
Abstract
Cercospora leaf spot (CLS), caused by Cercospora beticola, is a major disease of Beta vulgaris worldwide. No sexual stage is known for C. beticola but in its asexual form it overwinters on infected plant debris as pseudostromata, and travels short distances by rain splash-dispersed conidiospores. Cercospora beticola infects a broad range of host species and may be seedborne. The relative contribution of these inoculum sources to CLS epidemics on table beet is not well understood. Pathogen isolates collected from table beet, Swiss chard and common lambsquarters in mixed-cropping farms and monoculture fields in New York and Hawaii, USA, were genotyped (n = 600) using 12 microsatellite markers. All isolates from CLS symptoms on lambsquarters were identified as C. chenopodii. Sympatric populations of C. beticola derived from Swiss chard and table beet were not genetically differentiated. Results suggested that local (within field) inoculum sources may be responsible for the initiation of CLS epidemics in mixed-cropping farms, whereas external sources of inoculum may be contributing to CLS epidemics in the monoculture fields in New York. New multiplex PCR assays were developed for mating-type determination for C. beticola. Implications of these findings for disease management are discussed.
Similar content being viewed by others

Introduction
Population genetic studies of plant pathogens using selectively neutral genetic markers have made significant contributions to plant disease management in the past few decades1, 2. These studies apply knowledge of the genetic composition and patterns of genetic diversity in pathogen populations to test hypotheses regarding their biology and plant disease epidemiology3,4,5. Population genetics has been used to make inferences about the origin6,7,8,9, sources of inoculum10, 11, routes of introduction12, 13, migration pathways7, 14,15,16,17 and reproductive biology18, 19 of plant pathogens. Knowledge of the reproductive modes of plant pathogens is critical to disease management, due to its significant impacts on the evolutionary trajectory and epidemiology of pathogen populations. Recombination as a result of sexual reproduction is an influential evolutionary force that generates potentially adaptive genotypic diversity, purges deleterious mutations, and produces sexual propagules with enhanced survival and dispersal20. In the absence of a known sexual stage for a plant pathogen, population genetic studies can provide indirect evidence to confirm the asexuality of pathogen populations21,22,23, or reveal historical or potentially cryptic sex in presumably asexual fungi18, 19, 24. In pathogen populations with frequent sexual reproduction, high genotypic diversity, random association of selectively neutral genetic loci and equal distribution of the opposite mating-types (for heterothallic fungi) are expected. Alternatively, a strictly asexual population is characterized by low genotypic diversity, lack of recombinant genotypes, widespread occurrence of a few clonal lineages, non-random association of unlinked loci, and skewed distribution of alternate mating-types in the population20.
The presence of a sexual form remains unknown for the globally and economically important plant pathogen, Cercospora beticola, the cause of Cercospora leaf spot (CLS) of Beta vulgaris L. (sugar beet, table beet and Swiss chard)25. Despite the lack of a known sexual form, C. beticola is considered to be a heterothallic, ascomycete fungus due to the discovery of two alternate mating-type genes (MAT1-1-1 and MAT1-2-1) at equal frequency in parts of Europe26, 27. Population genetic studies have also suggested the potential presence of a sexual cycle for C. beticola in sugar beet production areas in North Dakota28. In contrast, no evidence for sexual reproduction has been found in other parts of the USA29 or the Middle East30.
In New York, CLS epidemics impose significant losses to both the fresh market and processing table beet (Beta vulgaris ssp. vulgaris) industries25. Symptoms include light brown to grey leaf spots, usually with a red to purple margin in red beet varieties, which expand into necrotic lesions that result in premature defoliation31. Conidiospores are mainly disseminated through water splash over short distances, resulting in multiple infection cycles within a season and polycyclic epidemics25.
Infested plant debris is considered to be a major source of inoculum initiating CLS epidemics32,33,34 as C. beticola in the form of mycelia and pseudostromata may persist for a maximum of 22 months35 to three years36 depending on climatic conditions. In small-scale, mixed-cropping farms in New York, table beet and Swiss chard (B. vulgaris ssp. cicla) are often grown in close proximity along with other vegetables, on an annual basis. It is, therefore, plausible that infested plant debris from previous years may be a major source of inoculum contributing to the annual CLS epidemics on susceptible hosts in these farms. In contrast, large-scale monoculture table beet fields in New York involve rotations with non-host crops in the same field for a recommended minimum duration of three years, which aims to reduce the persistence of inoculum in soil and on infested plant debris. Nevertheless, CLS epidemics are prevalent and result in substantial losses in these fields, even with rotations lasting up to five years37, and even in the fields that have never grown table beet (George Abawi, personal communication). The major sources of initial inoculum for CLS epidemics in monoculture table beet fields in New York are therefore largely unknown.
Understanding the relative contribution of inoculum sources to CLS epidemics is essential for the design of effective management strategies. If C. beticola populations in New York are capable of sexual reproduction, overwintering sexual structures may serve as primary inoculum for newly established table beet fields within several kilometres. Cercospora beticola is also known to infect a broad range of plants from various families32, 38. Therefore, infected weeds (especially from Amaranthaceae) are believed to serve as green bridges for C. beticola inocula during crop rotations. For example, common lambsquarters (Chenopodium album) is a susceptible weed species that is frequently found in New York table beet fields and often has CLS symptoms on the leaves. Other potential sources of inoculum include asexual spores carried by wind35, 39 or insects38, 39 from neighbouring table beet or Swiss chard productions. Seedborne inoculum32 has also been suggested to contribute to CLS epidemics in some European countries40, 41 but its contribution to CLS epidemics in New York has not been investigated.
If the primary inoculum is introduced from a source outside a field of interest, such as infested seeds or windborne ascospores, we would expect to see a panmictic population with low population structuring due to frequent migration among fields. In fungal populations with long-distance dispersal of ascospores, a pattern of isolation by distance (IBD) is usually detected, resulting from the gradual spread of the disease from an original source of inoculum, hence, resulting in an increase in genetic differentiation among populations as distance increases42, 43. Alternatively, if a local source of inoculum, e.g., locally-developed sexual or asexual structures or infected weeds, are the major source of primary inoculum, genetic structuring of C. beticola populations would be present.
Potential survival of C. beticola on weeds or other crops could have profound impacts on CLS epidemiology as it would promote genetic diversity by reducing the impact of genetic drift through increasing the effective pathogen population size. Moreover, this would reduce the amount of disruptive selection pressure that the pathogen would have undergone by switching from a pathogenic to saprophytic mode.
If C. beticola inoculum surviving on local weeds or other susceptible crops (e.g. Swiss chard) is a major contributor to CLS epidemics on table beet, we would expect sympatric populations isolated from these alternate hosts to be genetically undifferentiated from C. beticola populations on table beet. To test this hypothesis, we examined Cercospora spp. populations from symptomatic lambsquarters growing sympatrically with table beet. Also, we genotyped C. beticola populations from Swiss chard and table beet plants growing in sympatry in mixed-cropping farms and gardens in New York and Hawaii, to investigate whether diseased Swiss chard may be a source of primary inoculum initiating CLS epidemics on table beet.
The objectives of this study were to i) investigate the potential for sexual reproduction in C. beticola populations in New York; ii) test the hypothesis that inoculum from lambsquarters and Swiss chard may contribute to CLS epidemics on table beet; and iii) determine the distribution of genetic diversity in C. beticola populations within and among mixed-cropping and monoculture table beet production to understand the relative contributions of local or external sources of inoculum to CLS epidemics. Herein, the term ‘population’ is used to refer to a group of isolates defined under certain geographical or host criteria (e.g., collected from one field/state or from the same host/variety).
Results
Cercospora beticola isolation, identification and genotyping
In New York, 649 C. beticola isolates were obtained from 426 table beet and 14 Swiss chard plants collected from two mixed-cropping farms (Farms 1 and 2) and three monoculture table beet fields (Fields 3, 4 and 5), in 2015 (Fig. 1). Of these, 422 isolates were randomly selected for microsatellite genotyping and mating-type determination (Table 1). In Hawaii, 178 isolates were obtained from 34 and 65 table beet and Swiss chard plants, respectively, all of which were included for genotyping and mating-type determination (Table 1). Twenty-eight Cercospora isolates were obtained from symptomatic lambsquarters plants inter-mixed with table beet in Field 5. Morphological and cultural features of these isolates were distinct from C. beticola as conidiospores were shorter, subcylindrical in shape, straight to slightly curved; and also colonies on potato dextrose agar (PDA) were slow growing (~10 mm diameter after 14 days). Further multi locus sequence typing (ITS, actin, histone H3, translation elongation factor 1-α, and calmodulin) showed that these isolates belonged to C. chenopodii (data not shown), hence, were excluded from subsequent analyses. Cercospora beticola-specific primers (CercoCal-beta and CercoCal-R) resulted in a ~170 bp product in all C. beticola isolates from table beet and Swiss chard as well as C. chenopodii isolates from lambsquarters. This demonstrated that the primers CercoCal-beta and CercoCal-R were not specific to C. beticola. On the other hand, none of the microsatellite loci amplified a band in C. chenopodii (data not shown). Re-genotyping 25% of isolates detected no genotyping error except for one primer pair (CbSSR3), which had an error rate of 0.01.

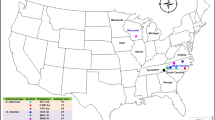
Sampling locations for Cercospora beticola populations collected in 2015 from (A) New York and (B) Hawaii. Numbers 1, 2, 3, 4, and 5 in New York represent Farm 1, Farm 2, Field 3, Field 4 and Field 5, respectively. In Hawaii, MCG, UH and DH represent community gardens in Honolulu (MCG = Manoa community garden, UH = University of Hawaii organic garden, and DH = Diamond Head community garden). The map presented here are modified from https://commons.wikimedia.org/wiki/File:Map_of_New_York_County_Outlines.svg and https://upload.wikimedia.org/wikipedia/commons/e/e5/Interstate_H1_map.png. Figures were produced in Microsoft Paint and Microsoft PowerPoint 2013.
Within-plant and within-lesion genotypic diversity
Two to four distinct MLGs were obtained from six of the 12 intensively sampled plants. Isolations from multiple lesions on single leaves detected two to four distinct MLGs in 12 of 21 leaves (57%), which, in 50% of the leaves, also belonged to different mating-types. Up to three distinct MLGs, showing variation at two to 10 loci, were associated with single CLS lesions on table beet and Swiss chard plants. In one lesion, distinct MLGs belonged to different mating-types.
Lack of host association in table beet and Swiss chard populations
At Farm 2, sympatric C. beticola populations collected from two varieties of table beet (‘Detroit’ and ‘Touchstone Gold’) were not genetically differentiated (ΦST = 0.002, P = 0.276; D = 0.007, 95% CI: 0.000–0.016; \({{\rm{G}}}_{\mathrm{ST}}^{^{\prime} }\) = −0.0134, CI: −0.020–−0.009). Moreover, no significant difference was detected in the genotypic diversity (P > 0.29) or genotypic composition (P = 0.19). From 20 recurrent MLGs at Farm 2, 13 were shared between these two table beet varieties. Likewise, sympatric populations from table beet and Swiss chard did not show significant population differentiation (Table 2). This was also demonstrated from the UPGMA dendrogram computed based on Nei’s distance (Fig. 2), in which sympatric populations from table beet and Swiss chard mostly grouped closer together, but more distant from populations from other fields. Therefore, for subsequent analyses, isolates from different hosts in one farm/garden were pooled into one population representing the corresponding farm/garden.

UPGMA dendrogram constructed based on Nei’s distance among Cercospora beticola populations from New York and Hawaii (MCG = Manoa community garden, UH = University of Hawaii organic garden, and DH = Diamond Head community garden), 2015, represented by state–field–host (TB = table beet and CH = Swiss chard).
Indices of genetic diversity
The number of alleles in the entire C. beticola population ranged from 2 (CbSSR2) to 24 (CbSSR3) with an average of 6.75 alleles per locus. Nei’s unbiased gene diversity (He) per locus ranged from 0.034 (CbSSR1) to 0.87 (CbSSR3). In New York, locus CbSSR24 was monomorphic in Farm 1, while CbSSR1 was monomorphic in the other four locations. All loci were polymorphic for C. beticola populations from Hawaii (Table 3). Overall, allelic richness; private allelic richness; and allelic and genotypic diversity after rarefaction was higher in the C. beticola population from New York compared to Hawaii (Table 3).
Multi-locus genotype analyses
In the 359 C. beticola isolates from New York, 130 unique MLGs were detected, resulting in 64% clonality of the population. Despite the relative high level of clonality in New York table beet fields, genotypic diversity was also high (λ ≥ 0.90) at all locations except for Farm 1, which had a moderate genotypic diversity (λ = 0.57). Each field contained a different dominant MLG (Fig. 3). For example, at Farm 1, MLG34 occurred more than 20 times in the population, which also had low genotypic evenness (Table 3). Of the 130 MLGs detected in New York, 60 MLGs were recurrent (occurred at least twice), and 14 were shared among fields (Fig. 4). Field 3 had the highest number of shared MLGs (12) while only one MLG from fields Farm 1 and 2 were present in another population (shared with Field 3).

Recurrent multi-locus genotypes and their frequency in Cercospora beticola populations from New York (Farm 1, Farm 2, Field 3, Field 4 and Field 5) and Hawaii (MCG = Manoa community garden, UH = University of Hawaii organic garden, and DH = Diamond Head community garden), 2015.

Discriminant Analysis of Principal Components for Cercospora beticola populations from New York (Farm 1, Farm 2, Field 3, Field 4 and Field 5) and Hawaii (MCG = Manoa community garden, UH = University of Hawaii organic garden, and DH = Diamond Head community garden), 2015.
In Hawaii, only 16 unique MLGs were detected among the 145 isolates, hence the 90% clonal fraction of C. beticola population (Table 3), which suggested the presence of a few dominant MLGs in the Hawaiian population (Fig. 3). Five MLGs were shared among locations in Hawaii, three of which occurred in all gardens. Only three MLGs were shared between New York and Hawaii, all of which occurred at Farm 2 (Fig. 3).
Farm 2 contained 20 recurrent MLGs, five of which occurred on both table beet and Swiss chard. In Hawaii, of the five recurrent MLGs in DH and MCG, three MLGs existed on both table beet and Swiss chard (Fig. S1). MLG245 occurred once in New York on table beet and once in Hawaii on Swiss chard.
For the 60 and 16 recurrent MLGs in New York and Hawaii, respectively, the probability of the MLGs having originated from independent sexual events was greater than 0.05 for only two and three MLGs (MLG80 and MLG207 for New York; and MLG10, MLG13 and MLG16 for Hawaii), confirming the clonal origin of the majority of the overrepresented MLGs.
Population structure and differentiation
AMOVA analysis of the entire population found 25% of the total observed genetic diversity was partitioned among the fields within states (P = 0.001), 65% among individuals within fields (P = 0.001), and 10% between the two states (P = 0.15). Similar results were obtained through AMOVA analysis using RST, where 20 and 80% of the total diversity existed among and within fields, respectively (P < 0.001), and none between New York and Hawaii (P = 1.00).
The pairwise index of differentiation ΦPT showed significant differentiation among the five fields within New York (P < 0.013) except for Fields 3 and 4, which were not significantly differentiated (P = 0.715) (Table 4). The three monoculture fields (Fields 3, 4 and 5) showed low population differentiation (ΦPT < 0.042, P < 0.05) while the two mixed-cropping farms (Farm 1 and 2) showed greater differentiation from each other (ΦPT = 0.240, P < 0.05) and from the monoculture fields (ΦPT > 0.137, P < 0.05) (Table 4).
Discriminant Analysis of Principal Components (DAPC) using fields as pre-defined populations also demonstrated that populations from the three monoculture table beet fields overlapped while those from mixed-cropping farms clustered separately (Fig. 4). The Mantel test detected significant association between the genetic and geographical distance of the five C. beticola populations in New York for the entire dataset (R 2 = 0.60, P = 0.039) but not for the clone-corrected data set (R 2 = 0.48, P = 0.052) (Fig. 5).

Standardized pairwise genetic distance (ΦPT/(1 − ΦPT)) plotted against geographic distance among five Cercospora beticola populations in New York, 2015. The open squares and dashed line are based on the complete data set, and the filled circles and solid line are based on the clone-corrected data set.
Population structure analysis with no a priori population assignment through Bayesian clustering analysis of both the clone-corrected and non-clone-corrected datasets resulted in detection of two distinct clusters in the population (Fig. S2). Likewise, a dendrogram produced based on Bruvo’s distance (Fig. 6) grouped the C. beticola isolates into two major clades. Two hundred and twenty-nine isolates were assigned to cluster 1 and 275 isolates were assigned to the remaining cluster. These two clusters did not correspond to sampling location or host (Fig. 6). DAPC analysis for the non-clone-corrected data set was inconclusive as the Bayesian Information Criterion (BIC) consistently decreased with the number of clusters until K reached the total number of MLGs. For the clone-corrected data set, however, an increase in BIC was observed with increasing number of K (Fig. S3). K = 6 was selected as the optimal number of clusters since the BIC value decreased by a negligible amount after K = 6 (Fig. S3). Three of these groups (1, 5 and 6) belonged to cluster 1 detected in STRUCTURE, group 1 and 4 belonged to cluster 2, while individuals in group 3 occurred in both the first and second clusters detected in STRUCTURE.

UPGMA dendrogram of Cercospora beticola isolates collected in 2015 from New York (Farm 1, Farm 2, Field 3, Field 4 and Field 5) and Hawaii (MCG = Manoa community garden, UH = University of Hawaii organic garden, and DH = Diamond Head community garden) based on Bruvo’s distance. Bootstrap support values greater than 50 are shown above the branches. The thick orange and blue branches denote the two clusters detected through Bayesian clustering method implemented in STRUCTURE. The colour of the isolates correspond to the field/community garden they belong as indicated in the right-hand-side legend. The colour of the outer ring corresponds to the six clusters detected by DAPC analysis as shown in the left-hand-side legend and Fig. S3.
Mating-type determination
Screening all isolates by multiplex PCR using primers CercosporaMat1F/R and 356/358 for mating-type determination (Table 5) resulted in multiple, non-specific PCR products in ~30% of the isolates, which confused mating-type determination (data not shown). The novel three-primer and four-primer mating-type assays developed based on non-exonic regions of the mating-type genes (Fig. 7; Table 5) successfully amplified a single band of the expected size in these isolates.

(A) Schematic alignment of the MAT1 idiomorphs in Cercospora beticola; MAT1-2-1 (top) and MAT1-1-1 (bottom), showing the position and orientation of the open reading frames with arrowed boxes, and position of introns with black lines. The position of the primers used for mating-type determination are shown with arrows bearing the primer number according to Table 5. The thick black and grey lines in the alignment represent differences and similarities, respectively. (B) Three-primer mating-type assay, resulting in PCR amplicons of ~1,700 and ~1,050 bp in MAT1-1 and MAT1-2 isolates, respectively. (C) Four-primer mating type assay, resulting in PCR amplicons of ~500 and ~700 bp in MAT1-1 and MAT1-2 isolates, respectively. The first and last line in each gel include 6 µL of O’GeneRuler 1 kb Plus DNA Ladder (Thermo Scientific).
Tests for random mating
Mating-type ratios
For the non-clone-corrected data set, mating-type frequencies deviated from a 1:1 ratio expected under random mating for all populations except for that derived from Farm 2 (Table 6). After clone-correction, MAT1-1 and MAT1-2 isolates were present in equal ratios in all fields except for 4 and 5 in New York (Table 6).
Linkage disequilibrium of microsatellite loci
Significant linkage of microsatellite loci was detected in all populations in New York and Hawaii even after clone-correction of datasets, rejecting the null hypothesis of random mating (Table 6).
Analyses of mating-type ratios and linkage disequilibrium of microsatellite loci conducted for the two clusters detected through Bayesian clustering implemented in STRUCTURE also rejected the null hypothesis of random mating (Table 7).
Test for recombination
The proportion of compatible loci (PrCP) was less than one in four sampling locations in New York and in two locations in Hawaii (Table 3). The observed PrCP values in all these locations were significantly larger than expected under the null hypothesis of random mating. Analysis of the phylogenetically compatible pairs of loci for the two clusters detected through Bayesian clustering implemented in STRUCTURE indicated that only 36 and 40% of the pairs of loci were compatible in cluster 1 and 2, respectively. For both clusters, the observed value of PrCP was significantly larger than expected under random mating (P < 0.001).
Discussion
To enhance our understanding of the epidemiology and sources of inoculum contributing to CLS epidemics on table beet, 12 microsatellite markers were used to characterise the genetic structure of C. beticola populations in contrasting table beet production systems in New York. Sympatric Cercospora spp. populations collected from table beet, Swiss chard and lambsquarters in New York and Hawaii were also studied to establish whether inoculum from alternative hosts contribute to CLS epidemics on table beet.
Cercospora beticola is known to infect a wide range of plants from various families32, 38, 44,45,46,47,48. Thus, infected weeds, especially from the family Amaranthaceae, are believed to serve as C. beticola inoculum reservoirs. One of these species is lambsquarters, which was found to be susceptible to C. beticola in glasshouse pathogenicity trials (data not shown). However, all Cercospora spp. collected from symptomatic lambsquarters plants growing in sympatry with table beet were identified as C. chenopodii. This is congruent with reports that studies in the Netherlands in 2003 also failed to isolate C. beticola from weeds49; and emphasises the need for more comprehensive surveys on a range of weeds from Amaranthaceae and other families to better understand the prevalence and incidence of C. beticola on hosts other than Beta vulgaris.
Sympatric C. beticola populations from different table beet varieties were not genetically differentiated based on microsatellite allele frequencies, nor did they differ in genotypic diversity or composition. Likewise, sympatric populations from Swiss chard and table beet in New York and Hawaii were not genetically differentiated. However, populations from Swiss chard and table beet from a mixed-cropping farm (Farm 2) in New York were significantly different in genotypic composition. Of the 20 recurrent MLGs in this farm, 13 were shared between the two table beet varieties while only five were shared between Swiss chard and table beet. This may be attributed to differences in virulence and/or aggressiveness of these MLGs on different hosts. For example, Melampsora larici-populina populations from cultivated and wild poplars showed little population differentiation based on neutral genetic markers while displaying different virulence profiles42. The lack of genetic differentiation of C. beticola populations from Swiss chard and table beet based on microsatellite allele frequencies is suggestive of lack of reproductive isolation between these populations. Thus, infected Swiss chard from nearby organic productions or home gardens may potentially serve as a source of inoculum for CLS epidemics in table beet fields.
In 50% of intensively sampled plants and leaves, multiple unique MLGs were detected on the same plant or leaf. Single lesions were also occasionally found to be associated with up to three unique MLGs. Co-infection of a single lesion with multiple MLGs has been reported in other pathosystems50, 51, including C. beticola on sugar beet in Europe41. Moretti et al. used Random Amplified Polymorphic DNA (RAPD) and Direct Amplification of Minisatellite region DNA (DAMD) to characterize 10 C. beticola isolates obtained from a single lesion on sugar beet, all of which showed distinct banding patterns41. As the arrangement of the mating-type genes in C. beticola indicates heterothallism26, 27, the co-occurrence of multiple MLGs in the same lesion may have profound impacts on population structure of this pathogen by enabling MLGs with alternate mating-types to sexually reproduce. Detection of two alternate mating-types in a single lesion in this study provides further evidence of the potential of C. beticola to undergo sexual reproduction.
High genotypic diversity, detection of admixed genotypes by Bayesian clustering and DAPC analyses (Figs S2 and S3), and lack of congruence between the clusters produced in UPGMA and DAPC analyses (Fig. 6), are all suggestive of recombination in the C. beticola population. Although the potential of rare parasexual events, somatic recombination or mitotic crossing over20, 52, 53 cannot be disregarded, the presence of mating-types in equal ratios is some table beet fields suggests that some C. beticola populations may be sexually reproducing.
On the other hand, overrepresentation of MLGs (high clonality), unequal frequency of mating-types in some fields, and linkage disequilibrium of microsatellite loci in all populations, even after clone correction, rejected the null hypothesis of random mating. Since other phenomena such as population structure/admixture can also result in linkage disequilibrium of loci20, tests of random mating were conducted on the two major clusters detected using Bayesian analyses (Table 7), which again rejected the null hypothesis of random mating. Severe bottlenecks and founder events, which occur frequently in agroecosystems, cause random changes in pathogen populations, which may result in linkage disequilibrium (LD) of loci54. Moreover, unequal frequency of mating-types and significant LD only reject the null hypothesis of random mating but do not prove the lack of sex as many fungal populations with the ability to sexually reproduce have been reported to have a clonal structure55, 56. Since most plant pathogenic fungi undergo clonal reproduction at some stage in their life cycles, deviations from random mating are expected and do not indicate lack of recombination54.
To test the hypothesis of strict clonality, rather than random mating, we tested the presence of phylogenetic incompatibility between pairs of loci. Under the assumption that recurrent, parallel and reverse mutations are highly improbable, phylogenetic incompatibility of loci may only arise by recombination54. Hence, the fact that 47, 65, 20 and 74% of pairs of loci were incompatible in Fields 3, 4, 5, and Farm 2 may be interpreted as presence of recombination in these populations. Although it is plausible that high levels of homoplasy, which is a known drawback of microsatellite loci57, contributed to the low rate of phylogenetic compatibility, the possibility of recombination cannot be disregarded.
Multiple studies have suggested that introduction of inoculum to initiate CLS epidemics occurs through rain splash dispersal of conidiospores from infected plant residues from previous years32, 33, 58. Survival of C. beticola inoculum on plant debris within soil has been reported in many studies to be at least 10 months35 to a maximum of three years36, depending on the climatic conditions. Planting sugar beet seed in infested soil was reported to result in infection of cotyledons and young seedlings33. Infection through the roots of mature plants has also been reported in sugar beet49, 59 but could not be reproduced in subsequent studies35. Wilting of beet leaves under hot conditions in the field occurs frequently, bringing the leaves in contact with the soil33. The moisture trapped under the lower surface of the leaves may contribute to leaf infection from C. beticola inoculum in the surface of the soil33. However, the monoculture table beet fields in New York follow rotation guidelines with three to five37 years of non-host crops between table beet. This infers that after five years, infected plant debris must be present with sporulation close to the soil surface. Further studies are required to quantify the long-term survival of C. beticola on plant residues, or in soil33 in New York.
Low genetic differentiation among the monoculture table beet fields (ΦPT ≤ 0.042), and lack of differentiation between Fields 3 and 4 was surprising. Fields 3 and 4 are more than six kilometres apart, and separated from Field 5 by at least 50 km. Considering the long rotation periods between table beet crops in processing, monoculture table beet fields, significant founder events leading to high genetic drift and stochastic differentiation of the C. beticola population may be expected60, unless external sources of inoculum dominate and homogenise these populations. In addition to the lack of genetic differentiation, Fields 3 and 4 also had eight MLGs in common, and each shared four MLGs with Field 5. Although this could be caused by the low power of the microsatellite loci to resolve these clones into distinct MLGs, distribution of these clones over processing fields can be indicative of genotype flow. CLS epidemics in fields where sugar beet had not been planted for 20 years in the Netherlands61 and table beet fields that had never been planted to table beet (George Abawi, personal communication), also suggest a means of long-distance dispersal for this pathogen.
A pattern of IBD is usually interpreted as the natural process of dispersal of windborne inoculum, resulting from the gradual spread of the disease from an original source of inoculum42. However, C. beticola conidiospores are believed to disseminate through rain splash32, 39, 58, and travel short distances (100 to 400 m) by wind32, 34, 62. Moreover, long-distance dispersal of ascospores, if present, cannot explain the dissemination of clones as C. beticola is heterothallic and potential sexual reproduction would not likely result in the production of clones. Plausible sources of long distance dispersal of the MLGs among the monoculture table beet fields are insects32, human movement of agricultural machinery or seed, the role of which in C. beticola dispersal requires further investigation. In this study, the seed for the three monoculture fields all originated from Mount Vernon, WA, USA.
Structuring of C. beticola populations from mixed-cropping farms in New York may be attributed to the dominant role of local sources of inoculum in initiating CLS epidemics in these fields. Populations from the two mixed-cropping farms in New York were highly differentiated from each other and also from the monoculture fields. Each of the two mixed-cropping farms contained a different dominant genotype (Fig. 3), and did not share genotypes with each other, indicative of limited dispersal between these fields. The seed for each of the mixed-cropping farms was obtained from different sources. Thus, if infested seed is a significant source of inoculum to CLS epidemics, the disparate population structure in the two mixed-cropping farms could have also been the result of seedborne inoculum, further emphasising the need for additional studies on the role of seed in CLS epidemics.
Future studies should quantify temporal changes in genotypic diversity and composition in mixed-cropping farms to enhance the understanding of the relative potential sources of inoculum to CLS epidemics. Moreover, population genomics studies and use of higher resolution molecular markers such as single nucleotide polymorphisms may have utility to improve our detection of recombination events or signs of sex in C. beticola populations in New York.
Methods
Sampling and isolation
Sampling of Cercospora beticola populations in New York was conducted in 2015 from two mixed-cropping farms (Farm 1 and Farm 2) and three monoculture table beet fields (Fields 3 and 4 = variety Ruby Queen; Field 5 = variety Red Ace) (Fig. 1). The mixed-cropping farms consisted of small-scale vegetable production producing a few rows of table beet, Swiss chard, and other fresh vegetables on an annual basis. Monoculture production consisted of large-scale (>0.2 km2) table beet fields with at least 3 years of a non-host crop prior to table beet cultivation.
Hierarchical sampling was conducted involving the arbitrary selection of sampling points at defined intervals along transects/rows, and sampling of multiple symptomatic plants within each location. The distance between the sampling points and number of plants sampled per location varied between the processing and fresh market production systems due to differences in the size of the fields. In the three monoculture fields, five sampling locations were selected at 20-m intervals along each of the two transects, which were separated by 50 m. Ten plants where arbitrarily sampled within a 5-m radius of each sampling location (n = 100 in each field). In addition, 28 symptomatic lambsquarters plants were collected from Field 5 to obtain a sympatric population of C. beticola. The mixed-cropping farms, each included only a few rows of table beet, which were intensively sampled at 5–10-m intervals along the rows. Farm 1 included three rows of table beet variety Detroit, from which 36 symptomatic plants were sampled. Farm 2 included three rows of table beet variety Touchstone Gold and one row of table beet variety Detroit as well as two rows of Swiss chard varieties Oriole and Ruby Red planted intermixed with each other. The incidence of CLS on the Swiss chard was substantially lower than on table beet at this location. Therefore, symptomatic leaves from 90 table beet and 14 Swiss chard plants were collected from Farm 2.
Sampling in Hawaii was conducted at two community gardens in Honolulu, and at the University of Hawaii at Manoa, in 2015 (Fig. 1). The community gardens consisted of small-scale plots of an average size of 10 × 15 m, planted to a range of vegetables in an intermixed manner. When Swiss chard or table beet plants with CLS symptoms were observed in these plots, 1–3 leaves per plant were sampled.
For all plants sampled in New York, only one C. beticola isolate per plant was obtained, except for 12 arbitrarily selected plants, from which multiple isolates were obtained from leaves of single plants, lesions on single leaves, and also from single lesions as described by Bolton et al.63. For Hawaiian samples, up to 10 leaves were arbitrarily chosen from each garden for intensive isolation, where multiple isolates were obtained from a single leaf, or a single lesion.
Cercospora beticola identification and genotyping
Genomic DNA was extracted from all fungal isolates as described previously37, and the species-specific PCR primers CercoCal-beta and CercoCal-R64 were used to confirm the identity of isolates as C. beticola. Genotyping was done for 600 isolates (178 isolates from Hawaii and 422 from New York) using 12 microsatellite markers (CbSSR1, CbSSR2, CbSSR3, CbSSR665, CbSSR20, CbSSR21, CbSSR22, CbSSR23, CbSSR24, CbSSR25, CbSSR26, and CbSSR2766) as described previously66. Alleles were scored using the Geneious Microsatellite Plugin v. 1.467. To reduce genotyping error, genotyping was replicated for 25% of the samples, including all isolates showing null or rare alleles, and error rates68 (el) were quantified for each locus. Genotyping was conducted a third time for samples that showed allelic variation in locus CbSSR3 between the two replications.
Microsatellite dataset refinement
Intensive isolation of multiple C. beticola strains from single leaves or lesions may result in re-isolating the same individual multiple times, which may skew the result of population genetic analyses. Therefore, the multi-locus genotypes (MLGs) for all isolates were determined in the R package poppr v.2.2.069, 70 to assess the occurrence of MLGs in single lesions and leaves. For subsequent genetic diversity and structure analyses, the data set was clone-corrected to the leaf level, i.e., identical MLGs isolated from a single lesion/leaf were identified and only one representative isolate from each leaf was retained for subsequent analyses. This resulted in 359 and 145 isolates from New York and Hawaii, respectively. In order to reduce the effect of genotyping error and missing data on genetic diversity, the data set was filtered in poppr using the Bruvo’s71 distance “farthest neighbour” algorithm, and filtering threshold estimated by the function cutoff_predict (0.02083333). All subsequent analyses were conducted on the filtered dataset.
Measures of genetic diversity
Nei’s72 measure of allelic diversity and number of private alleles were estimated in GenAlEx73, 74. Allelic richness (Ra) and private allelic richness (Pa) were estimated with rarefaction in ADZE v. 1.075. The number of multi-locus genotypes (MLGs), expected number of MLGs after rarefaction (eMLG), and evenness (E5) were estimated in poppr. Simpson’s76 complement index of genotypic diversity (λ; the probability that two randomly selected genotypes are different) was calculated in poppr, and was corrected for sample size by multiplying by N/(N-1). Statistical significance of the differences in genotypic diversities among populations was tested in GenoDive77 using 999 bootstraps. In order to investigate the sufficiency of the microsatellite loci to capture the genotypic diversity of the C. beticola populations, mean genotypic diversity was plotted against the number of loci using MultiLocus v.1.378. Recurrent genotypes (MLGs that occurred more than once) and their frequencies across populations were obtained using poppr. The probability that recurring MLGs could have arisen independently through sexual reproduction was investigated by estimating P sex in GenClone 2.079, and statistical significance was computed by 999 randomisations.
Population differentiation and spatial genetic structure
All analyses of population differentiation were conducted on a priori populations (defined as individuals from the same host/field/state) using both clone-corrected and non-clone-corrected datasets. Pairwise ΦPT 80, an analog of FST assuming a stepwise mutation model, was calculated with 999 randomisations in GenAlEx73, 74. Jost’s81 measure of population differentiation (D) and Hedrick’s82 \({{\rm{G}}}_{{\rm{ST}}}^{\text{'}}\) were estimated using the package mmod 83 in R, with 95% confidence intervals estimated after 1,000 bootstrap simulations. IBD for the five fields within New York was investigated through regression analysis of standardised genetic distance (ΦPT/(1 − ΦPT)) plotted against the logarithm of geographical distance. The Mantel84 test was conducted using ade4 implemented in mmod for both clone-corrected and non-clone-corrected data sets. Discriminant Analysis of Principal Components (DAPC)85 among pre-defined populations from fields were conducted in the R package adegenet v.2.0.186. The optimal number of PCs to retain was estimated through cross-validation using the function xvalDapc. A dendrogram of genetic distance among populations was produced through the unweighted pair group method with arithmetic mean (UPGMA) method in poppr based on Nei’s87 distance. Analysis of molecular variance (AMOVA)88 was conducted using ade4 implemented in poppr (999 permutations), and also by computing a distance matrix (RST) in Arlequin80 (1,000 randomisations). For AMOVA analysis in poppr, the option of filtering with a threshold of 0.05 was used to correct for genotypes that had equivalent distance due to missing data.
Population structure
The existence of an underlying structure without a priori assumption of populations was tested using three methods for both clone-corrected and non-clone-corrected datasets: the model-based Bayesian clustering method implemented in STRUCTURE v.2.3.489, DAPC conducted in the R package adegenet, which does not require assumptions of linkage equilibrium of loci, and a distance-based dendrogram through UPGMA using Bruvo’s71 distance in Poppr. Using STRUCTURE, assignment of MLGs to clusters was inferred for one to 15 clusters. Each model was simulated 10 times with a burn-in period of 100,000 Monte Carlo Markov Chains, and a run length of 1,000,000 iterations. The optimal K was chosen by computing ΔK90 using STRUCTURE HARVESTER v.0.6.9491. The 15 replicated runs for the optimal K were combined and a single graphical output was generated using CLUMPAK92. DAPC analysis was conducted using adegenet, with the optimal number of clusters determined using the function find.clusters. DAPC was then used to assign individuals into clusters, retaining the number of principal components encompassing 89% of the cumulative variance. A distance-based dendrogram using Bruvo’s distance was plotted using poppr with 1,000 bootstrap replicates, and branch support values greater than 50% were retained.
Mating-type determination
For mating-type determination, all isolates were initially screened in a multiplex PCR using CercosporaMat1F/R26 and 356/35828 primers. Isolates carrying the MAT1-1 and MAT1-2 alleles were expected to produce amplicons of 805 and 442 bp, respectively. However, in 30% of the isolates, multiple bands were amplified, which confused mating-type determination (data not shown). This could be due to the fact that C. beticola isolates, in addition to a complete MAT1 idiomorph, contain exonic fragments of both MAT1-1 and MAT1-2 alleles in their genome93. Thus, to improve specificity of the assay, new PCR primers were designed to the non-exonic regions of the MAT1 alleles (Fig. 7; Table 5), which were further used to determine the mating-type of the remaining isolates. Two assays were developed: 1) a three-primer assay consisting of one primer common to the two mating-type idiomorphs (CbMAT-R2; Fig. 7), and two primers specific to each of the mating-types (CbMAT1-F and CbMAT2-R); and 2) a four-primer assay consisting of two primers specific to MAT1-1-1 (CbMAT1-F and CbMAT1-R) and a primer specific to MAT1-2-1 (CbMAT2-F2), which was paired with a primer developed by Bolton et al.28 to amplify a product specific to MAT1-2 isolates.
The PCR mix for both the three-primer and four-primer assays (15 µl total volume) contained 1× PCR buffer (New England Biolabs, Inc., Ipswich, MA) including 1.5 mM MgCl2, 0.1 mM dNTPs (New England Biolabs), 0.1 μM of each primer, 0.8 U Taq polymerase (New England Biolabs), and 10 ng of template DNA. PCR conditions included an initial denaturation for 5 min at 95 °C; followed by 34 cycles of denaturation at 95 °C for 30 s; annealing at 62 °C and 58 °C for the three-primer and four-primer assays, respectively, for 30 s; and extension at 68 °C for 30 s; and a final extension at 68 °C for 5 min.
Tests for random mating
For each field, significant departures of mating-type frequencies from a 1:1 ratio were tested with a chi-square (χ2) goodness-of-fit test, for both clone-corrected and non-clone-corrected datasets. To investigate random association of microsatellite loci, the index of association (I a )94 and standardized index of association (\(\overline{{r}_{d}}\))78, which is independent of the number of loci, were estimated after 1,000 permutations in poppr, for both clone-corrected and non-clone corrected datasets from each field. Since population structure/admixture can also result in detection of linkage disequilibrium even when the population is randomly mating20, tests of random mating were also estimated for the clusters detected by Bayesian analysis, before and after clone-correction. These analyses could not be conducted for the six clusters detected in DAPC analysis due to the small number of the individuals in each cluster (especially after clone correction), which limits the power of LD analysis20, 95.
Detecting recombination
To investigate evidence for recombination in C. beticola populations, the proportion of compatible pairs of loci (PrCP)96 was estimated using MultiLocus. Methods based on the principle of compatibility among sites/loci are among the most powerful to detect recombination97. Two loci are considered compatible (PrCP = 1), if it is possible to account for all the observed genotypes by mutations only; without having to infer homoplasy (reversals, parallelisms, or convergences) or recombination. Under the assumption that recurrent, parallel and reverse mutations are very rare, phylogenetic incompatibility (PrCP < 1) provides evidence of genetic exchange, and may be interpreted as presence of recombination54. PrCP was estimated for C. beticola populations in each field as well as the clusters detected by Bayesian analysis. The statistical significance for the PrCP test was inferred by comparing the number of compatible pairs of loci in the observed data set to those from a randomly recombined data set in MultiLocus. The null hypothesis of random mating was rejected if more compatible loci than expected in a randomised population were observed (P < 0.05)98, 99.
References
McDonald, B. A. How can research on pathogen population biology suggest disease management strategies? The example of barley scald (Rhynchosporium commune). Plant Pathol. 64, 1005–1013 (2015).
McDonald, B. A. & Mundt, C. C. How knowledge of pathogen population biology informs management of Septoria tritici blotch. Phytopathology 106, 948–955 (2016).
Milgroom, M. G. & Peever, T. L. Population biology of plant pathogens: the synthesis of plant disease epidemiology and population genetics. Plant Dis. 87, 608–617 (2003).
Linde, C. Population genetic analyses of plant pathogens: new challenges and opportunities. Australas. Plant Pathol. 39, 23–28 (2010).
Grünwald, N. J. & Goss, E. M. Evolution and population genetics of exotic and re-emerging pathogens: novel tools and approaches. Annu. Rev. Phytopathol. 49, 249–267 (2011).
Goss, E. M. et al. The Irish potato famine pathogen Phytophthora infestans originated in central Mexico rather than the Andes. Proc. Natl. Acad. Sci. 111, 8791–8796 (2014).
Milgroom, M. G., Jiménez-Gasco, M., Olivares-García, C. & Jiménez-Díaz, R. M. Clonal expansion and migration of a highly virulent, defoliating lineage of Verticillium dahliae. Phytopathology 106, 1038–1046 (2016).
Prospero, S., Lutz, A., Tavadze, B., Supatashvili, A. & Rigling, D. Discovery of a new gene pool and a high genetic diversity of the chestnut blight fungus Cryphonectria parasitica in Caucasian Georgia. Infect. Genet. Evol. 20, 131–139 (2013).
Zaffarano, P. L., McDonald, B. A., Zala, M. & Linde, C. Global hierarchical gene diversity analysis suggests the Fertile Crescent is not the center of origin of the barley scald pathogen Rhynchosporium secalis. Phytopathology 96, 941–950 (2006).
Bennett, R. S., Milgroom, M. G., Sainudiin, R., Cunfer, B. M. & Bergstrom, G. C. Relative contribution of seed-transmitted inoculum to foliar populations of Phaeosphaeria nodorum. Phytopathology 97, 584–591 (2007).
Sommerhalder, R. J., McDonald, B. A., Mascher, F. & Zhan, J. Sexual recombinants make a significant contribution to epidemics caused by the wheat pathogen Phaeosphaeria nodorum. Phytopathology 100, 855–862 (2010).
Estoup, A. & Guillemaud, T. Reconstructing routes of invasion using genetic data: why, how and so what? Mol. Ecol. 19, 4113–4130 (2010).
Grünwald, N. J., Garbelotto, M., Goss, E. M., Heungens, K. & Prospero, S. Emergence of the sudden oak death pathogen Phytophthora ramorum. Trends Microbiol. 20, 131–138 (2012).
Schoebel, C. N., Stewart, J., Grunwald, N. J., Rigling, D. & Prospero, S. Population history and pathways of spread of the plant pathogen Phytophthora plurivora. PloS One 9, e85368 (2014).
Zaffarano, P. L., McDonald, B. A. & Linde, C. Phylogeographical analyses reveal global migration patterns of the barley scald pathogen Rhynchosporium secalis. Mol. Ecol. 18, 279–293 (2009).
Banke, S. & McDonald, B. A. Migration patterns among global populations of the pathogenic fungus Mycosphaerella graminicola. Mol. Ecol. 14, 1881–1896 (2005).
Goodwin, S. B., Cohen, B. A., Deahl, K. L. & Fry, W. E. Migration from northern Mexico as the probable cause of recent genetic changes in populations of Phytophthora infestans in the United States and Canada. Phytopathology 84, 553–558 (1994).
Milgroom, M. G., del M Jiménez-Gasco, M., García, C. O., Drott, M. T. & Jiménez-Díaz, R. M. Recombination between clonal lineages of the asexual fungus Verticillium dahliae detected by genotyping by sequencing. PloS One 9, e106740 (2014).
Meng, J. et al. Population genetic analysis reveals cryptic sex in the phytopathogenic fungus Alternaria alternata. Scientific Reports 5, 18250 (2015).
Milgroom, M. G. Recombination and the multilocus structure of fungal populations. Annu. Rev. Phytopathol. 34, 457–477 (1996).
Leo, A. E., Ford, R. & Linde, C. C. Genetic homogeneity of a recently introduced pathogen of chickpea, Ascochyta rabiei, to Australia. Biol. Invasions 17, 609–623 (2015).
Pethybridge, S. J., Scott, J. B. & Hay, F. S. Lack of evidence for recombination or spatial structure in Phoma ligulicola var. inoxydabilis populations from Australian pyrethrum fields. Plant Dis. 96, 746–751 (2012).
Vaghefi, N. et al. Rapid changes in the genetic composition of Stagonosporopsis tanaceti population in Australian pyrethrum fields. Phytopathology 105, 358–369 (2015).
Burt, A., Carter, D. A., Koenig, G. L., White, T. J. & Taylor, J. W. Molecular markers reveal cryptic sex in the human pathogen Coccidioides immitis. Proc. Natl. Acad. Sci. USA 93, 770–773 (1996).
Franc, G. D. Ecology and epidemiology of Cercospora beticola In Cercospora Leaf Spot of Sugar Beet and Related Species (eds. Lartey, R. T., Weiland, J. J., Panella, L., Crous, P. W. & Windels, C. E.) 7–19 (American Phytopathological Society, 2010).
Groenewald, M., Groenewald, J. Z., Harrington, T. C., Abeln, E. C. & Crous, P. W. Mating type gene analysis in apparently asexual Cercospora species is suggestive of cryptic sex. Fung. Genet. Biol. 43, 813–825 (2006).
Groenewald, M., Linde, C. C., Groenewald, J. Z. & Crous, P. W. Indirect evidence for sexual reproduction in Cercospora beticola populations from sugar beet. Plant Pathol. 57, 25–32 (2008).
Bolton, M. D. et al. Evaluation of the potential for sexual reproduction in field populations of Cercospora beticola from USA. Fungal Biol. 116, 511–521 (2012).
Obuya, J. O., Hanson, L. E. & Franc, G. D. Mating type idiomorphs distribution and their correlation to benzimidazole-resistance in Cercospora beticola from the Central High Plains region, USA. American Society of Sugar Beet Technologists, Proceedings from the 36th Biennial Meeting, doi:pp.10.5274.ASSBT.2011.114 (2011).
Bakhshi, M., Arzanlou, M. & Babai-Ahari, A. Uneven distribution of mating type alleles in Iranian populations of Cercospora beticola, the causal agent of Cercospora leaf spot disease of sugar beet. Phytopathol. Mediterr. 50, 101–109 (2011).
Koike, S. T., du Toit, L. J. & Abawi, G. S. Cercospora leaf spot of vegetable crops in the Chenopodiaceae. In: Cercospora Leaf Spot of Sugar Beet and Related Species (eds. Lartey, R. T., Weiland, J. J., Panella, L., Crous, P. W. & Windels, C. E.) 91–96 (American Phytopathological Society, 2010).
McKay, M. B. & Pool, V. W. Field studies of Cercospora beticola. Phytopathology 8, 119–136 (1918).
Nagel, C. M. The longevity of Cercospora beticola in soil. Phytopathology 28, 342–350 (1938).
Jones, R. K. & Windels, C. E. A management model for Cercospora leaf spot of sugar beets. University of Minnesota Extension Service AG-FO-5643-E (1991).
Khan, J. et al. Survival, dispersal, and primary infection site for Cercospora beticola in sugar beet. Plant Dis. 92, 741–745 (2008).
Solel, Z. Survival of Cercospora beticola, the causal agent of sugar beet leaf spot, in Israel. Trans. Br. Mycol. Soc. 54, 504–506 (1970).
Vaghefi, N., Hay, F. S., Kikkert, J. R. & Pethybridge, S. J. Genotypic diversity and resistance to azoxystrobin of Cercospora beticola on processing table beet in New York. Plant Dis. 100, 1466–1473 (2016).
Vestal, E. F. Pathogenicity, host response and control of Cercospora leaf-spot of sugar beets. Research Bulletin No 168 (Agricultural Experiment Station, Iowa State College of Agriculture and Mechanic Arts, Ames, 1933).
Meredith, D. S. Conidium release and dispersal in Cercospora beticola. Phytopathology 57, 889–893 (1967).
Turgay, E. B., Bakir, M., Ozeren, P., Katircioglu, Y. Z. & Maden, S. Detection of pathotypes and genetic diversity of Cercospora beticola. Plant Pathol. J. 26, 306–312 (2010).
Moretti, M., Karaoglanidis, G., Saracchi, M., Fontana, A. & Farina, G. Analysis of genotypic diversity in Cercospora beticola Sacc. field isolates. Ann. Microbiol. 56, 215–221 (2006).
Barrès, B. et al. Genetic structure of the poplar rust fungus Melampsora larici-populina: evidence for isolation by distance in Europe and recent founder effects overseas. Infect. Genet. Evol. 8, 577–587 (2008).
Rousset, F. Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance. Genetics 145, 1219–1228 (1997).
Bobev, S. G., Groenewald, J. Z. & Crous, P. W. First report of Cercospora beticola as a pathogen of German statice (Goniolimon tataricum) in Bulgaria. Plant Dis. 93, 553–553 (2009).
Groenewald, J. Z., Groenewald, M., Braun, U. & Crous, P. W. Cercospora speciation and host range In Cercospora leaf spot of sugar beet and related species (eds. Lartey, R. T., Weiland, J. J., Panella, L., Crous, P. W. & Windels, C. E.) 21–37 (American Phytopathological Society, 2010).
Houessou, J. H., Beed, F., Sikirou, R. & Ezin, V. First report of Cercospora beticola on lettuce (Lactuca sativa) in Benin. New Dis. Rep. 23, 16 (2011).
Lartey, R. T. et al. Safflower: A new host of Cercospora beticola. Plant Dis. 89, 797–801 (2005).
Rooney-Latham, S., Scheck, H. J. & Walber, T. M. First report of Cercospora beticola causing a leaf spot disease on Acanthus mollis in California. Plant Dis. 95, 224–224 (2011).
Vereijssen, J., Schneider, J. H. & Termorshuizen, A. J. Root infection of sugar beet by Cercospora beticola in a climate chamber and in the field. Eur. J. Plant Pathol. 112, 201–210 (2005).
Linde, C. C., Zhan, J. & McDonald, B. A. Population structure of Mycosphaerella graminicola: from lesions to continents. Phytopathology 92, 946–955 (2002).
McDonald, B. A., Zhan, J. & Burdon, J. J. Genetic structure of Rhynchosporium secalis in Australia. Phytopathology 89, 639–645 (1999).
Burdon, J. J., Marshall, D. R., Luig, N. H. & Gow, D. J. S. Isozyme studies on the origin and evolution of Puccinia graminis f. sp. tritici in Australia. Aus. J. Biol. Sci. 35, 231–238 (1982).
Goodwin, S. B., Cohen, B. A. & Fry, W. E. Panglobal distribution of a single clonal lineage of the Irish potato famine fungus. Proc. Natl. Acad. Sci. USA 91, 11591–11595 (1994).
Milgroom, M. G. Population Biology of Plant Pathogens: Genetics, Ecology, and Evolution. (APS Press, 2015).
Drenth, A., Janssen, E. M. & Govers, F. Formation and survival of oospores of Phytophthora infestans under natural conditions. Plant Pathol. 44, 86–94 (1995).
Milgroom, M. G. et al. Clonal population structure of the chestnut blight fungus in expanding ranges in southeastern Europe. Mol. Ecol. 17, 4446–4458 (2008).
Putman, A. I. & Carbone, I. Challenges in analysis and interpretation of microsatellite data for population genetic studies. Ecol. Evol. 4, 4399–4428 (2014).
Carlson, L. W. Relation of weather factors to dispersal of conidia of Cercospora beticola (Sacc). J. Am. Soc. Sugar Beet Tech. 14, 319 (1967).
Vereijssen, J., Schneider, H. J. H. M. & Termorshuizen, A. A. J. Possible root infection of Cercospora beticola in sugar beet. Eur. J. Plant Pathol. 110, 103–106 (2004).
Rivas, G. G., Zapater, M. M., Abadie, C. & Carlier, J. Founder effects and stochastic dispersal at the continental scale of the fungal pathogen of bananas. Mycosphaerella fijiensis Mol. Ecol. 13, 471–482 (2004).
Vereijssen, J., Schneider, J. H., Stein, A. & Jeger, M. J. Spatial pattern of Cercospora leaf spot of sugar beet in fields in long-and recently-established areas. Eur. J. Plant Pathol. 116, 187–198 (2006).
Lawrence, J. S. & Meredith, D. S. Wind dispersal of conidia of Cercospora beticola. Phytopathology 60, 1076–1078 (1970).
Bolton, M. D., Rivera, V. & Secor, G. Identification of the G143A mutation associated with QoI resistance in Cercospora beticola field isolates from Michigan, United States. Pest Manag. Sci. 69, 35–39 (2013).
Groenewald, M., Groenewald, J. Z. & Crous, P. W. Distinct species exist within the Cercospora apii morphotype. Phytopathology 95, 951–959 (2005).
Groenewald, M., Groenewald, J. Z., Linde, C. C. & Crous, P. W. Development of polymorphic microsatellite and single nucleotide polymorphism markers for Cercospora beticola (Mycosphaerellaceae). Mol. Ecol. Notes 7, 890–892 (2007).
Vaghefi, N. et al. De novo genome assembly of Cercospora beticola for microsatellite marker development and validation. Fungal Ecol. 26, 125–134 (2017).
Kearse, M. et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649 (2012).
Pompanon, F., Bonin, A., Bellemain, E. & Taberlet, P. Genotyping errors: causes, consequences and solutions. Nat. Rev. Genet. 6, 847–846 (2005).
Kamvar, Z. N., Tabima, J. F. & Grünwald, N. J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2, 281 (2014).
Kamvar, Z. N., Brooks, J. C. & Grünwald, N. J. Novel R tools for analysis of genome-wide population genetic data with emphasis on clonality. Front. Genet. 6, 208 (2015).
Bruvo, R., Michiels, N. K., D’Souza, T. G. & Schulenburg, H. A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 13, 2101–2106 (2004).
Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89, 583–590 (1978).
Peakall, R. O. D. & Smouse, P. E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 6, 288–295 (2006).
Peakall, R. O. D. & Smouse, P. E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28, 2537–2539 (2012).
Szpiech, Z. A., Jakobsson, M. & Rosenberg, N. A. ADZE: a rarefaction approach for counting alleles private to combinations of populations. Bioinformatics 24, 2498–2504 (2008).
Simpson, E. H. Measurement of diversity. Nature 163, 688 (1949).
Meirmans, P. G. & Van Tienderen, P. H. GENOTYPE and GENODIVE: two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 4, 792–794 (2004).
Agapow, P. & Burt, A. Indices of multilocus linkage disequilibrium. Mol. Ecol. Notes 1, 101–102 (2001).
Arnaud-Haond, S. & Belkhir, K. GENCLONE: a computer program to analyse genotypic data, test for clonality and describe spatial clonal organization. Mol. Ecol. Notes 7, 15–17 (2007).
Excoffier, L., Laval, G. & Schneider, S. Arlequin ver. 3.0: an integrated software package for population genetics data analysis. Evol. Bioinf. 1, 47–50 (2005).
Jost, L. GST and its relatives do not measure differentiation. Mol. Ecol. 17, 4015–4026 (2008).
Hedrick, P. W. A standardized genetic differentiation measure. Evolution 59, 1633–1638 (2005).
Winter, D. J. MMOD: an R library for the calculation of population differentiation statistics. Mol. Ecol. Res. 12, 1158–1160 (2012).
Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 27, 209–220 (1967).
Jombart, T., Devillard, S. & Balloux, F. Discriminant analysis of principal components: a new method for the analysis of genetically structured populations. BMC Genetics 11, 94 (2010).
Jombart, T. Adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 24, 1403–1405 (2008).
Nei, M. Genetic distance between populations. Am. Nat., 283–292 (1972).
Excoffier, L., Smouse, P. E. & Quattro, J. M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131, 479–491 (1992).
Pritchard, J. K., Stephens, M. & Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 155, 945–959 (2000).
Evanno, G., Regnaut, S. & Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611–2620 (2005).
Earl, D. A. & vonHoldt, B. M. “STRUCTURE HARVESTER”: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Res. 4, 359–361 (2012).
Kopelman, N. M., Mayzel, J., Jakobsson, M., Rosenberg, N. A. & Mayrose, I. Clumpak: a program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Res. 15, 1179–1191 (2015).
Bolton, M. D. et al. The heterothallic sugar beet pathogen Cercospora beticola contains exon fragments of both MAT genes that are homogenized by concerted evolution. Fung. Genet. Biol. 62, 43–54 (2014).
Brown, A. H. D., Feldman, M. W. & Nevo, E. Multilocus structure of natural populations of Hordeum spontaneum. Genetics 96, 523–536 (1980).
Brown, A. H. D. Sample sizes required to detect linkage disequilibrium between two or three loci. Theor. Popul. Biol. 8, 184–201 (1975).
Estabrook, G. F. & Landrum, L. A simple test for the possible simultaneous evolutionary divergence of two amino acid positions. Taxon 24, 609–613 (1975).
Posada, D. & Crandall, K. A. Evaluation of methods for detecting recombination from DNA sequences: computer simulations. Proc. Natl. Acad. Sci. 98, 13757–13762 (2001).
Bennett, R. S., Milgroom, M. G. & Bergstrom, G. C. Population structure of seedborne Phaeosphaeria nodorum on New York wheat. Phytopathology 95, 300–305 (2005).
Xu, J., Yan, Z. & Guo, H. Divergence, hybridization, and recombination in the mitochondrial genome of the human pathogenic yeast. Cryptococcus gattii. Mol. Ecol. 18, 2628–2642 (2009).
Acknowledgements
This research was supported by the United States Department of Agriculture, National Institute of Food and Agriculture Hatch project NYG-625424, and the Federal Capacity Funds Initiative (2015-16-118) managed by the New York State Agricultural Experiment Station (NYSAES), Cornell University, Geneva, New York, USA, and the NYSAES Director’s Controlled Endowment Fund. Thanks to Ms. Carol Bowden, Ms. Elizabeth Maloney and Mr. David Strickland for help with sample collection and excellent technical support. Thanks to growers and gardeners in New York and Hawaii for allowing access to their fields and plots for sample collection. We would like to thank the anonymous reviewer for constructive suggestions on an earlier version of this manuscript.
Author information
Authors and Affiliations
Contributions
N.V., J.R.K. and S.J.P. contributed to experimental design. N.V., J.R.K., S.C.N., and S.J.P. contributed to sample collection; N.V. performed the experiments, generated and analysed the data; N.V. and S.J.P. wrote the manuscript; J.R.K. and S.C.N. reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vaghefi, N., Nelson, S.C., Kikkert, J.R. et al. Genetic structure of Cercospora beticola populations on Beta vulgaris in New York and Hawaii. Sci Rep 7, 1726 (2017). https://doi.org/10.1038/s41598-017-01929-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-01929-4
This article is cited by
-
Determination of mating types of Fulvia fulva isolates from Turkey
Tropical Plant Pathology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
