young honeybee queens to thiacloprid ((a); control: n = 20, 200 µg/l: n = 17, 2000 µg/l: n = 15), or clothianidin ((b); control: n = 20; 10 µg/l: n = 19; 50 µg/l: n = 18) reduced the melanisation of an implanted nylon filament compared to control queens. Boxes show 1st and 3rd interquartile range with black lines denoting medians. Whiskers encompass 95% of the individuals, beyond which outliers (asterisks) reside. Treatments with different letters differ significantly from each other.
Antimicrobial activity of the hemolymph
The antimicrobial activity of the hemolymph, measured as the size of the inhibition zones, was significantly reduced in queens treated with all concentrations of thiacloprid or clothianidin compared to control queens (Fig. 4, KWT, p = 0.002). Inhibition zones of queens exposed to thiacloprid were significantly smaller than in control queens (89% of control value; MWU, p = 0.008; control: median = 19.37 mm, n = 15; 200 µg/l thiacloprid: median = 17.25 mm, n = 16), or treated with 2000 µg/l thiacloprid (84% of control value; MWU, p = 0.003, median = 16.42 mm, n = 15). Inhibition zones of queens treated with clothianidin were significantly smaller than in control queens (Fig. 4b, KWT, p < 0.001), with control queens displaying a larger inhibition zone than queens treated with 10 µg/l clothianidin (89% of control value; MWU, p < 0.001, median = 17.26 mm, n = 14), or treated with 50 µg/l clothianidin (85% of control value; MWU, p < 0.001, median = 16.54 mm, n = 14).

Neonicotinoid exposure reduced antimicrobial activity of hemolymph. The hemolymph inhibited the growth of grampostive bacteria (Micrococcus flavus) on agar plates. The seven day-treatment with thiacloprid ((a), control: n = 15: 200 µg/l: n = 16; 2000 µg/l: n = 15) or clothianidin ((b), control: n = 15: 10 µg/l: n = 14; 200 µg/l: n = 14) reduced the antimicrobial activity of the hemolymph, the diameter of the inhibition zones being smaller than in control queens. Boxes show 1st and 3rd interquartile range with black lines denoting medians. Whiskers encompass 95% of the individuals, beyond which outliers reside (asterisks). Significant differences indicated with letters.
Discussion
In this paper we report immunosuppressive effects of two neonicotinoid pesticides on general immune parameters of honeybee queens. We employed a broad array of methods to investigate the immune defence competence of queens: total and differential hemocyte counts, wound healing/melanisation, and antimicrobial activity of the hemolymph after immune stimulation. Our results indicate that the tested aspects of individual immunity are negatively affected by sublethal, environmentally relevant concentrations of neonicotinoids in newly emerged honeybee queens.
Hemocytes are the key components of insect cellular immune defence. They are responsible for phagocytosis or encapsulation of pathogens and for the closure of wounds36, 51. The mechanisms of cellular immunity in honeybees are not yet completely understood, mostly due to limited information about the cellular subtypes, their functions and abundances50, 52, 53. We found an overall reduction of hemocyte density at all concentrations tested. Based on morphological characterisation, we also observed changes in the abundance of subclasses in the population of hemocytes in clothiandin exposed queens, which may indicate a severe interference of neonicotinoid treatment with cellular immunity.
The staining of the actin cytoskeleton by phalloidin together with nuclear staining can be used as a valuable tool for hemocyte characterization. Granulocytes are the first type of hemocyte to attach to a foreign object during the encapsulation response50. We found a significant reduction in the number of granulocyte-like hemocytes in queens exposed to clothianidin. As in workers50, these granulocyte-like cells showed extreme spreading with extensive filopodia and lammellipodia formation and contained numerous vesicle-like structures which may indicate their involvement in phagocytosis36, 50. The reduction of granulocyte-like hemocytes may be particularly severe, since this type of active, differentiated hemocyte is considered responsible for the elimination of pathogens and is the first cell type to initiate the encapsulation process36, 54. In addition, W2-like hemocytes tended to be reduced in queens treated with clothianidin. The W2 hemocytes have been shown to be the second cell type which adheres to foreign surfaces, a specific function of this cell type has not yet been described50.
Hemocytes of the types W3 and W4 are normally freely floating in the hemolymph and interact with adherent granulocytes to form cell agglomerations36. We found a relative increase of W3-like cells in queens exposed to clothianidin. The overall number of W3- or W4-like hemocytes may be underestimated in our study, since most likely not all of them adhered to the surface of the glass cover slip36. In conclusion, an altered hemocyte density together with a reduction in specific subclasses of active, differentiated hemocytes caused by neonicotinoid exposure could likely impair the cellular immune defence, hence increasing a queen´s susceptibility towards pathogens. Nevertheless, honeybee hemocyte typing remains complex and it is difficult to compare the results obtained by different methods50, 52, 53. More detailed investigations are necessary to bring light into the process of hemocyte differentiation in honeybees, combining the development of reliable cell markers and time-lapse microscopy.
A central immune defence mechanism mediated by hemocytes is the encapsulation and melanisation of intruding pathogens and the closure of wounds. The melanisation reaction is catalysed by the enzyme phenoloxidase whose precursor prophenoloxidase is produced by hemocytes51. We observed a significantly reduced melanisation after treatment with neonicotinoids in all substances and concentrations tested. The ability to melanise and encapsulate a foreign object is positively correlated with resistance to viral infections, parasitoids, and parasites. Wound closure involves similar mechanisms as encapsulation and melanisation and is important for reducing virus transfer between bees51, 55. In addition, we showed that exposure to thiacloprid or clothianidin significantly reduced the antimicrobial activity of the hemolymph. The antimicrobial activity depends on the amount of antimicrobial peptides in the hemolymph, which are produced by hemocytes or fat body cells55. Together with the observed decrease in the melanisation response and changes in hemocyte composition and density, our findings may be interpreted as impairments of immune defence and disease resistance capacity of honeybee queens during exposure to neonicotinoids.
In our cage experiments, exposure to thiacloprid and clothianidin significantly affected the immunocompetence of young queens, even in sublethal concentrations as low as those reported from samples collected by honeybees. The thiacloprid or clothianidin concentrations fed over a period of seven days showed no lethal effects on queens and were therefore considered as sublethal for queens. Only the extremely high concentration of 200 µg/l clothianidin increased the mortality of attending worker bees and was therefore excluded from further analysis. According to Sanchez-Bayo and Goka56, mean thiacloprid residues are 75.1 µg/kg (max.: 1002 µg/kg, mean prevalence of 17.7%) in pollen, and 6.5 µg/kg in honey (max.: 208 µg/kg, 64% prevalence)56. Mean clothianidin residues in pollen were 9.4 µg/kg (max.: 41.2 µg/kg, mean prevalence of 11%) and in honey 1.9 µg/kg (max.: 10.1 µg/kg, mean prevalence of 17%). In the German bee monitoring, the highest observed concentrations of thiacloprid in beebread samples were 498 µg/kg32. Thiacloprid is frequently detected in honey samples, up to concentrations of 200 µg/kg4, 25.
In a colony the queen is constantly surrounded by attending worker bees which take care of her wellbeing and constantly supply her with high-quality food such as royal jelly. A queen can survive without worker bees for only a few days. However, this is a non-natural situation with unknown effects on the immune status of a queen. To approximate the natural situation in a laboratory experiment, ten attending bees were placed into each cage to provide for the queen. However, this makes it difficult to discern between the direct effects of neonicotinoids on the immune system of queens and indirect effects that may come from affected worker bees. In the concentrations used in immune assays, the mortality of queens or workers was not increased. Moreover, food consumption of sugar solution or pollen was comparable in all treatment groups, indicating normal feeding behaviour. Interestingly, in the presence of a queen, the neonicotinoids had no significant effect on the size of the HPG compared to controls. The mechanism by which xenobiotics like neonicotionoids, fungicides or varroacides affect HPG size is still unknown. Our findings may imply that the stimulus provided by the queen may counteract the HPG-reducing effects described for neonicotinoids. Although we cannot rule out any possible indirect effect, when considering the normal survival rate, the normal food consumption and the normal size of the HPGs together, we have no indications that our experimental queens may have suffered from a lack of care or feeding.
Adult queens receive mostly royal jelly from nurse bees, possibly with some additional honey1, 57. Contaminated food reaches queens through trophallaxis. The contamination might originate from the transfer of active substances through the mandibular and hypopharyngeal glands located in the heads of nurse bees or by the addition of contaminated honey58. Although little is known about pesticide contamination of royal jelly, neonicotinoids can be found in the heads of bees were the glands are located26. Neonicotinoids have also been detected in brood food59 and royal jelly when bees were exposed to contaminated pollen40. In addition, neonicotinoids can reduce the size of the royal jelly producing hypopharyngeal glands in nurse bees60, 61, thus queens may be indirectly affected by altered food quality or quantity.
It is not clear, whether queens in laboratory assays or in the social context of a colony are exposed to the same dosages of pesticides as worker bees. In a honeybee colony, the queen may be shielded from harmful agents like pesticides by the attending workers. Transmission experiments suggest that behavioural patterns are in place to protect the queen from viral infection by symptomatic bees. Either queens avoid direct contact with or uptake of food from diseased workers, or diseased workers try to avoid interaction with the queen62. It is not known, whether similar behavioural mechanisms may exist in a colony that prevent or reduce exposure of queens to pesticides.
To estimate the level of exposure to queens, the detoxification of neonicotinoids has also to be taken into account. In case queens failed to fully clear ingested pesticides from their bodies, the persistence of even small daily intakes could eventually accumulate to harmful or even lethal levels over time. Indeed, the lethality of the neonicotinoid imidacloprid to worker bees, ants or termites appears to be dependent on the duration of exposure: the longer the exposure time, the less amount of pesticide is needed to kill the worker63. So far, the temporal dynamics of the decrease of neonicotinoid residues has only been studied in worker bees26. To our knowledge, no data are available concerning the ability of queens to detoxify pesticides.
A honeybee colony represents an environment with a high chance for the spread of infections, because of its highly organised social structure and crowded population density64. In honeybees, transmission of viruses can occur horizontally among workers, or vertically, from the queen to her offspring. In queens, numerous viruses have been detected, e.g. DWV, chronic bee paralysis virus (CBPV), black queen cell virus, Kashmir bee virus, and sacbrood virus64. In a recent sanitary survey in Belgium, 75% of the eggs tested had at least one virus present65. It appears plausible that a queen with a weakened immune defence may be prone to infections, which she can transmit vertically to her offspring.
In honeybees, like in other insect species, a maternal immune experience can be transmitted to the progeny. This so called trans-generational immune priming has been demonstrated to have a positive impact on offspring resistance and survival of infections66, 67. Whether a queen with weakened immune defence can still sufficiently protect her offspring via these mechanisms has yet to be investigated. Since a queen lays between 175.000 or 200.000 eggs annually1, the health status of a queen is highly relevant for the overall colony health.
A young queen has to master several challenges in the early phase of her life. She has to fight her competitive sisters, perform risky nuptial flights, successfully mate with a sufficient number of drones, and start laying fertilised eggs in an adequate number and quality to be accepted by the colony1. Especially when the queen gets into contact with drones on her nuptial flights, she is potentially confronted with an additional load of pathogens68. We do not know whether the immunosuppressive effect of neonicotinoids may affect the ability of a queen to accomplish these tasks. The impact on the health and fitness of a queen may depend on the phase of life and the duration of the exposure to pesticides. Exposure to neonicotinoid pesticides during larval development of queens can have severe effects on her performance later in life: clothianidin and thiamethoxam treatment during the larval phase affects ovary size and reduces the number and quality of stored spermatozoa within queen spermathecae, which results in reduced egg laying success5. In our study, we analysed the immunocompetence of newly hatched queens after only seven days of exposure to neonicotinoids. However, in the field, queens may be exposed to pesticides for several months, including the winter season69. Whether queens are susceptible to neonicotinoids during all phases of their life, or whether the immunosuppressive effects are persistent or reversible has yet to be determined.
Immune suppression by pesticides like neonicotinoids opens the way to the spread and abundance of pathogens and parasites, which are the proximate mortality factors of honeybee colonies11, 36. Infestation with Varroa destructor, the suspected main cause for colony losses, was shown to be promoted by exposing colonies to neonicotinoid treated crops70. This study on the immunocompetence of queens complements previous, similar findings on the immunosuppressive effects of neonicotinoids on worker bees41, 47. Currently, regulatory requirements for evaluating the safety of pesticides to honeybees do not consider effects on the immune defence on workers, drones, or queens71. Given the key importance of queens to colony health and survival, the general lack of knowledge concerning both lethal and sub-lethal effects of pesticides on queen physiology is alarming. Our findings highlight the vulnerability of honeybee queens to common neonicotinoid pesticides, and demonstrate the need for future studies to identify relevant measures of queen health and disease susceptibility. Improving the understanding of honeybee immunity could provide new insights into the stress factors and their interactions that threaten honeybee survival and ultimately enable us to design strategies to protect them11.
Material and Methods
Rearing of Queens
Queens were produced using standard honeybee queen-rearing techniques72. Briefly, young grafted larvae were introduced into queenless starter colonies. After 5 days, sealed queen cells were transferred to an incubator (35 °C, 65% humidity; Grumbach, Asslar, Germany). Sister queens from eight different maternal lines were distributed equally among the treatment groups. After emergence, queens were visually inspected and transferred into standard metal cages (8.5 × 6.5 × 4 cm,) together with ten attendant workers from healthy experimental colonies that tested negative for virus infections (DWV, acute bee paralysis virus, CBPV, sac brood virus)4, 62.
Neonicotinoid exposure in laboratory cage experiments
The cages were supplied with water and pollen (collected at the Bee Institute Kirchhain, Germany), and ad libitum sugar syrup (Apiinvert, Mannheim, Germany) diluted to a 50% sugar solution (w/v with ambrosia Bienenfutter, Germany and distilled water) in a 5 ml syringe (Carl Roth, Karlsruhe, Germany). Cages were kept in an incubator (Binder, Tuttlingen, Germany; humidity provided by open water jars) in the dark at 33 °C73.
Thiacloprid was obtained from Sigma Aldrich (St. Louis, USA; analytical standards, purity 99.9%). A stock solution of 2 mg/ml thiacloprid in acetone was prepared in a glass flask and stored in the dark at room temperature (~15 °C) until use. A sugar solution (50% w/v) was prepared and thiacloprid stock solution was added to reach a final concentration of 100 or 200 µg/l thiacloprid. The final concentration of acetone in the feeding solutions was adjusted to 0.0086% (v:v) in all thiacloprid treatment groups, including the control. Clothianidin (Sigma Aldrich, analytical standards, purity 99.3%) was dissolved in water and added to the sugar solution (50% w/v) to obtain a final concentration of 10, 50, or 200 µg/l clothianidin. The pollen pastry was prepared from pollen collected at the Bee Institute Kirchhain or obtained from Imkereibedarf Bährle (Aschaffenburg, Germany).
Honeybees of each cage (= a queen with ten worker bees) were exposed to one of these concentrations for seven days. During the phase of exposure, queens remained within the group of the attendant bees in the cages. In that way queens were exposed to the dietary pesticide either directly by taking up contaminated food themselves or indirectly via the food (e.g. royal jelly) received from the attending worker bees. The consumption of food was recorded daily. To quantify the food consumption, the syringes containing the sugar solution and the caps containing the pollen were weighed after 24 hours in the cage. Food consumption was calculated by dividing the total amount of food consumed in 24 hours divided by the number of workers in the cage. Three empty cages contained an evaporation control. Dead individuals were removed every 24 or 48 hours (for details see Supplementary Table S2).
Subsequently, the immunocompetence of the queens was evaluated by one of the methods: quantification of hemocytes, antimicrobial activity of the hemolymph, or melanisation response. Any individual queen was used for only one single immune test. Each test was replicated at least three times with at least 14 queens per treatment group (details see below and Supplementary Table S1).
Hemolymph collection
Queens were anesthetized on ice before hemolymph was collected by inserting a microinjection needle (Hartenstein, Würzburg, Germany) into the proximal abdomen. Any fluid which appeared yellow or brown was discarded and excluded from further analysis as this was likely not hemolymph but gastric fluid33.
Total and differential hemocyte counts
Total hemocyte counts were performed as an indirect measurement of cellular immunocompetence33. For total hemocyte counts, 1 µl of hemolymph was transferred to a PCR-tube (Biozym, Hessisch Oldendorf, Germany) containing 3 µl PBS (pH 7.4; Sigma Aldrich, St. Louis, USA) and 1 µl of DAPI-staining solution (4′,6-diamidino-2-phenylindole; 1:100 dilution, lifetechnologies, Carlsbad, California, USA). Immediately after collection, the diluted hemolymph solution was transferred to a Bürker counting chamber (Carl Roth, Karlsruhe, Germany), where hemocytes were counted (average of five squares per queen) under a phase contrast/fluorescent microscope (Leica DMIL, Leica camera DFC 420 C). To verify the cellular character of the observed structures, DAPI staining was used as counterstaining of nuclear DNA47. Each experiment was repeated five times with 14 to 19 queens per treatment group.
To analyze whether exposure to pesticides affects the composition of the hemocyte population, differential hemocyte counts were conducted. The hemolymph (3 µl) was placed on sterile glass cover slips in a cell culture 24-multiwell plate (Sigma Aldrich) with distilled water in the spaces between the wells. The multiwell plate was placed in the incubator for 4 hours at 33 °C. Subsequently, the cells were fixed in formaldehyde solution (4% formaldehyde in PBS, Carl Roth) over night at 4 °C, rinsed three times in PBS and permeabilized in PBS + 0.2% Tween-20 (Sigma Aldrich) for 10 min. To visualize the cytoskeleton and the nuclei, cells were stained with Alexa 555-conjugated phalloidin at 1:200 (Invitrogen, Carlsbad, USA) and DAPI at 0.3 µM for 30 min, washed with PBS and mounted in Vectashield (VWR International, Darmstadt, Germany). Queen hemocytes were visualized using a confocal microscope (Zeiss LSM 700, Jena, Germany) or observed under the phase contrast/fluorescent microscope. Following the description and classification of Negri et al.50, we classified the queen hemocytes using morphological characteristics (eight to nine queens per treatment group, 351 to 530 cells counted per individual; for details see Supplementary Table S1).
Melanisation
To provoke a melanisation response, a nylon filament was partly inserted into the abdomen of a queen, thus mimicking the behaviour of Varroa destructor as previously described by Brandt et al.47. The strength of the immune reaction was measured by the degree of melanisation on the filament. Briefly, a nylon fishing line (0.2 mm diameter, Nexos, Naila, Germany) was cut into approximately 2.5 mm long segments and sterilized in 100% pure ethanol (Roth, Karlsruhe, Germany). Queens were anesthetized on ice, and the nylon filament was implanted in the abdomen through the intersegmental membrane between the 3rd and 4th tergum33, 74. After implantation, queens were transferred to a 2 ml microcentrifuge tube (Eppendorf, Hamburg, Germany) with holes poked through cap and sidewalls. After approx. four hours, the nylon filament was extracted, fixed in formaldehyde solution for at least 1 hour, rinsed three times in PBS, and subsequently mounted in glycerol (85%, Carl Roth). Each experiment was repeated four times (17 to 19 queens per treatment group). Three pictures per explant were taken at different focal depths. The mean grey value per filament served as a measure of melanisation and was quantified for the inserted portion of the filament using image analysis software73. The mean grey value of an non-implanted filament that served as background value was subtracted from the mean grey value of the implanted filaments47. Each treatment group (for details see Supplementary 15 to 20 queens per treatment group (for details see Supplementary Table S1).
Inhibition-Zone Assay
For inhibition zones assays, queens were exposed to neonicotinoids for seven days. On day six of the exposure, the immune system was challenged by the injection of 1 µl of heat-inactivated Escherichia coli (grown to OD 0.5). As previously described47, hemolymph was collected and stored at −20 °C until the assay was conducted. Antibacterial test plates (ø 9 cm) were prepared by adding 0.8 ml of live Micrococcus luteus bacteria suspension (OD 0.5) to 150 ml of sterile broth medium (48 °C, 1.5 g Agar No. 1, Oxoid; 3.75 g nutrient broth, Applichem). Per test plate, five holes (ø1 mm) were punched into the medium and 1 µl of hemolymph solution was added to each one. The plates were incubated at 38 °C overnight and the diameter of inhibition zones was measured. Each experiment was repeated three times with 14 to 16 queens per treatment group.
Hypopharyngeal gland size of attendant bees
In order to determine whether caged queens received sufficient food, the HPG size of attending worker bees was measured. To obtain worker bees of defined age, single frames of late stage capped brood were brought to the laboratory and incubated in the dark at 33 °C (humidity provided by open water jars). The frames with worker brood were collected from two institute colonies, which were regularly inspected for symptoms of diseases. Newly emerged bees (≤24 h) were collected, colour marked and returned to the colony of origin. On day seven after emergence, the marked bees were re-collected and placed into the cages containing the queens. After seven days of exposure to neonicotinoids in the laboratory, the workers were immobilized on ice and the HPGs were dissected in ice cold phosphate buffered saline (PBS, pH 7.4). The specimens were fixed in formaldehyde (4% in PBS, Carl Roth), rinsed three times in PBS and mounted in Aquapolymount (Polysciences, Eppelheim; Germany). Three pictures of each gland (only one gland per bee) were photographed using a Leica phase contrast/fluorescence microscope and image capturing software (Leica, LASV4.4, Wetzlar, Germany). To measure the size of the glands, the diameter of 15 acini per bee was measured with ImageJ software (ImageJ 1.490; Image Processing and Analysis in Java, http://rsb.info.nih.gov/ij/index.html)75. The experiment was repeated three times with 20 to 33 individuals per treatment group (for details see Supplementary Table S2).
Statistical methods
Total hemocyte counts, melanisation/mean grey values, and mean diameters of inhibition zones were not normally distributed, and hence non-parametric statistics were used. Each immunocompetence measure was compared between groups treated with neonicotinoids and untreated control queens using KWT followed by post-hoc pair wise comparisons with Mann–Whitney U tests (MWU). Proportions of cell types were arcsin-transformed before performing statistical testing. The probability levels inferior to 0.05 were corrected for multiple testing according to the Holm’s sequential Bonferroni procedure76. All statistical tests were run with the computer program SPSS for Windows (v. 20).
References
Winston, M. L. The Biology of the Honey Bee. (Harvard University Press, 1987).
Pettis, J. S., Rice, N., Joselow, K., vanEngelsdorp, D. & Chaimanee, V. Colony Failure Linked to Low Sperm Viability in Honey Bee (Apis mellifera) Queens and an Exploration of Potential Causative Factors. PloS one 11, e0147220, doi:10.1371/journal.pone.0147220 (2016).
vanEngelsdorp, D., Tarpy, D. R., Lengerich, E. J. & Pettis, J. S. Idiopathic brood disease syndrome and queen events as precursors of colony mortality in migratory beekeeping operations in the eastern United States. Prev Vet Med 108, 225–233, doi:10.1016/j.prevetmed.2012.08.004.pmid:22939774 (2013).
Genersch, E. et al. The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies*. Apidologie 41, 332–352 (2010).
Williams, G. R. et al. Neonicotinoid pesticides severely affect honey bee queens. Scientific reports 5, 14621, doi:10.1038/srep14621 (2015).
Sandrock, C. et al. Impact of chronic neonicotinoid exposure on honeybee colony performance and queen supersedure. PloS one 9, e103592, doi:10.1371/journal.pone.0103592 (2014).
Lee, K. V. et al. A national survey of managed honey bee 2013–2014 annual colony losses in the USA. Apidologie 46, 292–305 (2015).
Alaux, C. et al. Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environmental microbiology 12, 774–782, doi:10.1111/j.1462-2920.2009.02123.x (2010).
Brodschneider, R. & Crailsheim, K. Nutrition and health in honey bees. Apidologie 41, 278–294, doi:10.1051/apido/2010012 (2010).
Di Pasquale, G. et al. Influence of pollen nutrition on honey bee health: do pollen quality and diversity matter? PloS one 8, e72016, doi:10.1371/journal.pone.0072016 (2013).
Sanchez-Bayo, F. et al. Are bee diseases linked to pesticides? - A brief review. Environment international 89-90, 7–11, doi:10.1016/j.envint.2016.01.009 (2016).
Elbert, A., Haas, M., Springer, B., Thielert, W. & Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest management science 64, 1099–1105, doi:10.1002/ps.1616 (2008).
Mullin, C. A. et al. High Levels of Miticides and Agrochemicals in North American Apiaries: Implications for Honey Bee Health. PloS one 5, e9754, doi:10.1371/journal.pone.0009754 (2010).
Jeschke, P., Nauen, R., Schindler, M. & Elbert, A. Overview of the status and global strategy for neonicotinoids. Journal of agricultural and food chemistry 59, 2897–2908, doi:10.1021/jf101303g (2011).
van der Sluijs, J. P. et al. Neonicotinoids, bee disorders and the sustainability of pollinator services. Current Opinion in Environmental Sustainability 5, 293–305, doi:10.1016/j.cosust.2013.05.007 (2013).
Goulson, D., Nicholls, E., Botias, C. & Rotheray, E. L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, 1255957, doi:10.1126/science.1255957 (2015).
Henry, M. et al. Reconciling laboratory and field assessments of neonicotinoid toxicity to honeybees. Proceedings. Biological sciences/The Royal Society 282, doi:10.1098/rspb.2015.2110 (2015).
Henry, M. et al. A common pesticide decreases foraging success and survival in honey bees. Science 336, 348–350, doi:10.1126/science.1215039 (2012).
Desneux, N., Decourtye, A. & Delpuech, J. M. The sublethal effects of pesticides on beneficial arthropods. Annual review of entomology 52, 81–106, doi:10.1146/annurev.ento.52.110405.091440 (2007).
Goulson, D. Review: An overview of the environmental risks posed by neonicotinoid insecticides. Journal of Applied Ecology 50, 977–987, doi:10.1111/1365-2664.12111 (2013).
Pisa, L. W. et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environmental science and pollution research international 22, 68–102, doi:10.1007/s11356-014-3471-x (2015).
Vanbergen, A. J. & Initiative, T. I. P. Threats to an ecosystem service: pressures on pollinators. Frontiers in Ecology and the Environment 11, 251–259, doi:10.1890/120126 (2013).
Matsuda, K. et al. Neonicotinoids: insecticides acting on insect nicotinic acetylcholine receptors. Trends in pharmacological sciences 22, 573–580 (2001).
Tomizawa, M. & Casida, J. E. Unique neonicotinoid binding conformations conferring selective receptor interactions. Journal of agricultural and food chemistry 59, 2825–2828, doi:10.1021/jf1019455 (2011).
Blacquiere, T., Smagghe, G., van Gestel, C. A. & Mommaerts, V. Neonicotinoids in bees: a review on concentrations, side-effects and risk assessment. Ecotoxicology 21, 973–992, doi:10.1007/s10646-012-0863-x (2012).
Suchail, S., De Sousa, G., Rahmani, R. & Belzunces, L. P. In vivo distribution and metabolisation of 14C-imidacloprid in different compartments of Apis mellifera L. Pest management science 60, 1056–1062, doi:10.1002/ps.895 (2004).
Rundlof, M. et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 521, 77–80, doi:10.1038/nature14420 (2015).
Whitehorn, P. R., O’Connor, S., Wackers, F. L. & Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 336, 351–352, doi:10.1126/science.1215025 (2012).
Straub, L. et al. Neonicotinoid insecticides can serve as inadvertent insect contraceptives. Proceedings of the Royal Society B: Biological Sciences 283, doi:10.1098/rspb.2016.0506 (2016).
Kairo, G. et al. Drone exposure to the systemic insecticide Fipronil indirectly impairs queen reproductive potential. Scientific reports 6, 31904, doi:10.1038/srep31904 (2016).
European Commission. Commission implementing Regulation (EU) No 485/2013 of 24 May 2013 amending Implementing Regulation (EU) No 540/2011, as regards the condition of approval of the active substances clothianidin, thiamethoxam and imidacloprid, and prohibiting the use and sale of seeds treated with plant protection products containing those active substances. Official Journal of the European Union, L139/12 25.5.2013 (2013).
Rosenkranz, P. et al. Schlussbericht: Deutsches Bienenmonitoring – DeBiMo 2011–2013 (2014).
Wilson-Rich, N., Dres, S. T. & Starks, P. T. The ontogeny of immunity: development of innate immune strength in the honey bee (Apis mellifera). Journal of insect physiology 54, 1392–1399, doi:10.1016/j.jinsphys.2008.07.016 (2008).
Mao, W., Schuler, M. A. & Berenbaum, M. R. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proceedings of the National Academy of Sciences 110, 8842–8846, doi:10.1073/pnas.1303884110 (2013).
Frias, B., Barbosa, C. & Lourenço, A. Pollen nutrition in honey bees (Apis mellifera): impact on adult health. Apidologie, 1–11, doi:10.1007/s13592-015-0373-y (2015).
Negri, P. et al. Cellular immunity in Apis mellifera: studying hemocytes brings light about bees skills to confront threats. Apidologie, 1–10 (2015).
Steinmann, N., Corona, M., Neumann, P. & Dainat, B. Overwintering Is Associated with Reduced Expression of Immune Genes and Higher Susceptibility to Virus Infection in Honey Bees. PloS one 10, e0129956, doi:10.1371/journal.pone.0129956 (2015).
Anderson, D. L. & Trueman, J. W. Varroa jacobsoni (Acari: Varroidae) is more than one species. Exp Appl Acarol 24, 165–189 (2000).
Guseman, A. J. et al. Multi-Drug Resistance Transporters and a Mechanism-Based Strategy for Assessing Risks of Pesticide Combinations to Honey Bees. PloS one 11, e0148242, doi:10.1371/journal.pone.0148242 (2016).
Dively, G. P., Embrey, M. S., Kamel, A., Hawthorne, D. J. & Pettis, J. S. Assessment of chronic sublethal effects of imidacloprid on honey bee colony health. PloS one 10, e0118748, doi:10.1371/journal.pone.0118748 (2015).
Di Prisco, G. et al. A mutualistic symbiosis between a parasitic mite and a pathogenic virus undermines honey bee immunity and health. Proceedings of the National Academy of Sciences of the United States of America 113, 3203–3208, doi:10.1073/pnas.1523515113 (2016).
James, R. R. & Xu, J. Mechanisms by which pesticides affect insect immunity. Journal of invertebrate pathology 109, 175–182, doi:10.1016/j.jip.2011.12.005 (2012).
Aufauvre, J. et al. Parasite-insecticide interactions: a case study of Nosema ceranae and fipronil synergy on honeybee. Scientific reports 2, 326, doi:10.1038/srep00326 (2012).
Di Prisco, G. et al. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proceedings of the National Academy of Sciences of the United States of America 110, 18466–18471, doi:10.1073/pnas.1314923110 (2013).
Pettis, J. S., vanEngelsdorp, D., Johnson, J. & Dively, G. Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Die Naturwissenschaften 99, 153–158, doi:10.1007/s00114-011-0881-1 (2012).
Doublet, V., Labarussias, M., de Miranda, J. R., Moritz, R. F. & Paxton, R. J. Bees under stress: sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environmental microbiology, doi:10.1111/1462-2920.12426 (2014).
Brandt, A., Gorenflo, A., Siede, R., Meixner, M. & Büchler, R. The Neonicotinoids Thiacloprid, Imidacloprid and Clothianidin affect the immunocompetence of Honey Bees (Apis mellifera L.). Journal of insect physiology 86, 40–47 (2016).
Schmid, M. R., Brockmann, A., Pirk, C. W., Stanley, D. W. & Tautz, J. Adult honeybees (Apis mellifera L.) abandon hemocytic, but not phenoloxidase-based immunity. Journal of insect physiology 54, 439–444 (2008).
Gatschenberger, H., Azzami, K., Tautz, J. & Beier, H. Antibacterial immune competence of honey bees (Apis mellifera) is adapted to different life stages and environmental risks. PloS one 8, e66415, doi:10.1371/journal.pone.0066415 (2013).
Negri, P., Maggi, M., Szawarski, N., Lamattina, L. & Eguaras, M. Apis mellifera haemocytes in-vitro, What type of cells are they? Functional analysis before and after pupal metamorphosis. Journal of Apicultural Research 53, 576–589, doi:10.3896/ibra.1.53.5.11 (2014).
Evans, J. D. et al. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect molecular biology 15, 645–656, doi:10.1111/j.1365-2583.2006.00682.x (2006).
Graaf, D. Cd, Dauwe, R., Walravens, K. & Jacobs, F. J. Flow cytometric analysis of lectin-stained haemocytes of the honeybee (Apis mellifera). Apidologie 33, 571–579 (2002).
Marringa, W. J., Krueger, M. J., Burritt, N. L. & Burritt, J. B. Honey Bee Hemocyte Profiling by Flow Cytometry. PloS one 9, e108486, doi:10.1371/journal.pone.0108486 (2014).
Negri, P. et al. Apis mellifera hemocytes generate increased amounts of nitric oxide in response to wounding/encapsulation. Apidologie 45, 610–617, doi:10.1007/s13592-014-0279-0 (2014).
Strand, M. R. The insect cellular immune response. Insect Science 15, 1–14, doi:10.1111/j.1744-7917.2008.00183.x (2008).
Sanchez-Bayo, F. & Goka, K. Pesticide Residues and Bees – A Risk Assessment. PloS one 9, e94482, doi:10.1371/journal.pone.0094482 (2014).
Haydak, M. H. Honey Bee Nutrition. Annual review of entomology 15, 143–156, doi:10.1146/annurev.en.15.010170.001043 (1970).
Davies, A. R. & Shuel, R. W. Distribution of C1-labelled carbofuran and dimethoate in royal jelly, queen larvae and nurse honeybees. Apidiologie 19, 37–50 (1988).
Giroud, B., Vauchez, A., Vulliet, E., Wiest, L. & Bulete, A. Trace level determination of pyrethroid and neonicotinoid insecticides in beebread using acetonitrile-based extraction followed by analysis with ultra-high-performance liquid chromatography-tandem mass spectrometry. Journal of chromatography. A 1316, 53–61, doi:10.1016/j.chroma.2013.09.088 (2013).
Škerl, M. I. S. & Gregorc, A. Heat shock proteins and cell death in situ localisation in hypopharyngeal glands of honeybee (Apis mellifera carnica) workers after imidacloprid or coumaphos treatment. Apidologie 41, 73–86, doi:10.1051/apido/2009051 (2010).
Hatjina, F. et al. Sublethal doses of imidacloprid decreased size of hypopharyngeal glands and respiratory rhythm of honeybees in vivo. Apidologie 44, 467–480, doi:10.1007/s13592-013-0199-4 (2014)
Amiri, E., Meixner, M., Büchler, R. & Kryger, P. Chronic Bee Paralysis Virus in Honeybee Queens: Evaluating Susceptibility and Infection Routes. Viruses 6, 1188–1201, doi:10.3390/v6031188 (2014).
Rondeau, G. et al. Delayed and time-cumulative toxicity of imidacloprid in bees, ants and termites. Scientific reports 4, 5566, doi:10.1038/srep05566 (2014).
Chen, Y. P., Pettis, J. S., Collins, A. & Feldlaufer, M. F. Prevalence and transmission of honeybee viruses. Applied and environmental microbiology 72, 606–611, doi:10.1128/aem.72.1.606-611.2006 (2006).
Ravoet, J., De Smet, L., Wenseleers, T. & de Graaf, D. C. Vertical transmission of honey bee viruses in a Belgian queen breeding program. BMC Veterinary Research 11, 61, doi:10.1186/s12917-015-0386-9 (2015).
Hernandez Lopez, J., Schuehly, W., Crailsheim, K. & Riessberger-Galle, U. Trans-generational immune priming in honeybees. Proceedings. Biological sciences/The Royal Society 281, 20140454, doi:10.1098/rspb.2014.0454 (2014).
Salmela, H., Amdam, G. V. & Freitak, D. Transfer of Immunity from Mother to Offspring Is Mediated via Egg-Yolk Protein Vitellogenin. PLoS pathogens 11, e1005015, doi:10.1371/journal.ppat.1005015 (2015).
Amiri, E., Meixner, M. D. & Kryger, P. Deformed wing virus can be transmitted during natural mating in honey bees and infect the queens. Scientific reports 6, 33065, doi:10.1038/srep33065 (2016).
Reetz, J. E. et al. Uptake of Neonicotinoid Insecticides by Water-Foraging Honey Bees (Hymenoptera: Apidae)Through Guttation Fluid of Winter Oilseed Rape. J Econ Entomol 109, 31–40, doi:10.1093/jee/tov287 (2016).
Alburaki, M. et al. Performance of honeybee colonies located in neonicotinoid, treated and untreated cornfields in Quebec. Journal of Applied Entomology, doi:10.1111/jen.12336 (2016).
European Food and Safety Authority (EFSA). Guidance on the risk assessment of plant protection products on bees (Apis mellifera, Bombus spp. and solitary bees). EFSA Journal 11, 3295 (2013).
Büchler, R. et al. Standard methods for rearing and selection of Apis mellifera queens. Journal of Apicultural Research 52, 1–30, doi:10.3896/ibra.1.52.1.07 (2013).
OECD. Proposal for a new guideline for the testing of chemicals. Honey bee (Apis mellifera L.), chronic oral toxicity test, 10 day feeding test in the laboratory (2016).
Allander, K. & Schmid-Hempel, P. Immune defence reaction in bumble-bee workers after a previous challenge and parasitic coinfection. Functional Ecology 14, 711–717, doi:10.1046/j.1365-2435.2000.00476.x (2000).
Rasband, W. S. ImageJ. U. S. National Institutes of Health, Bethesda, Maryland, USA, http://imagej.nih.gov/ij/ (1997–2016).
Abdi, H. Holm’s sequential Bonferroni procedure. Encyclopedia of research design 1 (2010).
Acknowledgements
We gratefully thank W. Strasser for queen rearing, K. Petzold-Treibert for assistance in beekeeping, S. Backhaus, A. Gorenflo, B. Hohnheiser, and Ch. Geßner for technical support, D.T. Brandt for histochemical stainings, and all team members of the Bee Institute Kirchhain for support and fruitful discussions. This research was supported by the EU and Land Hessen “Förderung von Maßnahmen zur Verbesserung der Erzeugung und Vermarktung von Honig in Hessen”.
Author information
Authors and Affiliations
Contributions
A.B. and R.S. designed the study and performed the experiments. K.G. made microscope pictures. A.B. and R.S. analysed the data and wrote the manuscript. All authors edited and approved the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brandt, A., Grikscheit, K., Siede, R. et al. Immunosuppression in Honeybee Queens by the Neonicotinoids Thiacloprid and Clothianidin. Sci Rep 7, 4673 (2017). https://doi.org/10.1038/s41598-017-04734-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-04734-1
This article is cited by
-
Acute toxicity of sublethal concentrations of thiacloprid and clothianidin to immune response and oxidative status of honey bees
Apidologie (2022)
-
Varying impact of neonicotinoid insecticide and acute bee paralysis virus across castes and colonies of black garden ants, Lasius niger (Hymenoptera: Formicidae)
Scientific Reports (2021)
-
Exposure to low doses of pesticides induces an immune response and the production of nitric oxide in honeybees
Scientific Reports (2021)
-
Effects of developmental exposure to pesticides in wax and pollen on honey bee (Apis mellifera) queen reproductive phenotypes
Scientific Reports (2021)
-
Honeybee colonies compensate for pesticide-induced effects on royal jelly composition and brood survival with increased brood production
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
