
Abstract
Oxytocin (OT) orchestrates social and emotional behaviors through modulation of neural circuits. In the central amygdala, the release of OT modulates inhibitory circuits and, thereby, suppresses fear responses and decreases anxiety levels. Using astrocyte-specific gain and loss of function and pharmacological approaches, we demonstrate that a morphologically distinct subpopulation of astrocytes expresses OT receptors and mediates anxiolytic and positive reinforcement effects of OT in the central amygdala of mice and rats. The involvement of astrocytes in OT signaling challenges the long-held dogma that OT acts exclusively on neurons and highlights astrocytes as essential components for modulation of emotional states under normal and chronic pain conditions.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Data and code availability
Python code (used for ex vivo calcium imaging data analysis) can be found in Supplementary Software. All data that support the findings of this study are available from the corresponding authors upon reasonable request.
References
Lee, H.-J., Macbeth, A. H., Pagani, J. H. & Young, W. S. Oxytocin: the great facilitator of life. Prog. Neurobiol. 88, 127–151 (2009).
Knobloch, H. S. et al. Evoked axonal oxytocin release in the central amygdala attenuates fear response. Neuron 73, 553–566 (2012).
Chini, B., Verhage, M. & Grinevich, V. The action radius of oxytocin release in the mammalian CNS: from single vesicles to behavior. Trends Pharmacol. Sci. 38, 982–991 (2017).
Mitre, M. et al. A distributed network for social cognition enriched for oxytocin receptors. J. Neurosci. 36, 2517–2535 (2016).
Yoshida, M. et al. Evidence that oxytocin exerts anxiolytic effects via oxytocin receptor expressed in serotonergic neurons in mice. J. Neurosci. 29, 2259–2271 (2009).
Theodosis, D. T. Oxytocin-secreting neurons: a physiological model of morphological neuronal and glial plasticity in the adult hypothalamus. Front. Neuroendocrinol. 23, 101–135 (2002).
Pacholko, A. G., Wotton, C. A. & Bekar, L. K. Astrocytes—the ultimate effectors of long-range neuromodulatory networks? Front. Cell. Neurosci. 14, 1–12 (2020).
Huber, D., Veinante, P. & Stoop, R. Vasopressin and oxytocin excite distinct neuronal populations in the central amygdala. Science 308, 245–248 (2005).
Viviani, D. et al. Oxytocin selectively gates fear responses through distinct outputs from the central amygdala. Science 333, 104–107 (2011).
Han, R. T. et al. Long-term isolation elicits depression and anxiety-related behaviors by reducing oxytocin-induced GABAergic transmission in central amygdala. Front. Mol. Neurosci. 11, 246 (2018).
Neugebauer, V., Li, W., Bird, G. C. & Han, J. S. The amygdala and persistent pain. Neuroscientist 10, 221–234 (2004).
Tye, K. M. et al. Amygdala circuitry mediating reversible and bidirectional control of anxiety. Nature 471, 358–362 (2011).
Ji, R.-R., Donnelly, C. R. & Nedergaard, M. Astrocytes in chronic pain and itch. Nat. Rev. Neurosci. 20, 667–685 (2019).
Poisbeau, P., Grinevich, V. & Charlet, A. Oxytocin signaling in pain: cellular, circuit, system, and behavioral levels. Curr. Top. Behav. Neurosci. 35, 193–211 (2018).
Martin-Fernandez, M. et al. Synapse-specific astrocyte gating of amygdala-related behavior. Nat. Neurosci. 20, 1540–1548 (2017).
Althammer, F. et al. Three-dimensional morphometric analysis reveals time-dependent structural changes in microglia and astrocytes in the central amygdala and hypothalamic paraventricular nucleus of heart failure rats. J. Neuroinflammation 17, 221 (2020).
Yizhar, O. et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 477, 171–178 (2011).
Lee, H. J., Caldwell, H. K., Macbeth, A. H., Tolu, S. G. & Young, W. S. A conditional knockout mouse line of the oxytocin receptor. Endocrinology 149, 3256–3263 (2008).
Wallraff, A. et al. The impact of astrocytic gap junctional coupling on potassium buffering in the hippocampus. J. Neurosci. 26, 5438–5447 (2006).
Papouin, T., Dunphy, J. M., Tolman, M., Dineley, K. T. & Haydon, P. G. Septal cholinergic neuromodulation tunes the astrocyte-dependent gating of hippocampal NMDA receptors to wakefulness. Neuron 94, 840–854 (2017).
Robin, L. M. et al. Astroglial CB1 receptors determine synaptic D-serine availability to enable recognition memory. Neuron 98, 935–944 (2018).
Corkrum, M., Rothwell, P. E., Thomas, M. J., Kofuji, P. & Araque, A. Opioid-mediated astrocyte-neuron signaling in the nucleus accumbens. Cells 8, 586 (2019).
Li, Y. et al. Identity of endogenous NMDAR glycine site agonist in amygdala is determined by synaptic activity level. Nat. Commun. 4, 1760 (2013).
Decosterd, I. & Woolf, C. J. Spared nerve injury: an animal model of persistent peripheral neuropathic pain. Pain 87, 149–158 (2000).
Thompson, J. M. & Neugebauer, V. Amygdala plasticity and pain. Pain Res. Manag. 2017, 8296501 (2017).
Sieberg, C. B. et al. Neuropathic pain drives anxiety behavior in mice, results consistent with anxiety levels in diabetic neuropathy patients. Pain Rep. 3, e651 (2018).
László, K. et al. Positive reinforcing effect of oxytocin microinjection in the rat central nucleus of amygdala. Behav. Brain Res. 296, 279–285 (2016).
Goodin, B. R., Ness, T. J. & Robbins, M. T. Oxytocin - a multifunctional analgesic for chronic deep tissue pain. Curr. Pharm. Des. 21, 906–913 (2015).
Gravati, M. et al. Dual modulation of inward rectifier potassium currents in olfactory neuronal cells by promiscuous G protein coupling of the oxytocin receptor. J. Neurochem. 114, 1424–1435 (2010).
Breton, J.-D. et al. Oxytocin-induced antinociception in the spinal cord is mediated by a subpopulation of glutamatergic neurons in lamina I–II which amplify GABAergic inhibition. Mol. Pain 4, 19 (2008).
Angulo, M. C., Kozlov, A. S., Charpak, S. & Audinat, E. Glutamate released from glial cells synchronizes neuronal activity in the hippocampus. J. Neurosci. 24, 6920–6927 (2004).
Fellin, T. et al. Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors. Neuron 43, 729–743 (2004).
Corkrum, M. et al. Dopamine-evoked synaptic regulation in the nucleus accumbens requires astrocyte activity. Neuron 105, 1036–1047.e5 (2020).
Ma, Z., Stork, T., Bergles, D. E. & Freeman, M. R. Neuromodulators signal through astrocytes to alter neural circuit activity and behaviour. Nature 539, 428–432 (2016).
Pestana, F., Edwards-Faret, G., Belgard, T. G., Martirosyan, A. & Holt, M. G. No longer underappreciated: the emerging concept of astrocyte heterogeneity in neuroscience. Brain Sci. 10, 1–21 (2020).
Khakh, B. S. & Deneen, B. The emerging nature of astrocyte diversity. Annu. Rev. Neurosci. 42, 187–207 (2019).
Grinevich, V. & Neumann, I. D. Brain oxytocin: how puzzle stones from animal studies translate into psychiatry. Mol. Psychiatry 26, 265–279 (2021).
Quintana, D. S. et al. Advances in the field of intranasal oxytocin research: lessons learned and future directions for clinical research. Mol. Psychiatry 26, 80–91 (2021).
Kirsch, P. et al. Oxytocin modulates neural circuitry for social cognition and fear in humans. J. Neurosci. 25, 11489–11493 (2005).
Takata, N. et al. Optogenetic astrocyte activation evokes BOLD fMRI response with oxygen consumption without neuronal activity modulation. Glia 66, 2013–2023 (2018).
Figley, C. R. & Stroman, P. W. The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals. Eur. J. Neurosci. 33, 577–588 (2011).
Kastanenka, K. V. et al. A roadmap to integrate astrocytes into systems neuroscience. Glia 68, 5–26 (2019).
Poskanzer, K. E. & Yuste, R. Astrocytes regulate cortical state switching in vivo. Proc. Natl Acad. Sci. USA 2016, 1–10 (2016).
Kjaerby, C., Rasmussen, R., Andersen, M. & Nedergaard, M. Does global astrocytic calcium signaling participate in awake brain state transitions and neuronal circuit function? Neurochem. Res. 42, 1810–1822 (2017).
Fenoy, A. J., Goetz, L., Chabardès, S. & Xia, Y. Deep brain stimulation: are astrocytes a key driver behind the scene? CNS Neurosci. Ther. 20, 191–201 (2014).
Monai, H. & Hirase, H. Astrocytes as a target of transcranial direct current stimulation (tDCS) to treat depression. Neurosci. Res. 126, 15–21 (2018).
Mills, S. E. E., Nicolson, K. P. & Smith, B. H. Chronic pain: a review of its epidemiology and associated factors in population-based studies. Br. J. Anaesth. 123, e273–e283 (2019).
Woo, A. K. Depression and anxiety in pain. Rev. Pain 4, 8–12 (2010).
Oliveira, J. F., Sardinha, V. M., Guerra-Gomes, S., Araque, A. & Sousa, N. Do stars govern our actions? Astrocyte involvement in rodent behavior. Trends Neurosci. 38, 535–549 (2015).
Rouach, N., Koulakoff, A., Abudara, V., Willecke, K. & Giaume, C. Astroglial metabolic networks sustain hippocampal synaptic transmission. Science 322, 1551–1555 (2008).
Pannasch, U. et al. Astroglial networks scale synaptic activity and plasticity. Proc. Natl Acad. Sci. USA 108, 8467–8472 (2011).
Theis, M. et al. Accelerated hippocampal spreading depression and enhanced locomotory activity in mice with astrocyte-directed inactivation of connexin43. J. Neurosci. 23, 766–776 (2003).
Teubner, B. et al. Connexin30 (Gjb6)-deficiency causes severe hearing impairment and lack of endocochlear potential. Hum. Mol. Genet. 12, 13–21 (2003).
Shigetomi, E. et al. Imaging calcium microdomains within entire astrocyte territories and endfeet with GCaMPs expressed using adeno-associated viruses. J. Gen. Physiol. 141, 633–647 (2013).
Ting, J. T. et al. Preparation of acute brain slices using an optimized N-methyl-D-glucamine protective recovery method. J. Vis. Exp. (132), 53825 (2018).
Serrano, A., Haddjeri, N., Lacaille, J., Robitaille, R. & Centre-ville, S. GABAergic network activation of glial cells underlies hippocampal heterosynaptic depression. J. Neurosci. 26, 5370–5382 (2006).
Jourdain, P. et al. Glutamate exocytosis from astrocytes controls synaptic strength. Nat. Neurosci. 10, 331–339 (2007).
Ikegaya, Y., Le Bon-Jego, M. & Yuste, R. Large-scale imaging of cortical network activity with calcium indicators. Neurosci. Res. 52, 132–138 (2005).
Shigetomi, E., Bowser, D. N., Sofroniew, M. V. & Khakh, B. S. Two forms of astrocyte calcium excitability have distinct effects on NMDA receptor-mediated slow inward currents in pyramidal neurons. J. Neurosci. 28, 6659–6663 (2008).
Anlauf, E. & Derouiche, A. Glutamine synthetase as an astrocytic marker: its cell type and vesicle localization. Front. Endocrinol. 4, 144 (2013).
Luis-Delgado, O. E. et al. Calibrated forceps: a sensitive and reliable tool for pain and analgesia studies. J. Pain 7, 32–39 (2006).
Walf, A. A. & Frye, C. A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2, 322–328 (2007).
King, T. et al. Unmasking the tonic-aversive state in neuropathic pain. Nat. Neurosci. 12, 1364–1366 (2009).
Virtanen, P. et al. SciPy 1.0: fundamental algorithms for scientific computing in Python. Nat. Methods 17, 261–272 (2020).
Zeisel, A. et al. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 347, 1138–1142 (2015).
Batiuk, M. Y. et al. Identification of region-specific astrocyte subtypes at single cell resolution. Nat. Commun. 11, 1220 (2020).
Kofuji, P. & Araque, A. G-protein-coupled receptors in astrocyte–neuron communication. Neuroscience https://doi.org/10.1016/j.neuroscience.2020.03.025 (2020).
Octeau, J. C. et al. Transient, consequential increases in extracellular potassium ions accompany channelrhodopsin2 excitation. Cell Rep. 27, 2249–2261 (2019).
Reeves, A. M. B., Shigetomi, E. & Khakh, B. S. Bulk loading of calcium indicator dyes to study astrocyte physiology: key limitations and improvements using morphological maps. J. Neurosci. 31, 9353–9358 (2011).
Acknowledgements
This work was supported by the Centre National de la Recherche Scientifique contract UPR3212, the Université de Strasbourg contract UPR3212, IASP Early Career Research grant 2012, FP7 Career Integration grant 334455, Initiative of Excellence (IDEX) Attractiveness grant 2013, IDEX Interdisciplinary grant 2015, University of Strasbourg Institute for Advanced Study (USIAS) fellowship 2014-15, Foundation Fyssen research grant 2015, NARSAD Young Investigator grant 24821, ANR JCJC grant 19-CE16-0011-0 (to A.C.), ANR-DFG grant GR 3619/701 (to A.C. and V.G.), Alexander von Humboldt fellowship (to D.H.), Seed grant from DFG within the Collaborative Center SFB 1158 and Fyssen Foundation fellowship (to A.L.); DFG grants GR 3619/13-1, GR 3619/15-1 and GR 3619/16-1 (to V.G.); SFB 1158 (to C.P., J.S. and V.G.), SNSF-DFG grant GR 3619/8-1 (to R.S. and V.G.), the Fritz Thyssen Foundation (to V.G.), DFG Postdoctoral Fellowship AL 2466/1-1 (to F.A.), the Alexander von Humboldt Foundation (to D.H.), the Fyssen Foundation and PROCOP grant and SFB1158 seed grant for young scientists (to A.L.), post-doctoral fellowship from the Fund for Scientific Research Flanders (12V7519N) (to J.W.), Russian Science Foundation RSF (17-75-10061) and the subsidy allocated to the Kazan Federal University for the State Assignment, no. 0671-2020-0059 (to A.R.), the intramural research program of the NIMH (ZIAMH002498) (to W.S.Y.), National Institutes of Health grants R01NS094640 and R01HL090948, SFB/TRR 152 and ERC-CoG-772395 (to J.E.S.), the European Research Council (Consolidator Grant no. 683154) and the European Union’s Horizon 2020 Research and Innovation Program (Marie Sklodowska-Curie Innovative Training Networks, grant no. 722053, EU-GliaPhD) (to N.R.). The authors thank V. Lelièvre for in situ hybridization advice; R. Goutagny for in vivo optogenetics assistance; F. Magara for anxiety behavior advice; B. Kurpiers and the Interdisciplinary Neurobehavioral Core Facility of Heidelberg University for experiments performed there; S. Reibel and the Chronobiotron UMS 3415 for all animal care; and T. Splettstoesser (www.scistyle.com) for help with the preparation of Fig. 7.
Author information
Authors and Affiliations
Contributions
Conceptualization: A.C.; Methodology: A.C., B.Boutrel, C.M.L., C.P., D.K., F.A., I.D., J.E.S., J.W., J.Y.C., N.R., P.D., P.P., R.S., V.G., W.S.Y. and YG; Analysis: A.C., B.Bellanger, B.B.J., C.M.L., D.K., F.A., H.S.K.B., J.W. and S.G.; In situ hybridization: D.H., F.A., H.S.K.B., H.W., J.S. and M.E.; Immunohistochemistry: A.L., D.H., F.A., J.W., M.E. and M.D.S.G.; Imaris analysis: F.A., M.K.K. and R.K.R.; Ex vivo patch-clamp electrophysiology: A.B., A.C., J.W., S.G., D.K., I.W., B.Bellanger and M.A.; Ex vivo calcium imaging: A.B., C.M.L., D.K. and J.W.; Astrocyte characterization: A.B., A.R., B.Bellanger, D.K., I.W., M.E. and S.G.; Behavior: A.C., B.B.J., D.K. and J.W.; Mice line validation: W.S.Y.; Viral vectors validation: D.B., L.B., M.D.S.G., M.E. and V.G.; Spared nerve injuries: P.I. and M.P.; Writing: A.B., A.C., D.K., F.A., J.W. and V.G.; Funding acquisition: A.C. and V.G.; Supervision: A.C. and V.G.; Project administration: A.C.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Neuroscience thanks Thomas Papouin, Gertrudis Perea, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Validation of OTR expression in CeL astrocytes using several astrocytic markers.
a, Combination of FISH and IHC showing co-localization of GS (top left), NeuN (bottom left) and both markers (right) with OTR mRNA in rats. Scale bars are 100µm and 10µm. (n rats = 5, n slices = 20, n astrocytes = 1354, n neurons = 1254). b, proportion of OTR positive astrocytes and neurons in mice. n astrocytes = 897; n neurons = 688; n rats = 4 c, (top left) FISH overview for OTR mRNA (green), counterstained with polyclonal anti-ALDH1L1 antibody (red) in CeA. (bottom left) High magnification image of cells positive for both OTR mRNA and ALDH1L1 (double arrows); green arrows point OTR mRNA-positive cells; red arrows point ALDH1L1-positive cells. Scale bars: 400 (Top) and 50µm (Bottom). (Right) Quantification of ALDH1L1-positive cells positive for OTR mRNA. n astrocytes = 450; n rats = 4. d, RNAscope in situ hybridization showing GFAP (red) and OTR (green) expressing cells in mice CeA. Merged images include DAPI stain (blue); n = 3. (Bottom) Negative control probe targeting the bacterial gene DapB; n = 3. Scale bar is 10μm. st: stria terminalis. Data are expressed as mean across slices plus SEM and white circles represent individual cell data. (Statistics and numbers in Supplementary Table 1).
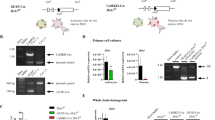
Extended Data Fig. 2 Validation of astrocyte-specific approaches.
a, (left) C1V1-mCherry (OxyOpto) expressing oxytocin neuron of the PVN. Scale bar is 20µm; n = 11. (right) Schematic representation of the whole cell patch clamp of the OxytOpto-expressing cells of the PVN. b, (left) λ542nm light exposure (yellow line) induce a depolarizing current, (right) enabling precise spiking of OxytOpto-expressing cells. c, Typical confocal image of CeL astrocytes co-labeled with SR101 and OGB1; n = 46. Scale bar 20μm. d, (left) Pseudo-color pictures of an SR101 positive cell identified as an astrocyte compared to neurons identified under oblique infrared light, Scale bar 10μm. (Middle) Electrophysiological properties of patched SR101+ (red, n = 82) and SR101- cells (grey, n = 20). Medians, quartiles and whiskers can be found in Supplementary Table 2. (Right) Typical responses to 20pA current steps of a SR101+ (red) and a SR101- cells (black). e, CeL SR101 positive cell filled with biocytin through whole cell patch-clamp (green) lacks NeuN signal (Blue); n = 3. Scale bar 50 μm. f, Illustration of calcium imaging data analysis method. (left) Fluorescence traces were splited into a “baseline” and a “post-stimulation” trace. Calcium transients were detected according to a prominence threshold and their frequency was quantified before and after the drug application. All data were averaged across astrocytes per slice, and this results was used as statistical unit. All data were compared (before vs. after drug application) and the results were expressed in ratio (baseline/drug effect), a ratio of 1 meaning neither an increase nor a decrease of the measured parameter. ns = 7, na = 36. g, (top left) Immunohistochemical staining for glutamine synthase (GS; blue), Cre-GFP (green) and Cre recombinase (red). (bottom left) Overview of the CeA, displaying correct viral targeting of the CeL subdivision. (right) Quantification of efficiency and specificity of the transduction. Efficiency: Over 1001 GS positive cells, 561 were also GFP positive, indicating an efficiency of 56±4.9%. Specificity: we counted a total of n = 977 GFP-positive cells, 940 of which were positive for GS (96.2±2.1%). None of the GFP or Cre signals were detected in NeuN positive cells (0 out of n = 850, 4 mice, not shown). Finally, 99.82 ± 0.2% of GS-positive astrocytes containing GFP signal were Cre-positive (n = 1001). n mice = 4. Scale bars: 400 (Bottom) and 20µm (Top). h, Immunohistochemical analysis of OTR expression in NeuN-positive cells of the CeL revealed no difference in OTR levels between control and GFAP-Cre injected animals; 85.0 ± 1.6% (n = 688, n = 4 mice) and 86.7 ± 1.6% (n = 660, n = 4 mice) respectively. Scale bars: 20µm. Data in f-h are expressed as mean across slices plus SEM and white circles represent individual cell data. ***p < 0.001, two-sided Mann-Whitney U test. (Statistics and numbers in Supplementary Table 2).
Extended Data Fig. 3 Astro-astrocytic communication in the CeL do not depends on purinergic nor NMDAR-dependent signalling.
a, Illustration of the 3D reconstruction and the quantification of the number of contact between astrocytes. (left) Pictures of the raw fluorescence obtained after anti-GFAP immunohistochemistry, (middle) based on this staining, we performed a 3D reconstruction of astrocytes and (right) evaluate the number of astrocyte-astrocyte contact using the given formula. With n = upper limit of summation, i = index of summation, xi = typical element (that is astrocytic contacts with 1µm or less proximity). With this approach, we quantified all astrocyte contacts without discrimination between individual astrocyte entities. b, (left) Typical ΔF traces following TGOT+TTX application in presence of PPADS (50 µM) + Suramin (75 µM) + A438079 (1 µM). (right) Proportion of responding astrocytes, AUC of ΔF traces and Ca2+ transients frequency normalized to baseline values following application of TGOT+TTX (0.4 μM) in presence of PPADS (50 µM) + Suramin (75 µM) + A438079 (1 µM) (ns = 10, na = 112). c, (left) Typical ΔF traces following TGOT+TTX application in presence of CPT (5 µM) or SCH 58261 (100 nM). (right) Proportion of responding astrocytes, AUC of ΔF traces and Ca2+ transients frequency normalized to baseline values following application of TGOT+TTX (0.4 μM) in presence of CPT (ns = 7, na = 22) or SCH (ns = 8, na = 32). d, (left) Typical ΔF traces following TGOT+TTX application in presence of AP5 (50 µM). (right) Proportion of responding astrocytes, AUC of ΔF traces and Ca2+ transients frequency normalized to baseline values following application of TGOT+TTX (0.4 μM) in presence of AP5 (ns = 6, na = 55). Data are expressed as mean across slices plus SEM and white circles represent individual cell data. White circles indicate average across astrocytes per slice. (Statistics and numbers in Supplementary Table 3).
Extended Data Fig. 4 Specific optogenetic stimulation of CeL astrocytes elicit calcium transients in a Ca2+-dependent manner.
a, (left) Immunohistochemistry image shows CeL cells transfected with rAAV-Gfap-C1V1(t/t)-mCherry (AstrOpto) with co-labeling for ALDH1L1. White arrow shows one cell expanded in insets. Scale bars are 25 and 10 µm (insets). (right) Quantification of the efficiency and specificity of transduction of C1V1 in CeL astrocytes. PGFAP- C1V1-mCherry rAAV vector was injected into rat CeA (bilaterally, 200nl). Specificity: Over 1090 mCherry-positive cells, 98.8±0.7% were positive for ALDH1L1. None of the analyzed cells were positive for NeuN. Efficiency: Over 1090 ALDH1L1 positive cells, 62.3±3.5% were also mCherry positive. n = 4 rats. b, (left) Typical ΔF traces following AstrOpto activation. λ542nm (1s @ 0.5Hz during 3 minutes) with (top 3 traces) or without (bottom 3 traces) extracellular calcium. (right) Proportion of responding astrocytes, AUC of ΔF traces and Ca2+ transients frequency normalized to baseline values following AstrOpto stimulation with (ns = 12, na = 49) or without Ca2+ in the ACSF (ns = 12, na = 53). Data are expressed as mean across slices plus SEM. White circles indicate average across astrocytes per slice. **p<0.01, ***p<0.001, two-sided Mann-Whitney U test. (Statistics and numbers in Supplementary Table 4).
Extended Data Fig. 5 Astro-neuronal communication depends on NMDAR but not purinergic transmission.
Figure 5. a, DAAO (0.15 IU/ml, incubation time > 1h30; n = 7) and AP5 (50 µM; n = 6) prevent the effect of photoactivation of astrocytes (AstrOpto) on IPSC frequency in CeM neurons. b, Two consecutive TGOT application effects on IPSCs frequencies in CeM neurons after DAAO (0.15 U/ml, incubation time > 1h30) followed by D-Serine (20 min, 100 μM) incubation of the same cells (n = 7). c, dOVT (1 µM; n = 6) prevents the effect of TGOT on IPSCs frequencies in CeM neurons. d, Effect of double (20 min apart) application of TGOT on IPSCs frequencies in CeM (0.4 µM n = 7). e, Effect of DCKA (10 µM, n = 15) and DNQX (25 µM; n = 10) on TGOT-induced increase in IPSC frequency in CeM neurons. f, Purinergic antagonists do not prevent TGOT effect on IPSC frequency in CeM neurons. PPADS (50 µM; n = 9), CPT (5 µM; n = 11), SCH (100 nM; n = 10). Data are expressed as averages plus SEM and white circles represent individual cell data. #p < 0.05, ##p < 0.01, ###p < 0.001, Friedman and Dunn’s Multiple comparisons, **p < 0.01, ***p < 0.001, two-sided unpaired t-test or Mann-Whitney U test. (Statistics and numbers in Supplementary Table 5).
Extended Data Fig. 6 SNI procedure does not modify the effect of OTR activation on CeL microcircuit activity.
a, 30 days post surgeries time course of mechanical pain threshold evolution across sham (n = 23) and SNI (n = 22) rats. Data are expressed as mean across animals plus SEM. b, (left) Typical ΔF traces following TGOT+TTX application in SNI rats. (right) Proportion of responding astrocytes, AUC of ΔF traces and Ca2+ transients frequency normalized to baseline values following application of TGOT+TTX (0.4 μM) in sham (ns = 16, na = 74) or SNI (ns = 18 et na = 136) rats. Data are expressed as mean across slices plus SEM. White circles indicate average across astrocytes per slice. c, TGOT effect on CeM neurons IPSCs frequencies is unchanged between Sham (n = 9) or SNI (n = 9) rats. Data are expressed as average plus SEM and white circles represent individual cell data. d, Two consecutive TGOT application effects on IPSCs frequencies in CeM neurons after DAAO (0.15 U/ml, incubation time > 1h30) followed by D-Serine (20 min, 100 μM) incubation of the same cells from acute brain slices of SNI animals (n = 11). Data are expressed as averages plus SEM and white circles represent individual cell data. e, Verification of cannulae implantation sites. (left) Schematic representation of the stereotaxic injection viewed in a coronal slice; n rats = 67, n mice = 52. (middle, right) Infrared and fluorescence microscopy picture showing the injection site of fluorescent latex beads in the CeL on horizontal slices. Scale is 1mm. ** p<0.01, *** P<0.001, two-sided paired t-test or Wilcoxon test, #p<0.05 Friedman test followed by Dunn’s multiple comparison. (Statistics and numbers in Supplementary Table 6).
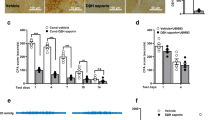
Extended Data Fig. 7 Effects of CeL astrocytes and OTR activities manipulations on locomotion and contralateral hindpaw sensitivity.
a,e, Experimental strategy for the specific expression of C1V1 in mice CeL astrocytes (a, AstrOpto) or the specific deletion of OTRs in mice CeL astrocytes (e, GFAP OTR KO). The treatments applied are color coded as the legend key indicate. Control indicate a vehicle injection. b,f, Mechanical pain threshold was assessed on the non-injured paw of SNI (bottom, gray box) and its equivalent in Sham (top) groups. TGOT or its vehicle, or astrocytes light-evoked activation of C1V1, were administered in the CeL and mechanical pain threshold assessed again at different time points. Rats (n): Sham control n = 12, TGOT n = 10, C1V1 n = 6; SNI control n = 12, TGOT n = 10, C1V1, n = 9. Mice (n): Sham WT n = 6, OTR cKO n = 8; SNI WT n = 5, OTR cKO n = 13. c,g, Locomotion was assessed through measurement of the distance travelled during the length of the elevated plus maze experiment, after administration of the different treatments. Rats (n): Sham control n = 10, TGOT n = 8, C1V1 n = 9; SNI control n = 20, TGOT n = 9, C1V1, n = 7. Mice (n): Sham WT Veh n = 7, WT TGOT n = 6; Sham OTR cKO Veh n = 5, OTR cKO TGOT n = 7; SNI WT Veh n = 7, WT TGOT n = 8; SNI OTR cKO Veh n = 5, OTR cKO TGOT n = 7. d,h, Locomotion was assessed through measurement of the distance travelled during the time of the conditioned place preference experiment, after administration of the different treatments. Rats (n): Sham control n = 13, TGOT n = 6, C1V1 n = 5; SNI control n = 10, TGOT n = 8, C1V1, n = 8. Mice (n): Sham WT Veh n = 5, WT TGOT n = 8; Sham OTR cKO Veh n = 5, OTR cKO TGOT n = 4; SNI WT Veh n = 5, WT TGOT n = 5; SNI OTR cKO Veh n = 4, OTR cKO TGOT n = 6. Data are expressed as mean across animals plus SEM. (Animals number and Statistics in Supplementary Table 7).
Supplementary information
Supplementary Tables
Tables 1–7: Numerical values and statistical analysis of data presented in Figs.1–7 and Extended Data Figs. 1–7. Table 8: List of reagents used in this study
Supplementary Software
Calcium imaging analysis using Python code
Rights and permissions
About this article

Cite this article
Wahis, J., Baudon, A., Althammer, F. et al. Astrocytes mediate the effect of oxytocin in the central amygdala on neuronal activity and affective states in rodents. Nat Neurosci 24, 529–541 (2021). https://doi.org/10.1038/s41593-021-00800-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-021-00800-0
This article is cited by
-
A conceptual framework for astrocyte function
Nature Neuroscience (2023)
-
Detection, processing and reinforcement of social cues: regulation by the oxytocin system
Nature Reviews Neuroscience (2023)
-
Astrocytes in Chronic Pain: Cellular and Molecular Mechanisms
Neuroscience Bulletin (2023)
-
Oxytocin Receptor Expression in Hair Follicle Stem Cells: A Promising Model for Biological and Therapeutic Discovery in Neuropsychiatric Disorders
Stem Cell Reviews and Reports (2023)
-
An analgesic pathway from parvocellular oxytocin neurons to the periaqueductal gray in rats
Nature Communications (2023)
