
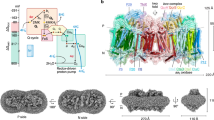
Abstract
In the mycobacterial electron-transport chain, respiratory complex III passes electrons from menaquinol to complex IV, which in turn reduces oxygen, the terminal acceptor. Electron transfer is coupled to transmembrane proton translocation, thus establishing the electrochemical proton gradient that drives ATP synthesis. We isolated, biochemically characterized, and determined the structure of the obligate III2IV2 supercomplex from Mycobacterium smegmatis, a model for Mycobacterium tuberculosis. The supercomplex has quinol:O2 oxidoreductase activity without exogenous cytochrome c and includes a superoxide dismutase subunit that may detoxify reactive oxygen species produced during respiration. We found menaquinone bound in both the Qo and Qi sites of complex III. The complex III-intrinsic diheme cytochrome cc subunit, which functionally replaces both cytochrome c1 and soluble cytochrome c in canonical electron-transport chains, displays two conformations: one in which it provides a direct electronic link to complex IV and another in which it serves as an electrical switch interrupting the connection.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


References
Bald, D., Villellas, C., Lu, P. & Koul, A. Targeting energy metabolism in Mycobacterium tuberculosis, a new paradigm in antimycobacterial drug discovery. MBio 8, e00272–17 (2017).
Sarewicz, M. & Osyczka, A. Electronic connection between the quinone and cytochrome C redox pools and its role in regulation of mitochondrial electron transport and redox signaling. Physiol. Rev. 95, 219–243 (2015).
Zhang, Z. et al. Electron transfer by domain movement in cytochrome bc 1. Nature 392, 677–684 (1998).
Darrouzet, E., Moser, C. C., Dutton, P. L. & Daldal, F. Large scale domain movement in cytochrome bc(1): a new device for electron transfer in proteins. Trends Biochem. Sci. 26, 445–451 (2001).
Berry, E. A., De Bari, H. & Huang, L. S. Unanswered questions about the structure of cytochrome bc1 complexes. Biochim. Biophys. Acta 1827, 1258–1277 (2013).
Wikström, M., Sharma, V., Kaila, V. R. I., Hosler, J. P. & Hummer, G. New perspectives on proton pumping in cellular respiration. Chem. Rev. 115, 2196–2221 (2015).
Lee, H. J., Reimann, J., Huang, Y. & Ädelroth, P. Functional proton transfer pathways in the heme-copper oxidase superfamily. Biochim. Biophys. Acta 1817, 537–544 (2012).
Megehee, J. A., Hosler, J. P. & Lundrigan, M. D. Evidence for a cytochrome bcc-aa3 interaction in the respiratory chain of Mycobacterium smegmatis. Microbiology 152, 823–829 (2006).
Kao, W.-C. et al. The obligate respiratory supercomplex from Actinobacteria. Biochim. Biophys. Acta 1857, 1705–1714 (2016).
Niebisch, A. & Bott, M. Purification of a cytochrome bc-aa3 supercomplex with quinol oxidase activity from Corynebacterium glutamicum: identification of a fourth subunity of cytochrome aa3 oxidase and mutational analysis of diheme cytochrome c1. J. Biol. Chem. 278, 4339–4346 (2003).
Graf, S. et al. Rapid electron transfer within the III-IV supercomplex in Corynebacterium glutamicum. Sci. Rep. 6, 34098 (2016).
Matsoso, L. G. et al. Function of the cytochrome bc1-aa3 branch of the respiratory network in mycobacteria and network adaptation occurring in response to its disruption. J. Bacteriol. 187, 6300–6308 (2005).
Sone, N. et al. A novel hydrophobic diheme c-type cytochrome: purification from Corynebacterium glutamicum and analysis of the QcrCBA operon encoding three subunit proteins of a putative cytochrome reductase complex. Biochim. Biophys. Acta 1503, 279–290 (2001).
Björck, M. L. et al. Reaction of S. cerevisiae mitochondria with ligands: kinetics of CO and O2 binding to flavohemoglobin and cytochrome c oxidase. Biochim. Biophys. Acta 1858, 182–188 (2017).
Hunte, C., Koepke, J., Lange, C., Rossmanith, T. & Michel, H. Structure at 2.3 A resolution of the cytochrome bc(1) complex from the yeast Saccharomyces cerevisiae co-crystallized with an antibody Fv fragment. Structure 8, 669–684 (2000).
Lange, C. & Hunte, C. Crystal structure of the yeast cytochrome bc1 complex with its bound substrate cytochrome c. Proc. Natl Acad. Sci. USA 99, 2800–2805 (2002).
Ädelroth, P., Ek, M. S., Mitchell, D. M., Gennis, R. B. & Brzezinski, P. Glutamate 286 in cytochrome aa3 from Rhodobacter sphaeroides is involved in proton uptake during the reaction of the fully-reduced enzyme with dioxygen. Biochemistry 36, 13824–13829 (1997).
Yoshikawa, S. et al. Redox-coupled crystal structural changes in bovine heart cytochrome c oxidase. Science 280, 1723–1729 (1998).
Ostermeier, C., Harrenga, A., Ermler, U. & Michel, H. Structure at 2.7 A resolution of the Paracoccus denitrificans two-subunit cytochrome c oxidase complexed with an antibody FV fragment. Proc. Natl Acad. Sci. USA 94, 10547–10553 (1997).
Hosler, J. P. The influence of subunit III of cytochrome c oxidase on the D pathway, the proton exit pathway and mechanism-based inactivation in subunit I. Biochim. Biophys. Acta 1655, 332–339 (2004).
Svensson-Ek, M. et al. The X-ray crystal structures of wild-type and EQ(I-286) mutant cytochrome c oxidases from Rhodobacter sphaeroides. J. Mol. Biol. 321, 329–339 (2002).
Sun, C. et al. Structure of the alternative complex III in a supercomplex with cytochrome oxidase. Nature 557, 123–126 (2018).
Qin, L., Sharpe, M. A., Garavito, R. M. & Ferguson-Miller, S. Conserved lipid-binding sites in membrane proteins: a focus on cytochrome c oxidase. Curr. Opin. Struct. Biol. 17, 444–450 (2007).
Wenz, T. et al. Role of phospholipids in respiratory cytochrome bc1 complex catalysis and supercomplex formation. Biochim. Biophys. Acta 1787, 609–616 (2009).
Smirnova, I. A., Ädelroth, P., Gennis, R. B. & Brzezinski, P. Aspartate-132 in cytochrome c oxidase from Rhodobacter sphaeroides is involved in a two-step proton transfer during oxo-ferryl formation. Biochemistry 38, 6826–6833 (1999).
Schnackenberg, J. et al. Amino acid sequence, crystallization and structure determination of reduced and oxidized cytochrome c6 from the green alga Scenedesmus obliquus. J. Mol. Biol. 290, 1019–1030 (1999).
Spagnolo, L. et al. Unique features of the sodC-encoded superoxide dismutase from Mycobacterium tuberculosis, a fully functional copper-containing enzyme lacking zinc in the active site. J. Biol. Chem. 279, 33447–33455 (2004).
Jenney, F. E. Jr., Verhagen, M. F. J. M., Cui, X. & Adams, M. W. W. Anaerobic microbes: oxygen detoxification without superoxide dismutase. Science 286, 306–309 (1999).
Vygodina, T. V., Pecoraro, C., Mitchell, D., Gennis, R. & Konstantinov, A. A. Mechanism of inhibition of electron transfer by amino acid replacement K362M in a proton channel of Rhodobacter sphaeroides cytochrome c oxidase. Biochemistry 37, 3053–3061 (1998).
Lundgren, C. A. K. et al. Scavenging of superoxide by a membrane-bound superoxide oxidase. Nat. Chem. Biol. 14, 788–793 (2018).
Ojemyr, L. N., Lee, H. J., Gennis, R. B. & Brzezinski, P. Functional interactions between membrane-bound transporters and membranes. Proc. Natl Acad. Sci. USA 107, 15763–15767 (2010).
Marantz, Y., Nachliel, E., Aagaard, A., Brzezinski, P. & Gutman, M. The proton collecting function of the inner surface of cytochrome c oxidase from Rhodobacter sphaeroides. Proc. Natl Acad. Sci. USA 95, 8590–8595 (1998).
Gray, H. B. & Winkler, J. R. Long-range electron transfer. Proc. Natl Acad. Sci. USA 102, 3534–3539 (2005).
Parikh, A. et al. Development of a new generation of vectors for gene expression, gene replacement, and protein-protein interaction studies in mycobacteria. Appl. Environ. Microbiol. 79, 1718–1729 (2013).
Thomas, P. E., Ryan, D. & Levin, W. An improved staining procedure for the detection of the peroxidase activity of cytochrome P-450 on sodium dodecyl sulfate polyacrylamide gels. Anal. Biochem. 75, 168–176 (1976).
Goodhew, C. F., Brown, K. R. & Pettigrew, G. W. Haem staining in gels, a useful tool in the study of bacterial c-type cytochromes. Biochim. Biophys. Acta 852, 288–294 (1986).
Barr, I. & Guo, F. Pyridine hemochromagen assay for determining the concentration of heme in purified protein solutions. Bio Protoc. 5, e1594 (2015).
Berry, E. A. & Trumpower, B. L. Simultaneous determination of hemes a, b, and c from pyridine hemochrome spectra. Anal. Biochem. 161, 1–15 (1987).
Wittig, I., Braun, H. P. & Schägger, H. Blue native PAGE. Nat. Protoc. 1, 418–428 (2006).
Kim, M. S. et al. Isolation and characterization of a hybrid respiratory supercomplex consisting of mycobacterium tuberculosis cytochrome bcc and mycobacterium smegmatis cytochrome aa3. J. Biol. Chem. 290, 14350–14360 (2015).
Brandt, U., Schägger, H. & von Jagow, G. Purification of cytochrome-c oxidase retaining its pulsed form. Eur. J. Biochem. 182, 705–711 (1989).
Kuo, W.-Y., Huang, C.-H., Shih, C. & Jinn, T.-L. Cellular extract preparation for superoxide dismutase (SOD) activity assay. Bio Protoc. 3, e811 (2013).
Wu, C. H. H. et al. Identification and subcellular localization of a novel Cu,Zn superoxide dismutase of Mycobacterium tuberculosis. FEBS Lett. 439, 192–196 (1998).
Rubinstein, J. L. & Brubaker, M. A. Alignment of cryo-EM movies of individual particles by optimization of image translations. J. Struct. Biol. 192, 188–195 (2015).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Bai, X. C., Rajendra, E., Yang, G., Shi, Y. & Scheres, S. H. W. Sampling the conformational space of the catalytic subunit of human γ-secretase. eLife 4, e11182 (2015).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Drozdetskiy, A., Cole, C., Procter, J. & Barton, G. J. JPred4: a protein secondary structure prediction server. Nucleic Acids Res. 43(W1), W389–W394 (2015).
Omasits, U., Ahrens, C. H., Müller, S. & Wollscheid, B. Protter: interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 30, 884–886 (2014).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66, (213–221 (2010).
Pettersen, E. F. et al. UCSF Chimera: a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12–21 (2010).
Barad, B. A. et al. EMRinger: side chain-directed model and map validation for 3D cryo-electron microscopy. Nat. Methods 12, 943–946 (2015).
Bienert, S. et al. The SWISS-MODEL Repository: new features and functionality. Nucleic Acids Res. 45(D1), D313–D319 (2017).
Acknowledgements
The C. glutamicum and M. smegmatis strains were provided by M. Bott (Forschungszentrum Jülich) and B. Kana (University of Witwatersrand), respectively. V. Nandicoori (NII, New Delhi) provided the PST-KT plasmid. We thank S. Graf for help with purification of the C. glutamicum supercomplex, and A. Osyczka and E. Berry for valuable discussions. This work was supported by Knut and Alice Wallenberg Foundation (KAW) grants 2013.0006 (P.B., M.H., and P.Ä.) and 2017.0275 (M.H.), and Canadian Institutes of Health Research grant MOP81294 and the Canada Research Chairs program (J.L.R.). The Mass Spectrometry Based Proteomic Facility (Uppsala University), the TEM facility at Stockholm University, and the cryo-EM facility at the Science for Life Laboratory provided the infrastructure. Titan Krios cryo-EM data were collected at the Toronto High-Resolution High-Throughput Cryo-EM Facility, supported by the Canada Foundation for Innovation and Ontario Research Fund.
Author information
Authors and Affiliations
Contributions
R.G.N., P.Ä., M.H., and P.B. conceived and designed the research. R.G.N., O.F., and D.S. performed protein purification, and biochemical and enzymatic analyses. J.S. collected spectral and kinetic data. R.G.N. and Q.K. performed negative-stain EM analyses. B.W. and R.G.N. performed cryo-EM sample preparation and initial data collection and analysis. S.B. collected the final cryo-EM data and performed image analysis. H.G. performed various calculations with the cryo-EM data. B.W. built the de novo atomic structures. J.L.R. supervised the cryo-EM analysis. P.B., M.H., P.Ä., and J.L.R. interpreted data. P.B., J.L.R., and M.H. wrote the manuscript. B.W., M.H., J.S., and J.L.R. prepared data figures and legends.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Purification of the intact supercomplex (SC).
a. The SC was purified via a 3×FLAG tag on QcrB. b. Purification profile on 4-16% gradient SDS PAGE. Protein bands were identified by mass spectrometry as indicated. The bands marked by an asterisk (*) showed lower MS scores. Markers (kDa) are indicated. c. SC purified via the 3×FLAG-tag was subjected to SEC. The sample is highly homogenous as evident by the single peak. The elution volumes for the SEC standards Thyroglobulin (669 kDa), Apoferritin (443 kDa) and Amylase (200 kDa) were used to estimate the size of the SC (~970 kDa), as shown in the inset. d. 4-16 % gradient SDS-PAGE after SEC. e. SEC peak fractions were pooled and analyzed with BN-PAGE. Native markers are indicated. f. In order to obtain highly pure sample for mass spectrometry the BN-PAGE band was cut out and run on an SDS-PAGE. Novel components identified by MS are indicated on the left. The bands marked with an asterisk (*) showed lower scores. g. Pyridine hemochrome assay of the supercomplex, reduced minus oxidized spectrum is shown. The peaks of the corresponding hemes are indicated in the figure. The estimated ratio of heme a, b and c is 1.1:0.9:1.3.
Supplementary Figure 2 Composition of the M. smegmatis supercomplex.
In addition to complexes III2 and IV2, four additional subunits are identified based on mass spectrometry. Open reading frame as well as number of predicted transmembrane helices (TMH) (Protter) are given.
Supplementary Figure 3 Cryo-EM map validation.
a. Example micrograph with typical particle images circled in red. b. Example 2-D class average images showing different views of the structure. c. Fourier shell correlation curve for estimation of overall map resolution. d. Estimation of local resolution.
Supplementary Figure 4 M. smegmatis complex III.
Comparison of M. smegmatis complex III quinone binding sites (blue) to those of complex III from S. cerevisiae (3CX5) (green) and chicken (3l71) (purple). a. Qo-site in relation to inhibitor bound structures, purple: 3l71 with azoxystrobin, green, 3cx5 with stigmatellin. MQ9 binds in a pocket formed partly by QcrC (yellow), which has no direct counterpart in the other complex III structures. Structural differences in the α-helix between the respective binding sites highlighted. b. Qi-site in relation to quinone binding in 3l71. Structural differences are highlighted.
Supplementary Figure 5 M. smegmatis cytochrome c oxidase.
a. Cardiolipin-mediated interactions between CtaE and CtaF, bound by two arginine residues, as well as CtaF and complex III. b. in the M. smegmatis SC. All cardiolipin headgroups face the N-side of the membrane.
Supplementary Figure 6 Detection of SodC associated with the III2IV2 supercomplex.
a. The M. smegmatis supercomplex and control protein (left panel) were resolved on CN-PAGE. The bleached band corresponding to the size of the MsSC (“SodC in SC”) indicates SodC activity, not observed in the control protein. A second bleached band (indicated by “free SodC”) running below 66 kDa presumably represents unbound SOD. The right panel shows the same gel after CBB staining to visualize the proteins. b. Inhibition curve for bovine Cu-Zn SOD (Sigma) and the M. smegmatis supercomplex. The concentration of the supercomplex is expressed in pmol of heme a. c. Detection of SodC activity in the M. smegmatis but not the C. glutamicum SC. Left panel- MsSC shows associated positive SodC activity, which is absent in CgSC. Right panel, MsSC, and CgSC protein profile on BN-PAGE.
Supplementary Figure 7 Additional subunits in M. smegmatis cytochrome c oxidase.
a. New subunits identified at complex IV include MSMEG_4692, MSMEG_4693 as well as three unidentified additional subunits. b. Superposition of MSMEG_4692 (blue) to the structure of a TPM-domain with ATPase/ADPase activity from Bizionia argentinensis (5ANP, pink). Despite no detectable sequence homology the domains are structurally very similar with an RMSD of 1.8 Å over 110 aligned residues. TMP-domains are a poorly characterized family shown to be capable of phosphate hydrolysis from various substrates, including nucleotides.
Supplementary Figure 8 Rescue of the M. smegmatis ∆qcrCAB strain by the qcrCAB operon in the plasmid and verification of expression.
a. Colonies of ∆qcrCAB strain harboring the construct PST-KTqcrCAB grow significantly larger than those of the control strain (PST-KT). The 7H9 agar plates are shown after 5 days of growth at 37 °C. b. 7H9 liquid culture. PST-KTqcrCAB bearing cells show comparable growth to wild-type M. smegmatis (bearing PST-KT) while control plasmid (PST-KT) bearing ∆qcrCAB cells show impaired growth. The OD600 after 48 hr of growth at 37°C (starting OD = 0.05) are shown. Error bars indicate SD (n=3). c. Verification of the expression of SC proteins. Detection of FLAG tagged cytochrome b in Ms∆qcrCAB+PST-KTqcrCAB strain by western blotting. A single band is detected at 57 kDa. Lanes 1 and 2 are loaded with lysates from Ms∆qcrCAB+PST-KTqcrCAB and WT+PST-KT cells, respectively. d. Detection of cytochrome cc by heme staining. Membranes purified from the ∆qcrCAB+PST-KTqcrCAB were resolved on SDS-PAGE and subjected to heme staining. The molecular weight of marker proteins is shown on the left.
Supplementary Figure 9 Cryo-EM image processing workflow.
a. Summary of image processing steps for final cryo-EM map generation. b. Orientation distribution plot for particle images contributing to the final cryo-EM map.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–9
Supplementary Video 1
M. smegmatis supercomplex dynamics. Dynamics of the cyt c domain of the M. smegmatis supercomplex
Rights and permissions
About this article

Cite this article
Wiseman, B., Nitharwal, R.G., Fedotovskaya, O. et al. Structure of a functional obligate complex III2IV2 respiratory supercomplex from Mycobacterium smegmatis. Nat Struct Mol Biol 25, 1128–1136 (2018). https://doi.org/10.1038/s41594-018-0160-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41594-018-0160-3
This article is cited by
-
Mycobacterial Regulatory Systems Involved in the Regulation of Gene Expression Under Respiration-Inhibitory Conditions
Journal of Microbiology (2023)
-
Structural basis for safe and efficient energy conversion in a respiratory supercomplex
Nature Communications (2022)
-
Targeting the cytochrome bc1 complex for drug development in M. tuberculosis: review
Molecular Diversity (2022)
-
Homology modeling and molecular docking simulation of some novel imidazo[1,2-a]pyridine-3-carboxamide (IPA) series as inhibitors of Mycobacterium tuberculosis
Journal of Genetic Engineering and Biotechnology (2021)
-
Structure and assembly of the mammalian mitochondrial supercomplex CIII2CIV
Nature (2021)
