
Abstract
The synthesis of poly(ADP-ribose) (PAR) reconfigures the local chromatin environment and recruits DNA-repair complexes to damaged chromatin. PAR degradation by poly(ADP-ribose) glycohydrolase (PARG) is essential for progression and completion of DNA repair. Here, we show that inhibition of PARG disrupts homology-directed repair (HDR) mechanisms that underpin alternative lengthening of telomeres (ALT). Proteomic analyses uncover a new role for poly(ADP-ribosyl)ation (PARylation) in regulating the chromatin-assembly factor HIRA in ALT cancer cells. We show that HIRA is enriched at telomeres during the G2 phase and is required for histone H3.3 deposition and telomere DNA synthesis. Depletion of HIRA elicits systemic death of ALT cancer cells that is mitigated by re-expression of ATRX, a protein that is frequently inactivated in ALT tumors. We propose that PARylation enables HIRA to fulfill its essential role in the adaptive response to ATRX deficiency that pervades ALT cancers.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Data availability
Original immunofluorescence and colony-formation images are separately available in the Figshare data depository (https://figshare.com/s/17f00c7faa765b329c22). Proteomics data has been deposited at ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD020243. Source data are provided with this paper.
References
Gupte, R., Liu, Z. & Kraus, W. L. PARPs and ADP-ribosylation: recent advances linking molecular functions to biological outcomes. Genes Dev. 31, 101–126 (2017).
Slade, D. et al. The structure and catalytic mechanism of a poly(ADP-ribose) glycohydrolase. Nature 477, 616–620 (2011).
Koh, D. W. et al. Failure to degrade poly(ADP-ribose) causes increased sensitivity to cytotoxicity and early embryonic lethality. Proc. Natl Acad. Sci. USA 101, 17699–17704 (2004).
Hanzlikova, H. et al. The importance of poly(ADP-ribose) polymerase as a sensor of unligated Okazaki fragments during DNA replication. Mol. Cell 71, 319–331.e3 (2018).
Maya-Mendoza, A. et al. High speed of fork progression induces DNA replication stress and genomic instability. Nature 559, 279–284 (2018).
García-Expósito, L. et al. Proteomic profiling reveals a specific role for translesion DNA polymerase η in the alternative lengthening of telomeres. Cell Rep. 17, 1858–1871 (2016).
Schmutz, I., Timashev, L., Xie, W., Patel, D. J. & de Lange, T. TRF2 binds branched DNA to safeguard telomere integrity. Nat. Struct. Mol. Biol. 24, 734–742 (2017).
Mateos-Gomez, P. A. et al. Mammalian polymerase θ promotes alternative NHEJ and suppresses recombination. Nature 518, 254–257 (2015).
Doksani, Y. & de Lange, T. Telomere-internal double-strand breaks are repaired by homologous recombination and PARP1/Lig3-dependent end-joining. Cell Rep. 17, 1646–1656 (2016).
Dunham, M. A., Neumann, A. A., Fasching, C. L. & Reddel, R. R. Telomere maintenance by recombination in human cells. Nat. Genet. 26, 447–450 (2000).
Dilley, R. L. et al. Break-induced telomere synthesis underlies alternative telomere maintenance. Nature 539, 54–58 (2016).
Roumelioti, F.-M. et al. Alternative lengthening of human telomeres is a conservative DNA replication process with features of break-induced replication. EMBO Rep. 17, 1731–1737 (2016).
James, D. I. et al. First-in-class chemical probes against poly(ADP-ribose) glycohydrolase (PARG) inhibit DNA repair with differential pharmacology to olaparib. ACS Chem. Biol. 11, 3179–3190 (2016).
Yeager, T. R. et al. Telomerase-negative immortalized human cells contain a novel type of promyelocytic leukemia (PML) body. Cancer Res. 59, 4175–4179 (1999).
Londoño-Vallejo, J. A., Der-Sarkissian, H., Cazes, L., Bacchetti, S. & Reddel, R. R. Alternative lengthening of telomeres is characterized by high rates of telomeric exchange. Cancer Res. 64, 2324–2327 (2004).
O’Sullivan, R. J. et al. Rapid induction of alternative lengthening of telomeres by depletion of the histone chaperone ASF1. Nat. Struct. Mol. Biol. 21, 167–174 (2014).
Berti, M. et al. Human RECQ1 promotes restart of replication forks reversed by DNA topoisomerase I inhibition. Nat. Struct. Mol. Biol. 20, 347–354 (2013).
Clynes, D. et al. Suppression of the alternative lengthening of telomere pathway by the chromatin remodelling factor ATRX. Nat. Commun. 6, 7538–11 (2015).
Cho, N. W., Dilley, R. L., Lampson, M. A. & Greenberg, R. A. Interchromosomal homology searches drive directional ALT telomere movement and synapsis. Cell 159, 108–121 (2014).
Mortusewicz, O., Fouquerel, E., Amé, J.-C., Leonhardt, H. & Schreiber, V. PARG is recruited to DNA damage sites through poly(ADP-ribose)- and PCNA-dependent mechanisms. Nucleic Acids Res. 39, 5045–5056 (2011).
Kaufmann, T. et al. A novel non-canonical PIP-box mediates PARG interaction with PCNA. Nucleic Acids Res. 45, 9741–9759 (2017).
Zhang, J.-M., Yadav, T., Ouyang, J., Lan, L. & Zou, L. Alternative lengthening of telomeres through two distinct break-induced replication pathways. Cell Rep. 26, 955–968.e3 (2019).
Illuzzi, G. et al. PARG is dispensable for recovery from transient replicative stress but required to prevent detrimental accumulation of poly(ADP-ribose) upon prolonged replicative stress. Nucleic Acids Res. 42, 7776–7792 (2014).
Gagné, J.-P. et al. Proteome-wide identification of poly(ADP-ribose) binding proteins and poly(ADP-ribose)-associated protein complexes. Nucleic Acids Res. 36, 6959–6976 (2008).
Gagné, J.-P. et al. Quantitative proteomics profiling of the poly(ADP-ribose)-related response to genotoxic stress. Nucleic Acids Res. 40, 7788–7805 (2012).
Jungmichel, S. et al. Proteome-wide identification of poly(ADP-ribosyl)ation targets in different genotoxic stress responses. Mol. Cell 52, 272–285 (2013).
Karras, G. I. et al. The macro domain is an ADP-ribose binding module. EMBO J. 24, 1911–1920 (2005).
Ahel, D. et al. Poly(ADP-ribose)-dependent regulation of DNA repair by the chromatin remodeling enzyme ALC1. Science 325, 1240–1243 (2009).
Gomez, M. et al. PARP1 is a TRF2-associated poly(ADP-ribose)polymerase and protects eroded telomeres. Mol. Biol. Cell 17, 1686–1696 (2006).
Masson, M. et al. XRCC1 is specifically associated with poly(ADP-ribose) polymerase and negatively regulates its activity following DNA damage. Mol. Cell. Biol. 18, 3563–3571 (1998).
Altmeyer, M. et al. The chromatin scaffold protein SAFB1 renders chromatin permissive for DNA damage signaling. Mol. Cell 52, 206–220 (2013).
Acs, K. et al. The AAA-ATPase VCP/p97 promotes 53BP1 recruitment by removing L3MBTL1 from DNA double-strand breaks. Nat. Struct. Mol. Biol. 18, 1345–1350 (2011).
Flynn, R. L. et al. TERRA and hnRNPA1 orchestrate an RPA-to-POT1 switch on telomeric single-stranded DNA. Nature 471, 532–536 (2011).
Gagné, J.-P., Hunter, J. M., Labrecque, B., Chabot, B. & Poirier, G. G. A proteomic approach to the identification of heterogeneous nuclear ribonucleoproteins as a new family of poly(ADP-ribose)-binding proteins. Biochem. J. 371, 331–340 (2003).
Polo, S. E. et al. Regulation of DNA-end resection by hnRNPU-like proteins promotes DNA double-strand break signaling and repair. Mol. Cell 45, 505–516 (2012).
Adamson, B., Smogorzewska, A., Sigoillot, F. D., King, R. W. & Elledge, S. J. A genome-wide homologous recombination screen identifies the RNA-binding protein RBMX as a component of the DNA-damage response. Nat. Cell Biol. 14, 318–328 (2012).
Altmeyer, M. et al. Liquid demixing of intrinsically disordered proteins is seeded by poly(ADP-ribose). Nat. Commun. 6, 8088–12 (2015).
Singatulina, A. S. et al. PARP-1 activation directs FUS to DNA damage sites to form PARG-reversible compartments enriched in damaged DNA. Cell Rep. 27, 1809–1821.e5 (2019).
Schrank, B. R. et al. Nuclear ARP2/3 drives DNA break clustering for homology-directed repair. Nature 559, 61–66 (2018).
Tagami, H., Ray-Gallet, D., Almouzni, G. & Nakatani, Y. Histone H3.1 and H3.3 complexes mediate nucleosome assembly pathways dependent or independent of DNA synthesis. Cell 116, 51–61 (2004).
Goldberg, A. D. et al. Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell 140, 678–691 (2010).
Pchelintsev, N. A. et al. Placing the HIRA histone chaperone complex in the chromatin landscape. Cell Rep. 3, 1012–1019 (2013).
Adam, S., Polo, S. E. & Almouzni, G. Transcription recovery after DNA damage requires chromatin priming by the H3.3 histone chaperone HIRA. Cell 155, 94–106 (2013).
Lewis, P. W., Elsaesser, S. J., Noh, K.-M., Stadler, S. C. & Allis, C. D. Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres. Proc. Natl Acad. Sci. USA 107, 14075–14080 (2010).
Juhász, S., Elbakry, A., Mathes, A. & Löbrich, M. ATRX promotes DNA repair synthesis and sister chromatid exchange during homologous recombination. Mol. Cell 71, 11–24.e7 (2018).
Heaphy, C. M. et al. Altered telomeres in tumors with ATRX and DAXX mutations. Science 333, 425–425 (2011).
Lovejoy, C. A. et al. Loss of ATRX, genome instability, and an altered DNA damage response are hallmarks of the alternative lengthening of telomeres pathway. PLoS Genet. 8, e1002772 (2012).
Zhang, H. et al. RPA interacts with HIRA and regulates H3.3 deposition at gene regulatory elements in mammalian cells. Mol. Cell 65, 272–284 (2017).
Ray-Gallet, D. et al. Dynamics of histone H3 deposition in vivo reveal a nucleosome gap-filling mechanism for H3.3 to maintain chromatin integrity. Mol. Cell 44, 928–941 (2011).
Elsässer, S. J. et al. DAXX envelops a histone H3.3-H4 dimer for H3.3-specific recognition. Nature 491, 560–565 (2012).
Episkopou, H. et al. Alternative lengthening of telomeres is characterized by reduced compaction of telomeric chromatin. Nucleic Acids Res. 42, 4391–4405 (2014).
Tang, Y. et al. Structure of a human ASF1a–HIRA complex and insights into specificity of histone chaperone complex assembly. Nat. Struct. Mol. Biol. 13, 921–929 (2006).
Ask, K. et al. Codanin-1, mutated in the anaemic disease CDAI, regulates Asf1 function in S-phase histone supply. EMBO J. 31, 2013–2023 (2012).
Acknowledgements
We are indebted to M. Lund Nielsen for sharing detailed protocols and advice in relation to enrichment of PARylated proteins with AF1521. We are grateful to M. Altmeyer, D. Clynes, R. Greenberg, B. Johnson, K. Miller, S. Oesterreich, S. Polo, D. Slade, H. van Attikum and S. Smith for sharing high quality reagents. We thank G. Delgoffe, A. Menk and J. Stewart-Ornstein for guidance with experiments and sharing resources. Resources and facilities at the UPMC Hillman Cancer Center were supported by Comprehensive Cancer Center Support Grant NCI/no. P30CA047904. Research funding was provided to individual investigators from the following agencies; R.J.O. NCI/no. 5R01CA207209-02 and American Cancer Society no. RSG-18-038-01-DMC; S.C.W. NIH/no. 1S10OD019973-O1; A.I.N. NIH U24CA210967 and R01 GM094231; R.W.S. NIH/no. R01CA148629 and NIEHS/no. R01ES014811. N.K. is supported by a T32 training grant, NIGMS/no. T32GM008424-25. This work was also supported by grants C480/A11411 and C5759/A17098 from Cancer Research UK to D.I.J., I.D.W., K.M.S and D.O.; and ERC-2015-ADG- 694694 ‘ChromADICT’, ANR-16-CE15-0018 ‘CHRODYT’, ANR-16-CE11-0028 ‘REPLICAF’, ANR-16-CE12-0024 ‘CHIFT’, ITN-765966 Curie ‘EPISYSTEM’ and ITN-813327 Curie ‘CHROMDESIGN’ to G.A. and D.R.G.
Author information
Authors and Affiliations
Contributions
R.J.O. conceived and designed the study; S.M.H. designed and conducted all experimentation; R.J.O. and S.M.H analyzed the data. N.K., R.B., M.L.L., J.L.R., J.B.G., L.G.-E., A.R.W. and R.J.O. assisted with experimentation and analyzed data. C.T.W. and S.C.W. assisted with optical-imaging experiment design, optimization and analyzes. D.I.J., I.D.W., K.M.S. and D.O. produced, validated and provided the PARG inhibitors. F.V.L., D.M. and A.I.N. generated and analyzed proteomic data. J.L. and R.W.S. provided essential reagents. D.R.-G. and G.A. provided essential reagents, assisted with experimental design and interpretation of results. S.M.H. and R.J.O. wrote the manuscript with input from coauthors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Beth Moorefield was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
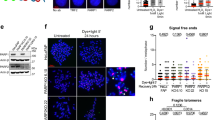
Extended Data Fig. 1 Disruption of PAR turnover perturbs recombinogenic activity at ALT telomeres.
a, Western blot of PARP1 or PARG knockdown and PAR accumulation in U2OS cells expressing shRNAs. Extracts from 2 mM hydroxyurea (HU)-PARGi (5μM/24hrs) treated U2OS cells serve as a control for PAR induction. b, Representative IF images of APBs (PML-TTAGGG) in U2OS cells expressing the indicated shRNAs. c, Quantification of APBs (% positive cells) in U2OS, Saos2 and HeLa LT cells expressing the indicated shRNAs. d, Quantification of telomere sister chromatid exchanges (t-SCE) (% per metaphase) in U2OS, Saos2 and HeLa LT cells expressing the indicated shRNAs. n refers to the number of metaphase spreads analyzed from N = 3 independent assays. e, Left: Western blot of U2OS cells transfected with non-targeting and/or RECQ1 siRNAs treated with PARPi (5μM/72hrs). Right: Quantification of t-SCEs in U2OS and HeLa LT cells transfected with NT siRNA, and/or RECQ1 siRNA. DMSO/PARPi (5μM/24hrs). n refers to the number of metaphase spreads analyzed from N = 4 independent assays. f, Cell cycle profiles and (g) cellular [ADP/ATP] ratio of U2OS and Hela LT cells treated with inhibitors for 12 days. h, PFGE of DMSO, PARPi (100 nM) or PARGi (1 μM) treated VA13 cells. The red dot indicates mean telomere length (kb). i, Representative images and quantification from clonogenic survival assays in ALT + cells, TEL + cells and (j) U2OS cells expressing ATRX treated with DMSO, PARPi (100μM) or PARGi (1μM) for 7 days. All scale bars in IF panels=5μm. All graphed data in the figure are mean ± s.e.m except d, e which are mean ± s.d. Unless otherwise stated, (n) is the number of cells analyzed and the number of independent assays (N) conducted is represented by black circles. Statistical significance was determined using one-way ANOVA except except (d) where the Mann-Whitney test was used and (i-j) where Students t test was used. Uncropped blots for a, e and digital images are deposited on Figshare. Graphed data is available as Source Data.
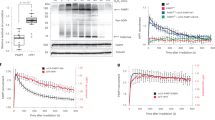
Extended Data Fig. 2 PARylation is an early and direct mediator of TRF1-FokI DSB formation.
a, [ADP/ATP] ratio in DMSO/PARGi treated WT-TRF1-FokI U2OS cells. Cells were treated with 1.5 mM/1 hr MMS. (b) Representative IF images and quantification of PAR at WT-TRF1-FokI DSBs after PARGi, PARGi-PARPi or TNKS1 knockdown. c, Representative IF images and quantification showing GFP-PARP1 localization in WT-TRF1-FokI cells treated with PARPi, PARGi or both. d, Representative IF images and quantification showing GFP-PARG localization in WT-TRF1-FokI U2OS cells. e, Left: Representative IF images and quantification of telomere foci size per cell in VA13 and Hela LT cells transfected with WT-TRF1-FokI from N = 2 independent assays. f, Representative stills of telomere (eGFP-TRF1) movement in U2OS cells treated with DMSO, PARPi or PARGi. Graph displays the cumulative Mean Squared Displacement (MSD) of 100 telomeres. g, Top: Schematic of DNA combing in G2-synchronized WT-TRF1-FokI cells treated with DMSO, PARPi, PARGi, or co-treated with PARPi and PARGi. Left: Quantification of telomeric fiber length of combined pulses. Right: Violin plot analysis of fork velocity. h, Graphs of CldU/IdU tract distribution of telomeric fibers in inhibitor treated U2OS-TRF1-FokI cells. n refers to the number of fibers containing TTAGGG signals analyzed from N = 2 independent assays. i, Representative IF images and quantification of BrdU synthesis at telomeres in the indicated cell lines after transfection with WT-TRF1-FokI and treated with PARGi or PARPi. j, Representative IF images and quantification of PCNA and (k) POLD3 localization at WT-TRF1-FokI telomeres treated with treated with DMSO, PARPi, PARGi, PARGi-Me or PARPi-PARGi. All inhibitor treatments, 5μM/4hrs unless otherwise indicated. All scale bars in IF panels=5μm. All graphed data in the figure are mean ± s.e.m. Unless otherwise stated, (n) is the number of cells analyzed and the number of independent assays (N) conducted is represented by black circles. Statistical significance was determined using one-way ANOVA. Digital images are deposited on Figshare. Graphed data is available as Source Data.
Extended Data Fig. 3 PAR-dependent recruitment of proteins to TRF1-FokI induced telomeric DSBs.
a, Spectral counts for the indicated proteins that were identified by Af1521-PAR proteomics. b, Western blot analysis illustrating the expression of GFP fusion proteins in U2OS WT-TRF1-FokI cells. GFP antibody was used to blot for protein expression in each treatment. c, Representative IF images showing the localization of the indicated GFP fusion proteins in WT-TRF1-FokI U2OS cells following treatment with DMSO, PARPi and combined PARGi/PARPi. d, In vivo PARylation assay with GFP tagged FUS, RBMX and ARP3. * indicates the band corresponding to the immunoprecipitated GFP-tagged target protein on PAR blots. e, Western blotting was performed with antibodies to validate siRNA knockdown of endogenous protein hits from Af1521-PAR proteomics in U2OS cells. f, Cell cycle profile of U2OS and VA13 cells after siRNA knockdown of protein hits from Af1521-PAR proteomics. All inhibitor treatments, 5μM/4hrs. All scale bars in IF panels=5μm. Uncropped blots for b, d-e and digital images are deposited on Figshare. Graphed data is available as Source Data.
Extended Data Fig. 4 Selectivity of HIRA for localization to telomeres in ALT cancer cells telomeric DSBs is independent of RPA and is necessary for telomere DNA synthesis.
a, Representative IF images of HIRA-YFP localization in ALT + and TEL + cell lines treated with DMSO/PARGi. b, Representative IF images of HIRA-YFP localization in U2OS cells after exposure to 30 J/m2ultra-violet C (UV-C) and 10 Gy ionizing irradiation (γIR). 5 μM PARGi was added for 30 mins following irradiation. c, Left: Western blot validation of RPA70 knockdown in U2OS cells. Middle: Representative IF images of HIRA-YFP localization at telomeres in U2OS cells after RPA70 knockdown. Right: Quantification of HIRA-YFP localization to telomeres in indicated conditions from N = 2 independent assays. d, Western blot validation of HIRA, CABIN1 and UBN1 siRNA knockdown in U2OS cells. e, Graphs of CldU/IdU tract distribution of >30 telomeric fibers in NT siRNA and HIRA siRNA transfected U2OS-TRF1-FokI cells. f, Representative IF images and quantification of BrdU synthesis at telomeres in indicated cell lines that are transfected with WT-TRF1-FokI and HIRA siRNA. All inhibitor treatments, 5 μM/4 hrs. All scale bars in IF panels=5 μm. Unless otherwise stated, (n) is the number of cells analyzed and the number of independent assays (N) conducted is represented by black circles. Uncropped blots for c-d and digital images are deposited on Figshare. Graphed data is available as Source Data.
Extended Data Fig. 5 HIRA compensates for and forms a synthetic lethal interaction with ATRX loss.
a, Representative IF images and quantification of HIRA-YFP localization in Hela LT cells transfected with ATRX siRNA and WT-TRF1-FokI, as well as treated with PARGi (5 μM, from N = 2 independent assays. b, Western blot validation of HIRA knockdown in ALT + and TEL + cell lines using two different shRNA sequences (#1 and #2). c, Western blot validation of HIRA and ATRX knockdown in TEL + HeLa LT cell lines using the indicated shRNAs. d, Representative images and quantification of clonogenic survival assays with Hela LT cell line stably expressing scrambled non-targeting (NT), ATRX (#A and #B), and HIRA (#1 and #2) shRNAs for 5 days. All inhibitor treatments, 5μM/4hrs. All scale bars in IF panels=5μm. Unless otherwise stated, (n) is the number of cells analyzed and the number of independent assays (N) conducted is represented by black circles. Uncropped blots for c-d and digital images are deposited on Figshare. Graphed data is available as Source Data.
Extended Data Fig. 6 Analysis of potential binding of PAR by HIRA.
a, GFP-PARP1, YFP-HIRA or GFP-FUS were transiently transfected in U2OS cells. Immunoprecipitated and blotted GFP-fusion proteins were incubated with biotinylated PAR and detected using an anti-PAR (10H) antibody. GFP fusion proteins were detected with HRP conjugated GFP antibody. b, Western blot showing depletion of endogenous HIRA and complementation with the indicated HIRA constructs in U2OS cells. Uncropped blot images for panel a,b are shown in Supplementary Fig. 1.
Supplementary information
Supplementary Information
Supplementary Figure 1.
Supplementary Video 1
Telomere mobility in ALT U2OS cells treated with DMSO.
Supplementary Video 2
Telomere mobility in ALT U2OS cells treated with PARPi.
Supplementary Video 3
Telomere mobility in ALT U2OS cells treated with PARGi.
Supplementary Table 1
Listing of proteins identified by mass spectrometry.
Source data
Source Data Fig. 1
Source data for graphs
Source Data Fig. 2
Source data for graphs
Source Data Fig. 3
Source data for graphs
Source Data Fig. 4
Source data for graphs
Source Data Fig. 5
Source data for graphs
Source Data Fig. 6
Source data for graphs
Source Data Extended Data Fig. 1
Source data for graphs
Source Data Extended Data Fig. 2
Source data for graphs
Source Data Extended Data Fig. 3
Source data for graphs
Source Data Extended Data Fig. 4
Source data for graphs
Source Data Extended Data Fig. 5
Source data for graphs
Rights and permissions
About this article

Cite this article
Hoang, S.M., Kaminski, N., Bhargava, R. et al. Regulation of ALT-associated homology-directed repair by polyADP-ribosylation. Nat Struct Mol Biol 27, 1152–1164 (2020). https://doi.org/10.1038/s41594-020-0512-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41594-020-0512-7
This article is cited by
-
PARP2 promotes Break Induced Replication-mediated telomere fragility in response to replication stress
Nature Communications (2024)
-
HIRA vs. DAXX: the two axes shaping the histone H3.3 landscape
Experimental & Molecular Medicine (2024)
-
Signalling inhibition by ponatinib disrupts productive alternative lengthening of telomeres (ALT)
Nature Communications (2023)
-
Regulation of Rad52-dependent replication fork recovery through serine ADP-ribosylation of PolD3
Nature Communications (2023)
-
Functions of ADP-ribose transferases in the maintenance of telomere integrity
Cellular and Molecular Life Sciences (2022)
