
Abstract
Pyrogenic carbon (PyC) and n-alkane data from sediments in the northern South China Sea reveal variations in material from C4 plants in East Asia over the last ~19 Ma. These data indicate the likely presence of C4 taxa during the earliest part of the record analysed, with C4 species also prominent during the mid and late Miocene and especially the mid Quaternary. Notably the two records diverge after the mid Quaternary, when PyC data indicate a reduced contribution of C4 taxa to biomass burning, whereas plant-derived n-alkanes indicate a greater abundance of C4 plants. This divergence likely reflects differences in the predominant source areas of organic materials accumulating at the coring site, with PyC representing a larger source area that includes material transported in the atmosphere from more temperate (relatively cooler and drier) parts of East Asia. Variations in the relative abundances of C3 and C4 taxa appear to be linked to a combination of environmental factors that have varied temporally and geographically and that are unique to East Asia. A major expansion of C4 biomass in warmer subtropical parts of eastern Asia from ~1 Ma and particularly from ~0.4 Ma is later than other parts of the world.
Similar content being viewed by others

Introduction
The Calvin-Benson cycle, the process through which plants convert inorganic carbon (C) and water to three C (C3) sugar molecules, originated when atmospheric composition was very different from present1, 2. One modification to the cycle, leading to reduced photorespiration effects and improved photosynthetic efficiency under certain conditions (e.g. moisture stress and relatively low pCO2), involves production of four C (C4) oxaloacetate as the first-formed product of photosynthesis. Uncertainty surrounds the exact date of origin of this modification. That said, there is no widely accepted evidence of C4 taxa pre-dating the Oligocene3, with molecular studies indicating that the C4 photosynthetic pathway first appeared between 35 and 30 Ma4, a period that includes a major decline in pCO2 5. Only ~3% of angiosperm species currently support the C4 pathway. However, despite being utilized by a relatively small proportion of the global flora, C4 taxa – predominantly in the form of grasses – account for a substantial proportion of vegetation cover of Earth6, and include important food staples.
An expansion of plants utilising the C4 photosynthetic pathway (e.g., tropical grasses) relative to those using the C3 pathway (e.g., trees, shrubs and temperate grasses) in the late Miocene constitutes one of the most important biogeographical transformations of the Cenozoic7,8,9,10. The timing of this expansion appears to have varied geographically, occurring later in East Asia than other parts of the world11,12,13,14,15. As evidence has accumulated, however, the expansion of C4 plants in East Asia appears to have been phased, rather than a single event, and may have occurred much earlier in some parts of the region16, 17, than in others18. Determining when and where the C4 photosynthetic pathway first appeared on the Asian continent is difficult because direct evidence, in the form of plant remains such as pollen and phytoliths, is rare. Furthermore, when records are available, they are often incomplete; C4 taxa often occupy environments that are not conducive to the preservation of organic material in situ, and achieving a high level of certainty in distinguishing between many C4 and C3 taxa in pollen and phytolith records can be problematic.
Stable C isotope (δ13C) values for C3 plants globally range between −37‰ and −20‰ (average −28.5‰)19, while the δ13C values for C4 plants range from −15‰ to −9‰ (average −13‰)20,21,22. Pyrogenic carbon (PyC), also known as elemental carbon (EC) or black carbon (BC), is produced through the incomplete combustion of biomass and can preserve the original C isotope value of the parent plants23. PyC is generally resistant to decomposition, and upon extraction from sediments can be used not only to document the frequency and intensity of past fire activity, but also changes in the relative contributions of C4 and C3 plants in combusted biomass15, 16, 23,24,25. Similarly, long chain n-alkanes (≥n-C27), which are important building blocks of lipids forming the protective waxy cuticle covering leaf and stem tissue, are resistant to diagenetic alteration26. Where herbaceous vegetation predominates, n-alkanes of C31 and C33 are abundant components of sediments, while larger proportions of n-alkanes of C27 and C29 are generally indicative of wooded and forested landscapes (i.e. C3 taxa)27,28,29. Thus ratios of measured levels of n-alkanes, such as C31/C27, can be used to reconstruct the relative contributions of herbaceous and woody plants to organic C preserved in sediments27. Furthermore, although long chain n-alkanes typically have lower δ13C values than bulk tissue30, fractionation effects can be removed, so that the PyC and long chain n-alkanes composition of organic material in sediments can be used as a basis for inferring past variations in the relative abundances of C3 and C4 plants in, respectively, combusted biomass and vegetation23, 30, 31.
The South China Sea (SCS), a marginal sea between Asia and the Pacific Ocean, is dominated climatically by the East Asian monsoon16, 32, 33. Sediments accumulating on the bed of the northern SCS, largely supplied by major rivers but also including aeolian material transported by the East Asian Winter Monsoon (EAWM)34, provide a potential source of information on environmental variations affecting continental East Asia during the late Cenozoic. To date, there is only one sediment-based record of variations in PyC from the northern SCS16. A second core, collected from a deep water site in the southern part of the SCS, has yielded a record of variations in n-alkane content of sediment, presumably largely of low latitude origin, for the last 5 Ma29. Here we present new data in the form of variations in PyC and n-alkanes from the same sediment core from the northern SCS relating to the last ~19 Ma. These new data are compared with published information in order to describe and explain the unique history of C3 and C4 taxa in East Asia since the early Miocene.
Study area, materials and methods
The Ocean Drilling Program (ODP) Leg 184, Site 1146 (19°27.4′N, 116°16.4′E, 2092 m water depth) retrieved sediment from a small rift basin on the mid-continental slope of the northern SCS, located less than 50 km to the northeast of ODP Site 1147/1148 (Fig. 1). The sediment sequence obtained at ODP Site 1146 extended to a sub-seafloor depth of 643 m composite depth (mcd). For this study, a total of 149 2-cm-thick sample slices were collected at 4 m intervals between 641 mcd and the surface. Chronological control was established on the basis of palaeomagnetism and biostratigraphy35 (S1, Supplementary material), with the ages of sample depths between control points established through linear interpolation (Fig. S1). According to the age-depth model for ODP Site 114635, the sequence of sediments between 641 mcd and the surface covers the last ~19 Ma, with the sampling interval of 4 m adopted in the current research equating to a temporal resolution of ~100 ka.

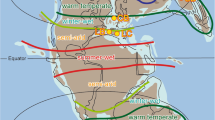
Map showing location of ODP Site 1146 and other important sites mentioned in the paper. The upper left panel contains the legend for the map. Ocean core locations ODP Site 1081 (South Atlantic, 19°37.2′S, 11°19.2′E, 793.8 m water depth), ODP site 717 (Bengal Fan, 0°56′S, 81°23′E, 4734.7 m water depth) and DSDP Leg 10 Site 94 (Gulf of Mexico, 24°20′N, 88°28.2′W, 1793 m water depth) mentioned in the text of the paper are not shown. The map was generated specifically for this paper using the ETOPO5 gridded elevation dataset (Version ETOPO5, http://www.ngdc.noaa.gov/mgg/global/etopo5.HTML) and Coreldraw (Version Coreldraw X7, http://www.corel.com/cn); no copyright problems are envisaged.
PyC was extracted using the method of Gustafsson et al.36. Sediment samples were pretreated with 1 M NaOH, HCl and HNO3 to remove inorganic C and some of the organic forms that might lead to BC formation during the pre-combustion step. PyC was then measured following combustion at 375 °C in air for 24 h, with the δ13C values for PyC (δ13CPyC) values determined using a MAT-253 mass spectrometer. Isotopic compositions are expressed as deviations relative to the V-PDB standard with a precision of ±0.2‰ or better. This procedure differs from the oxidation method used to measure PyC (therein termed BC) in sediments from ODP Sites 1147/1148 and the Lingtai Section of the Chinese Loess Plateau (CLP)16, 37.
For analysis of high molecular weight n-alkanes, samples were freeze-dried before being ground manually. After adding an internal standard (nC24D50), a ~3 g sample was ultrasonically extracted in dichloromethane–methanol (3:1, v/v). Extracts were dried and saponified with 6% KOH–methanol. Hexane was then used to extract hydrocarbons, which were separated by column chromatography into the polar (i.e. alkanol) and non-polar (i.e. alkane) fractions. The δ13C values for individual alkanes (δ13CAlk) were determined using a GV Isochrom II system interfaced to a Hewlett-Packard 5890 gas chromatograph. The gas chromatograph was fitted with a fused Si column HP-5 MS (30 m-0.32 mm-0.25 mm) connected to the combustion interface. Helium was used as the carrier gas with a flow rate of 1.2 mL min−1. The temperature was programmed to remain at 80 °C for 2 min, before rising to 220 °C at a rate of 10 °C min−1, and then to 290 °C at a rate of 3 °C min−1. The temperature was then maintained at 290 °C for 15 min. CO2 was used as a reference gas, which was automatically introduced into the isotope ratio mass spectrometer before and after each analysis. The δ13C values were calibrated against a standard mixture of n-alkanes (nC12–nC32) of known isotopic composition (Indiana University, USA) and reported as ‰ relative to Vienna Peedee Belemnite (V-PDB). Replicate analyses showed that the standard deviation for each compound was less than 0.3‰. In order to maintain the reproducibility and accuracy of results, standards were run between samples, and each sediment sample was analyzed at least twice.
The δ13C value of atmospheric CO2 has not remained constant over time9, 23, 38, 39. Moreover, fractionation effects can occur during the production, transport and sedimentation of organic matter23, 40, 41, and between atmospheric CO2 and plant organic C, as well as varying between different taxa42, 43. These sources of variation introduce additional uncertainty into calculations of the relative contribution to combusted biomass and relative abundance in vegetation of C4 taxa based on, respectively, δ13CPyC and δ13CAlk, and hence need to be accounted for when interpreting records of ancient δ13C. Carbon isotopic enrichment factors for PyC and n-alkanes relative to atmospheric CO2 (εPyC-CO2 and εAlk-CO2, respectively) were thus applied to calculate C4 abundance in the current study (S2 and Fig. S2, Supplementary material).
Results
The PyC content of sediments from ODP Site 1146 (Fig. 2a, Table S1) shows an overall slight increase from the early Miocene through to the Quaternary, ranging from 0.014% to 0.16% (average 0.06%). This trend is superimposed upon strong variability in the data, with the amplitude of variation particularly large from the mid Quaternary. The latter period includes two intervals of high PyC content, centred upon ~1 Ma and 0.2 Ma. δ13CPyC values range from −25‰ to −17.2% (average −21.2‰), and also show an overall increase since the early Miocene, before declining after ~2 Ma (Fig. 2b, Table S1). Relatively high (less negative) δ13CPyC values date to the early Miocene (~18–19 Ma), mid Miocene (~14 Ma), late Miocene/early Pliocene (~7–4.5 Ma), and to the early to mid Quaternary (~2–1 Ma).

Abundance and carbon isotope composition data from n-alkanes and pyrogenic C (PyC) from ODP core 1146: (a) PyC content; (b) δ13C values of PyC (δ13CPyC); (c) C31/C27 n-alkane ratio; (d) Average δ13C values of long-chain n-alkanes (δ13CAlk); (e) Record of δ13C of atmospheric CO2 39; (f) The contribution of C4 taxa to combusted biomass based on carbon isotopic enrichment factor for PyC (εPyC-CO2); (g) C4 abundance based on carbon isotopic enrichment factor for n-alkanes (εAlk-CO2). Arrows indicate the overall trend in proxies. Two yellow bars show the area of uncertainty in reconstructing the relative prominence of C3 and C4 taxa based on εPyC-CO2 and εAlk-CO2, and thus the basis for the very conservative estimates of prominence of C4 taxa mentioned in the text of this paper.
Trends in C31/C27 ratios (Fig. 2c, Table S1) are broadly similar to those exhibited by PyC; C31/C27 ratios vary over the last ~19 Ma, showing an overall slight increase to ~1 Ma, with an increased frequency of above average values after ~7.5 Ma, exhibiting greatest variability and highest values from ~1 Ma. In addition, C31/C27 ratios fluctuate through the early and mid Miocene, with four peaks around ~18 Ma, ~15.5 Ma, ~14 Ma and ~11 Ma.
The weighted mean δ13C values of long chain alkanes, determined from (nC27 * δ13CnC27 + nC29 * δ13CnC29 + nC31*δ13CnC31)/(nC27 + nC29 + nC31), is abbreviated to δ13CAlk. Values of δ13CAlk varied from −33.2‰ to −25.6‰ (average −29.9‰) (Fig. 2d, Table S1). Although fluctuating, an overall trend of declining (increasingly negative) values is evident, from the earliest parts of the record (~19 Ma) to the mid Quaternary, with three zones of relatively high (less negative) δ13CAlk values dating to around the early Miocene (18–19 Ma), late mid Miocene (11.5–12.5 Ma), and middle and later part of the late Miocene (~7.5–5.2 Ma). δ13CAlk values peak (least negative) after ~1 Ma, especially from 0.4 Ma. Over the entire sequence of sediments described here, trends in δ13CAlk values are generally opposite to those of δ13CPyC, particularly from the early Quaternary and especially from ~1 Ma.
Variations in δ13C value of atmospheric CO2 39 (Fig. 2e), interpolated to correspond directly with dated δ13CPyC and δ13CAlk records, were used to derive estimates of εPyC-CO2 and εAlk-CO2, respectively. Conservative estimates of the relative contribution of C4 taxa to combusted biomass and vegetation ranged from, respectively, 0–34.7% (average 12.9%) and 0–41.6% (average 9.9%) (Fig. 2f and g). Values of both εPyC-CO2 and εAlk-CO2 indicate the likely presence of C4 taxa in East Asia during the early Miocene, later part of mid Miocene and the late Miocene through to the Late Quaternary. Some differences are evident in the two sources of information, however. The εPyC-CO2 data tend to indicate a relatively significant and continuous, though strongly varying, C4 contribution to combusted biomass, especially when compared with the corresponding estimated abundance of C4 taxa in terrestrial vegetation based on εAlk-CO2 data. The largest differences between the two datasets date to the Pliocene and Quaternary; εPyC-CO2 data indicate an overall substantial contribution of C4 grasses to combusted biomass from the late Miocene through to the Quaternary, though with an overall decline evident after ~2 Ma. Lower εAlk-CO2 values indicate a predominance of C3 grasses from around the beginning of the Pliocene through to ~1 Ma. This is supported to an extent by the enriched C31/C27 ratios, which suggest a greater herbaceous contribution to organic C from the late Miocene. An expansion of herbaceous vegetation generally (including a greater proportion of C4 grasses at times) from ~1 Ma, particularly from ~0.4 Ma, is suggested by enriched high δ13CAlk data values and generally higher average C31/C27 ratios.
Discussion
Deviation of PyC and n-alkane records in ODP 1146 from northern SCS
Differences in n-alkane and PyC data from the same core likely reflect differences in both provenance of the C on which the measurements are based, and in the mode of transport of the C from the terrestrial source to the site of deposition. PyC is formed at high temperatures during the combustion of biomass and emitted to the atmosphere before a proportion is deposited in sedimentary environments44, with the amount deposited dependent on atmospheric transport (influenced by wind velocity and direction and topography), precipitation and depositional processes. Several studies have indicated that PyC preserved in deep-sea sediments in the SCS originates via long-range transport from burning in central and southeastern Asia45,46,47,48,49,50,51 (S3, Supplementary material). Atmospheric transport from these sources is particularly active during EAWM, when dry continental conditions can also enhance the likelihood of vegetation fires52, 53. In general, highly seasonal and semi-arid climate conditions will result in biomass that is prone to burning, at least towards the end of the dry season, and, on occasion, in intense fires24. Thus levels of PyC accumulating in sediments in the SCS are affected not only by the extent and composition of vegetation available for combustion, but also by burning regime and by variations in the EAWM in particular24. Variations in PyC therefore likely represent an integration of conditions over a broad geographic area, with the source area of PyC accumulating in the northern SCS potentially including cold-adapted vegetation in more temperate, higher altitude and latitude parts of continental East Asia16. This is evident in similarities between variations in δ13CPyC measured at ODP Site 1146 discussed here and δ13CBC from Site 114816, and from the CLP over the last ~7 Ma15 (Fig. 3).

Summary of data for ODP core 1146 compared with similar data from sites proximate to, and on, the continent of Asia. Charts in the left panel summarise data north to south across East Asia. (a) and (b) δ13CTOC from transect sections on Chinese Loess Plateau (from northwest to southeast: Lantian, Yanyu)12, 18; (c) and (d) δ13Calk values from core 1146 since the last 3 Ma and since about 19 Ma (this study). The right panel summarises data from a far larger area. (e) δ13C values of PyC from core 1146 in this study; (f) and (g) δ13C values of black carbon from 1147/8 and δ13C values of PyC from Lingtai loess section16, 37; (h) and (i) marine sediments in Indian Ocean (ODP 717)62 and Bengal Fan (ODP 722)63; (j) Published pCO2 records since the mid-Miocene based on a compilation of CO2 curves (green line)59; (k) Simplified composite δ18O record of deep-sea benthic foraminifera as a proxy for global temperature variations65. Dashed line shows in enlarged form the record for δ13Calk in core 1146 since the last ~3 Ma. Solid line highlights the divergence of δ13C values occurring since the mid Quaternary. Arrows show direction of overall change in the data.
By comparison, n-alkanes are mainly derived from plant leaves30. Surface sediment n-C29 concentrations are in good agreement with data on pollen from approximately the same suite of modern sediments in the northern SCS54, 55. The main delivery mechanism for n-alkanes is likely to be dominated by fluvial inputs and then marine currents via the Bashi and Taiwan straits55. Some n-alkanes may also have been deposited from the atmosphere, along with aeolian dust and aerosols associated with biomass burning56. However, the fact that levels of n-alkanes and PyC measured in the same sediment samples are not well-correlated suggests the two proxies have different provenances (Fig. S3). Possibly therefore, n-alkanes accumulating in the past in sediments in the northern SCS originated from relatively well-vegetated parts of subtropical East Asia, rather than as a result of long distance aeolian input from more temperate land to the north, where the vegetation cover is comparatively sparse24. Catchments draining into the northern SCS are likely to be the dominant means by which organic material containing plant lipids is transported mainly from sub-tropical latitudes55, with variations in δ13CAlk therefore affected by a different set of drivers to PyC abundance, including changes in monsoonal precipitation and fluvial discharge, catchment conditions and the extent and proximity of land relative to ocean57.
Variations in C4 contribution to vegetation and combusted biomass in East Asia recorded by PyC and n-alkane data
Variations in the prominence of C4 taxa estimated from εPyC-CO2 (relative contribution to combusted biomass) and εAlk-CO2 (relative abundance in vegetation) are generally in phase with the δ13CPyC and δ13CAlk records, respectively. The δ13CPyC values are also in general agreement with variations in δ13CBC from ODP sites 1147/114816 (Fig. 3), notwithstanding the aforementioned systematic offset in values due to differences in the analytical techniques employed23. Variations in δ13CAlk are also within a similar range to that observed in a previously reported record from the northern SCS57 covering a shorter period of time than that discussed here.
The presence of C4 taxa during the early (~18.5 Ma) and mid Miocene (~14 Ma) and from the late Miocene (~7.5 Ma) is supported by existing data9, 15,16,17, 58. Variations in abundances of C4 taxa have been attributed to the changes in pCO2 10, 59 and to evolution of East Asian monsoon12, 16, seasonality18, aridity and/or fires15, 58. Intensification of the East Asian Summer Monsoon (EASM) is thought to date to about 24 Ma, with a peak in intensity during the mid Miocene, a weakening after ~8 Ma and strengthening again during the late Pliocene/early Quaternary29, 30, while strengthening of the EAWM is thought to have occurred 16–14 Ma, ~8 Ma, and ~3 Ma and to have further intensified from the mid Quaternary60. Increasing PyC content indicates rising levels of biomass burning, with periods of relatively high contributions to combusted biomass from C4 taxa evident ~14 Ma, ~7–4.5 Ma and ~2.2–1.5 Ma. Increased contributions of C4 taxa to biomass burning may represent a climatically promoted competitive advantage for C4 photosynthesis, due not only to a strengthened EAWM and greater aridity57, 61, but also to increased seasonality as a result of renewed intensification of the EASM, especially from ~3 Ma. From the mid Quaternary, both above average PyC levels and C31/C27 ratios could be a response to increased aridity, possibly due to a strengthening of the EAWM following further expansion of the northern hemisphere ice sheets12, 29, 32. The contribution of C4 taxa to combusted biomass declines from ~2 Ma, presumably because of a greater presence of C3 plants in the source region for PyC. Evidence from the CLP for increased prominence of C3 taxa during much of the Quaternary may represent a combination of a more intense EAWM and reduced temperatures during the main growing season in more temperate parts of the region, with both factors favouring C3 photosynthesis15, 62.
At lower latitudes, C3 taxa were prominent from the mid Miocene to the Pliocene/Quaternary boundary, under warm and humid climate conditions presumably owing to the influence of the EASM33. Reduced moisture and increased seasonality from the late Pliocene through to ~1 Ma, with the majority of precipitation occurring during the growing season29, could have facilitated sustained increases in the relative abundance of C3 grasses, as evidenced by relatively lower εAlk-CO2 values and higher C31/C27 ratios. As seasonal conditions developed and vegetation canopies became more open, expansion of C4 biomass could have become tightly coupled in a feedback relationship to changes in burning regime and aridity that effectively selected against shrubs and trees (i.e. C3 species) and promoted further expansion of grasslands in which C4 taxa were prominent. An expansion of C4 taxa in subtropical East Asia from ~1 Ma, and especially ~0.4 Ma, may have been triggered by increased seasonality, aridity and changes to the fire regime.
A synthesis of evidence for C4 variations across Asia
δ13C values from fossil enamel and soils from the northwestern CLP (the northeastern margin of the Qinghai–Tibet Plateau) suggest increased abundance of C4 taxa from ~3 Ma, with C4 taxa becoming a significant component of the local vegetation around ~1.0 Ma13. An expansion of C4 biomass ~1.0 Ma and, at lower altitude, ~0.4 Ma, on the southeastern parts of the CLP is evident in δ13C data from Yanyu (620 m amsl) and Lantian (769 m amsl)12, 18 (Fig. 3). The evidence from Lantian and Yanyu is thus consistent with our δ13CAlk data from the northern SCS.
The δ13C values of n-alkanes obtained from palaeosols associated with the Siwalik Group and marine sediments from the Indus (ODP Site 722) and Bengal (ODP Site 717C) fans63, 64, dating to 12–11 Ma, indicate a predominantly C3 flora during the mid Miocene (Fig. 3). A subsequent expansion of C4 taxa during the late Miocene in western and southern Asia has been linked to aridification64, although other factors, such as increased burning15, 25 and lower pCO2 may have played a role. Evidence of an expansion of C4 biomass from ~8–6 Ma that peaked in the early to mid Quaternary in Asia, the South Atlantic (ODP Site 1081)65, and Gulf of Mexico (DSDP Site 94)9 implies the possible effect of global decreases in moisture availability along with ice sheet formation and expansion that commenced during the late Miocene, as recorded in δ18O variations preserved in the tests of foraminifera66.
The detailed reconstruction of variations in the prominence of C4 taxa estimated from εPyC-CO2 and εAlk-CO2 is limited by a number of uncertainties, including the effects of changes in atmospheric pCO2 and fractionation due to moisture stress42, and potentially also O2/CO2 ratios67. Notwithstanding these uncertainties, the available evidence is broadly consistent with the conclusion that the rise to prominence of C4 taxa over a substantial proportion of subtropical East Asia differed in timing, likely reflecting local differences in the relative strengths of the suite of potential causal factors. Beginning in the late Pliocene, increased seasonality – with the majority of precipitation occurring during the growing season – and aridity overall could have facilitated sustained increases in the relative abundance of open vegetation in which C4 taxa were important components. As seasonal climate conditions developed and vegetation canopies became more open, expansion of C4 biomass may have become tightly coupled in a feedback relationship with changes in burning regime that effectively selected against shrubs and trees (i.e., C3 species) and promoted further expansions of C4 grasslands15, 64, 68. Lower temperatures associated with continued uplift of central Asia coupled with a strengthened EAWM and the effects of fluctuating Quaternary ice sheets after ~2 Ma could have shifted habitats previously characterized by a relatively high proportion of C4 taxa back within an envelope of environmental conditions that were more suited to C3 species. From around the same time, but at lower altitudinal and subtropical parts of the region, aridity, enhanced by an expanded coverage of ice in the NH and large-scale hydrological dynamics, may have forced an abrupt expansion of C4 biomass29, thereby contributing to a history of C3/C4 variations in East Asia that is unique for the continent and more widely.
Conclusions
The abundance and C isotope composition of n-alkanes and PyC in samples from a sediment core from the northern SCS are used to reconstruct a history of variations in the relative prominence of C4 and C3 taxa in East Asia over the last ~19 Ma. Although present throughout the record, C4 taxa appear to have been prominent at times during the Miocene and especially from the late Pliocene to mid Quaternary. However, n-alkane (a proxy of relative contribution to vegetation) and PyC (a proxy of relative contribution to combusted biomass) isotope data diverge from the mid Quaternary, most likely reflecting differences in provenance, with PyC representing a larger source area that includes higher altitude and more temperate parts of the region, and atmospheric processes as the primary means of transporting organic material from the interior of the continent to the coring site. Temperatures in these already cooler parts of the continent appear to have fallen to levels too low to support C4 taxa from the mid Quaternary. New evidence presented in this paper in the context of existing data indicates that variations in the level and seasonality of rainfall and associated changes in biomass burning appear to have been important drivers of changes in the relative contributions of C3 and C4 taxa to East Asian vegetation throughout much of the past ~19 Ma. An expansion of C4 taxa at low altitude/latitude from ~1 Ma, and especially ~0.4 Ma, may have been triggered by increased aridity and changes to the fire regime. This expansion commenced more recently than in other parts of the tropics and sub-tropics, and is one characteristic of a novel history of variations in C3 and C4 taxa in continental East Asia.
References
Ehleringer, J., Sage, R., Flanagan, L. & Pearcy, R. Climate change and the evolution of C4 photosynthesis. Trends in Ecology and Evolution 6, 95–99 (1991).
Bekker, A. et al. Dating the rise of atmospheric oxygen. Nature 427, 117–120 (2004).
Sage, R. & Stata, M. Photosynthetic diversity meets biodiversity: the C4 plant example. Journal of Plant Physiology 172, 104–119 (2015).
Christin, P., Osborne, C., Sage, R., Arakaki, M. & Edwards, E. C4 eudicots are not younger than C4 monocots. Journal of Experimental Botany 62, 3171–3181 (2011).
Pagani, M., Zachos, J., Freeman, K., Tipple, H. & Bohaty, S. Marked decline in atmospheric carbon dioxide concentrations during the Palaeogene. Science 309, 600–603 (2005).
Grace, J., San, J., Meir, P., Miranda, H. & Montes, R. Productivity and carbon fluxes of tropical savannas. Journal of Biogeography 33, 387–400 (2006).
Quade, J., Cerling, T. & Bowman, J. Development of Asian monsoon revealed by marked ecological shift during the latest Miocene in northern Pakistan. Nature 342, 163–166 (1989).
Cerling, T. et al. Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153–158 (1997).
Tipple, B. & Pagani, M. A 35 Myr North American leaf-wax compound-specific carbon and hydrogen isotope record: implications for C4 grasslands and hydrologic cycle dynamics. Earth and Planetary Science Letters 299, 250–262 (2010a).
Edwards, E., Osborne, C., Strömberg, C., Smith, S. & C4 Grasses Consortium. The origins of C4 grasslands, Integrating evolutionary and ecosystem science. Science 328, 587–591 (2010).
Ding, Z. & Yang, S. C3/C4 vegetation evolution over the last 7.0 Myr in the Chinese Loess Plateau, evidence from pedogenic carbonate δ13C. Palaeogeogr. Palaeoclim. Palaeoecol. 160, 291–299 (2000).
An, Z. et al. Multiple expansions of C4 plant biomass in East Asia since 7 Ma coupled with strengthened monsoon circulation. Geology 33, 705–708 (2005).
Wang, Y. & Deng, T. A 25 m.y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau. Earth and Planetary Science Letters 236, 322–338 (2005).
Passey, B. et al. Strenthened East Asian summer monsoons during a period of high-latitude warmth? Isotopic evidence from Mio-Pliocene fossil mammals and soil carbonates from northern China. Earth and planetary Science Letters 277, 443–452 (2009).
Zhou, B. et al. Late Pliocene-Pleistocene expansion of C4 vegetation in semi-arid East Asia linked to increased burning. Geology 42, 1067–1070 (2014).
Jia, G., Peng, P., Zhao, Q. & Jian, Z. Changes in terrestrial ecosystem since 30 Ma in East Asia: stable isotope evidence from black carbon in the South China Sea. Geology 31, 1093–1096 (2003).
Zhang, Y., Xiong, S., Ding, Z., Lu, H. & Jiang, W. Carbon-oxygen isotope records of pedogenic carbonate from the Early Miocene-Pleistocene loess-red clay in the vicinity of the Liupanshan region and its implications for the early origin of C4 plants in the Chinese Loess Plateau. Quaternary Sciences 31, 800–811 (2011).
Sun, J., Lu, T., Zhang, Z., Wang, X. & Lu, W. Stepwise expansions of C4 biomass and enhanced seasonal precipitation and regional aridity during the Quaternary on the southern Chinese Loess Plateau. Quaternary Science Reviews 34, 57–65 (2012).
Kohn, M. J. Carbon isotope composition of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate. Proceedings of the National Academy of Sciences USA 107, 19691–19695 (2010).
Deines, P. The isotopic composition of reduced organic carbon in Handbook of environmental isotope Geochemistry I, the terrestrial environment (ed. Fritz, P. & Fontes, J. C.) 329–406 (1980).
Liu, W. et al. Carbon isotopic composition of modern soil and paleosol as a response to vegetation change on the Chinese Loess Plateau. Sci China Ser D-Earth Sci 48, 93–99 (2002).
Wang, G., Feng, X., Han, J., Zhou, L., Tan, W. & Su, F. Paleovegetation reconstruction using δ13C of Soil Organic Matter. Biogeosciences 5, 1325–1337 (2008).
Bird, M. & Gröcke, D. Determination of the abundance and carbon isotope composition of elemental carbon in sediments. Geochimica et Cosmochimica Acta 61, 3413–3423 (1997).
Zhou, B. et al. The history of wildfire in the Chinese Loess Plateau during the last 420 ka and its implications to environmental and climate changes. Palaeogeography, Palaeoclimatology, Palaeoecology 252, 617–625 (2007).
Kim, D., Lee, Y., Hyeong, K. & Yoo, C. M. Terrestrial biome distribution in the Late Neogene inferred from a black carbon record in the northeastern equatorial Pacific. Scientific Reports 6, 32847 (2016).
Bird, M. et al. Terrestrial vegetation change inferred from n-alkane 13C analysis in the marine environment. Geochimica et Cosmochimica Acta 59, 2853–2858 (1995).
Cranwell, P. Chain-length distribution of n-alkanes from lake sediments in relation to post-glacial environmental change. Freshwater Biol. 3, 259–265 (1973).
Meyers, P. Applications of organic geochemistry to paleolimnological reconstructions: A summary of Examples from the Laurentian Great Lakes. Organic Geochemistry 34, 261–289 (2003).
Li, L., Li, Q., Tian, J., Wang, H. & Wang, P. Low latitude hydro-climatic changes during the Plio-Pleistocene: evidence from high resolution alkane records in the southern South China Sea. Quaternary Science Reviews 78, 209–224 (2013).
Collister, J., Rieley, G., Stern, B., Eglinton, G. & Fry, B. Compound-specific δ13C analysis of leaf lipids from plants with differing carbon dioxide metabolism. Organic Geochemistry 21, 619–627 (1994).
Zech, M. & Glaser, B. Improved compound-specific δ13C analysis of n-alkanes for application in palaoenvironmental studies. Rapid Communications in Mass Spectrometry 22, 135–142 (2008).
Wang, P. et al. Evolution and variability of the Asian monsoon system: state of the art and outstanding issues. Quaternary Science Reviews 24, 595–629 (2005).
Clift, P., Wan, S. & Bluaxtajn, J. Reconstructing chemical weathering, physical erosion and monsoon intensity since 25 Ma in the northern South China Sea: A review of competing proxies. Earth-Science Reviews 130, 86–102 (2014).
Wan, S., Li, A., Clift, P. & Stuut, J. Development of the East Asian monsoon: Mineralogical and sedimentologic records in the northern South China Sea since 20 Ma. Palaeogeography, Palaeoclimatology, Palaeoecology 254, 561–582 (2007).
Shipboard Scientific Party. Site 1146. Proceedings of the Ocean Drilling Program, Part A: Initial Reports, 184 (2000).
Gustafsson, O. et al. Evaluation of a protocol for the quantification of black carbon in sediments. Global. Biogeochemical Cycles 15, 881–890 (2001).
Zhou, B., Shen, C., Zheng, H. & Zhao, M. Vegetation evolution on the central Chinese Loess Plateau since late Quaternary evidenced by elemental carbon isotopic composition. Chinese Science Bulletin 54, 2082–2089 (2009).
Farquhar, G., Ehleringer, J. & Hubick, K. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Plysiol. Mol. Biol. 40, 503–537 (1989).
Tipple, B., Meyers, S. & Pagani, M. Carbon isotope ratio of Cenozoic CO2: A comparative evaluation of available geochemical proxies. Paleoceanography 25, PA3202 (2010b).
Bird, M. & Ascough, P. Isotopes in pyrogenic carbon: A review. Organic Geochemistry 42, 1529–1539 (2012).
Liu, L., Yang, S., Cui, L. & Hao, Z. Stable carbon isotopic composition of black carbon in surface soil as a proxy for reconstructing vegetation on the Chinese Loess Plateau. Palaeogeography, Palaeoclimatology, Palaeoecology 388, 109–114 (2013).
Schubert, B. & Jahren, H. The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29–43 (2012).
Schubert, B. & Jahren, H. Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435–438 (2015).
Thevenon, F. et al. Combining charcoal and elemental black carbon analysis in sedimentary archives: Implications for past fire regimes, the pyrogenic carbon cycle, and the human–climate interactions. Global & Planetary Change 72, 381–389 (2010).
Ma, P., Gattiker, J., Liu, X. & Rasch, P. A novel approach for determining source-receptor relationships in model simulations: a case study of black carbon transport in northern hemisphere winter. Environmental Research Letters 8, 24042–24049 (2013).
Lee, Y. et al. An integrated approach to identify the biomass burning sources contributing to black carbon epidodes in Hong Kong. Atmospheric Environment 80, 478–487 (2013).
Luo, X. et al. Polycyclic aromatic hydrocarbons in suspended particulate matter and sediments from the Pearl River Estuary and adjacent coastal areas, China. Environmental Pollution 139, 9–20 (2006).
Chuang, M. et al. Aerosol chemical properties and related pollutants measured in Dongsha Island in the northern South China Sea during –SEAS/Dongsha Experiment. Atmospheric Environment 78, 82–92 (2013).
Lin, N. et al. An overview of regional experiments on biomass burning aerosos and related pollutants. Atmopheric Environment 78, 1–19 (2013).
Wang, S. et al. First detailed observations of long-range transported dust over the northern South China Sea. Atmospheric Environment 45, 4804–4808 (2011).
Wang, S. et al. Origin, transport, and vertical distribution of atmospheric pollutants over the northern South China Sea during the 7-SEAS/Dongsha Experiment. Atmospheric Environment 78, 124–133 (2013).
Wang, L. et al. East Asian monsoon climate during the Late Pleistocene: high-resolution sediment records from the South China Sea. Marine Geology 156, 145–284 (1999).
Wu, D. et al. Black carbon over the South China Sea and in various continental locations in South China. Atmos. Chem. Phys. 13, 12257–12270 (2013).
Sun, X. & Li, X. A pollen record of the last 37 ka in deep sea core 17940 from the northern slope of the South China Sea. Marine Geology 156, 227–244 (1999).
Pelejero, C. Terrigenous n-alkane input in the South China Sea: High-resolution records and surface sediments. Chemical Geology 200, 89–103 (2003).
Zhao, Y. et al. Non-polar organic compounds in marine aerosols over the northern South China Sea: Influence of continental outflow. Chemosphere 153, 332–339 (2016).
Zhou, B. et al. Climate and vegetation variations since the LGM recorded by biomarkers from a sediment core in the northern South China Sea. Journal of Quaternary Science 27, 948–955 (2012).
Keeley, J. & Rundel, P. Fire and the Miocene expansion of C4 grasslands. Ecology Letters 8, 683–690 (2005).
Beerling, D. & Royer, D. Convergent Cenozoic CO2 history. Nature Geoscience 4, 418–420 (2011).
An, Z., Kutzbach, J., Prell, W. & Porter, S. Evolution of Asian monsoons and phased uplift of the Himalayan Plateau since Late Miocene times. Nature 411, 62–66 (2001).
Guo, Z. et al. Onset of Asian desertification by 22 Myr ago inferred from loess deposits in china. Nature 416, 159–163 (2002)
Zhang, Z., Zhao, M., Lu, H. & Faiia, A. Lower temperature as the main cause of C4 plant declines during the glacial periods on the Chinese Loess Plateau. Earth and Planetary Science Letters 214, 467–481 (2003).
Freeman, K. & Colarusso, L. Molecular and isotope records of C4 grassland expansion in the late Miocene. Geochimica et Cosmochimica Acta 65, 1439–1454 (2001).
Huang, Y., Clemens, S., Liu, W., Wang, Y. & Prell, W. Large-scale hydrological change drove the late Miocene C4 plant expansion in the Himalayan foreland and Arabian Peninsula. Geology 35, 531–534 (2007).
Hoetzel, S., Dupont, L., Schefuß, E., Rommerskirchen, F. & Wefer, G. The role of fire in Miocene to Pliocene C4 grassland and ecosystem evolution. Nature Geoscience 6, 1027–1030 (2013).
Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K. Trends, rhythms and aberrations in global climate. Science 292, 686–693 (2001).
Beerling, D., Lake, J., Berner, R., Hickey, I., Taylor, D. & Royer, D. Carbon isotope evidence implying high O2/CO2 ratios in the Permo-Carboniferous atmosphere. Geochimica et Cosmochimica Acta 21, 3757–3767 (2002).
Strömberg, C. Evolution of grasses and grassland ecosystems. Annual Review of Earth and Planetary Sciences 39, 517–544 (2011).
Acknowledgements
This research used samples and data provided by the IODP. The authors are grateful to Gangjian Wei, Jia Guodong, Wang Guoan and Liu Weiguo for beneficial discussions and suggested revisions to an earlier version of the paper. We appreciate the assistance of Yuhong Liao, Huashan Chen, Hui Wang, Ling Li, Yingfeng Xu and Yinhua Pan in the laboratory. All δ13CAlk analyses were conducted in the State Key Laboratory of Organic Geochemistry (SKLOG), Guangzhou Institute of Geochemistry, Chinese Academy of Sciences, while δ13CTOC and δ13CPyC analyses were conducted in the State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences. The support of the National Key Basic Research Program of China (2015CB953800), the National Natural Science Foundation of China (41172149, 40802034), Science Fund for Creative Research Groups of NSFC (41321062), State Scholarship Fund of China Scholarship council (No. 2011832365) and the National University of Singapore and James Cook University, Australia, is also gratefully acknowledged.
Author information
Authors and Affiliations
Contributions
Bin Zhou and Hongbo Zheng organized the collection and analysis of samples from core ODP 1146 and designed the research. Bin Zhou, David Taylor and Michael Bird decided upon the focus and scope of this paper. Bin Zhou, David Taylor, Michael Bird and Christopher Wurster drafted the written content of the paper in consultation with the other co-authors. Enlou Zhang measured and analyzed the PyC content of sediment samples, while Luhua Xie measured and analyzed n-alkanes content. All co-authors were involved in interpreting the data and are in agreement with the interpretations arrived at, and conclusions drawn from the research. All co-authors have approved content of the article, and confirm that there are no conflicts of interest relating to the research described.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhou, B., Bird, M., Zheng, H. et al. New sedimentary evidence reveals a unique history of C4 biomass in continental East Asia since the early Miocene. Sci Rep 7, 170 (2017). https://doi.org/10.1038/s41598-017-00285-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-00285-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
