
Abstract
Tissue repair after spinal cord injury requires the mobilization of immune and glial cells to form a protective barrier that seals the wound and facilitates debris clearing, inflammatory containment and matrix compaction. This process involves corralling, wherein phagocytic immune cells become confined to the necrotic core, which is surrounded by an astrocytic border. Here we elucidate a temporally distinct gene signature in injury-activated microglia and macrophages (IAMs) that engages axon guidance pathways. Plexin-B2 is upregulated in IAMs and is required for motor sensory recovery after spinal cord injury. Plexin-B2 deletion in myeloid cells impairs corralling, leading to diffuse tissue damage, inflammatory spillover and hampered axon regeneration. Corralling begins early and requires Plexin-B2 in both microglia and macrophages. Mechanistically, Plexin-B2 promotes microglia motility, steers IAMs away from colliding cells and facilitates matrix compaction. Our data therefore establish Plexin-B2 as an important link that integrates biochemical cues and physical interactions of IAMs with the injury microenvironment during wound healing.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
Data availability
The data that support the findings of this study are available from the corresponding authors upon request. RNA-seq data files were deposited at the National Center for Biotechnology Information (NCBI) GEO repository under accession number GSE113566.
References
Silver, J. & Miller, J. H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 5, 146–156 (2004).
Burda, J. E. & Sofroniew, M. V. Reactive gliosis and the multicellular response to CNS damage and disease. Neuron 81, 229–248 (2014).
Courtine, G. & Sofroniew, M. V. Spinal cord repair: advances in biology and technology. Nat. Med. 25, 898–908 (2019).
Davalos, D. et al. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 8, 752–758 (2005).
Beck, K. D. et al. Quantitative analysis of cellular inflammation after traumatic spinal cord injury: evidence for a multiphasic inflammatory response in the acute to chronic environment. Brain 133, 433–447 (2010).
Popovich, P. G. & Jones, T. B. Manipulating neuroinflammatory reactions in the injured spinal cord: back to basics. Trends Pharmacol. Sci. 24, 13–17 (2003).
Hines, D. J., Hines, R. M., Mulligan, S. J. & Macvicar, B. A. Microglia processes block the spread of damage in the brain and require functional chloride channels. Glia 57, 1610–1618 (2009).
Carlson, S. L., Parrish, M. E., Springer, J. E., Doty, K. & Dossett, L. Acute inflammatory response in spinal cord following impact injury. Exp. Neurol. 151, 77–88 (1998).
Friedel, R. H. et al. Plexin-B2 controls the development of cerebellar granule cells. J. Neurosci. 27, 3921–3932 (2007).
Daviaud, N., Chen, K., Huang, Y., Friedel, R. H. & Zou, H. Impaired cortical neurogenesis in plexin-B1 and -B2 double deletion mutant. Dev. Neurobiol. 76, 882–899 (2016).
Nakada, M. et al. Molecular targets of glioma invasion. Cell. Mol. Life Sci. 64, 458–478 (2007).
Gurrapu, S. & Tamagnone, L. Transmembrane semaphorins: multimodal signaling cues in development and cancer. Cell Adh. Migr. 10, 675–691 (2016).
Koropouli, E. & Kolodkin, A. L. Semaphorins and the dynamic regulation of synapse assembly, refinement, and function. Curr. Opin. Neurobiol. 27, 1–7 (2014).
Kumanogoh, A. (ed.) Semaphorins—A Diversity of Emerging Physiological and Pathological Activities (Springer Japan, 2015).
Kumanogoh, A. & Kikutani, H. Immunological functions of the neuropilins and plexins as receptors for semaphorins. Nat. Rev. Immunol. 13, 802–814 (2013).
Negishi-Koga, T. & Takayanagi, H. Bone cell communication factors and semaphorins. Bonekey Rep. 1, 183 (2012).
Pasterkamp, R. J. Getting neural circuits into shape with semaphorins. Nat. Rev. Neurosci. 13, 605–618 (2012).
Sakurai, A., Doçi, C. L., Doci, C. & Gutkind, J. S. Semaphorin signaling in angiogenesis, lymphangiogenesis and cancer. Cell Res. 22, 23–32 (2012).
Tran, T. S., Kolodkin, A. L. & Bharadwaj, R. Semaphorin regulation of cellular morphology. Annu. Rev. Cell Dev. Biol. 23, 263–292 (2007).
Worzfeld, T. & Offermanns, S. Semaphorins and plexins as therapeutic targets. Nat. Rev. Drug Discov. 13, 603–621 (2014).
Xia, J. et al. Semaphorin–plexin signaling controls mitotic spindle orientation during epithelial morphogenesis and repair. Dev. Cell 33, 299–313 (2015).
Mo, A. et al. Epigenomic signatures of neuronal diversity in the mammalian brain. Neuron 86, 1369–1384 (2015).
Parkhurst, C. N. et al. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell 155, 1596–1609 (2013).
Kuboyama, T. et al. HDAC3 inhibition ameliorates spinal cord injury by immunomodulation. Sci. Rep. 7, 8641 (2017).
Parikh, P. et al. Regeneration of axons in injured spinal cord by activation of bone morphogenetic protein/Smad1 signaling pathway in adult neurons. Proc. Natl Acad. Sci. USA 108, E99–E107 (2011).
Maier, V. et al. Semaphorin 4C and 4G are ligands of Plexin-B2 required in cerebellar development. Mol. Cell. Neurosci. 46, 419–431 (2011).
Kettenmann, H., Hanisch, U. K., Noda, M. & Verkhratsky, A. Physiology of microglia. Physiol. Rev. 91, 461–553 (2011).
Basso, D. M. et al. Basso mouse scale for locomotion detects differences in recovery after spinal cord injury in five common mouse strains. J. Neurotrauma 23, 635–659 (2006).
Bradbury, E. J. et al. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 416, 636–640 (2002).
Hickman, S. E. et al. The microglial sensome revealed by direct RNA sequencing. Nat. Neurosci. 16, 1896–1905 (2013).
Bays, J. L. & DeMali, K. A. Vinculin in cell–cell and cell–matrix adhesions. Cell. Mol. Life Sci. 74, 2999–3009 (2017).
Keren-Shaul, H. et al. A unique microglia type associated with restricting development of Alzheimer’s disease. Cell 169, 1276–1290.e17 (2017).
Liddelow, S. A. et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 541, 481–487 (2017).
van Furth, R. Origin and turnover of monocytes and macrophages. Curr. Top. Pathol. 79, 125–150 (1989).
Fogg, D. K. et al. A clonogenic bone marrow progenitor specific for macrophages and dendritic cells. Science 311, 83–87 (2006).
Robinson, S. P., Langan-Fahey, S. M., Johnson, D. A. & Jordan, V. C. Metabolites, pharmacodynamics, and pharmacokinetics of tamoxifen in rats and mice compared to the breast cancer patient. Drug Metab. Dispos. 19, 36–43 (1991).
Deng, S. et al. Plexin-B2, but not Plexin-B1, critically modulates neuronal migration and patterning of the developing nervous system in vivo. J. Neurosci. 27, 6333–6347 (2007).
Hirschberg, A. et al. Gene deletion mutants reveal a role for semaphorin receptors of the plexin-B family in mechanisms underlying corticogenesis. Mol. Cell. Biol. 30, 764–780 (2010).
Saha, B., Ypsilanti, A. R., Boutin, C., Cremer, H. & Chedotal, A. Plexin-B2 regulates the proliferation and migration of neuroblasts in the postnatal and adult subventricular zone. J. Neurosci. 32, 16892–16905 (2012).
Deb Roy, A. et al. Optogenetic activation of Plexin-B1 reveals contact repulsion between osteoclasts and osteoblasts. Nat. Commun. 8, 15831 (2017).
Wanner, I. B. et al. Glial scar borders are formed by newly proliferated, elongated astrocytes that interact to corral inflammatory and fibrotic cells via STAT3-dependent mechanisms after spinal cord injury. J. Neurosci. 33, 12870–12886 (2013).
Anderson, M. A. et al. Astrocyte scar formation aids central nervous system axon regeneration. Nature 532, 195–200 (2016).
Kaur, C., Rathnasamy, G. & Ling, E. A. Biology of microglia in the developing brain. J. Neuropathol. Exp. Neurol. 76, 736–753 (2017).
Zhao, X. & Guan, J. L. Focal adhesion kinase and its signaling pathways in cell migration and angiogenesis. Adv. Drug Deliv. Rev. 63, 610–615 (2011).
Meltzer, S. et al. Epidermis-derived semaphorin promotes dendrite self-avoidance by regulating dendrite-substrate adhesion in Drosophila sensory neurons. Neuron 89, 741–755 (2016).
Zeisel, A. et al. Brain structure. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 347, 1138–1142 (2015).
Zhang, Y. et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 34, 11929–11947 (2014).
Bellver-Landete, V. et al. Microglia are an essential component of the neuroprotective scar that forms after spinal cord injury. Nat. Commun. 10, 518 (2019).
Brennan, F. H., Hall, J. C. E., Guan, Z. & Popovich, P. G. Microglia limit lesion expansion and promote functional recovery after spinal cord injury in mice. Preprint at bioRxiv https://www.biorxiv.org/content/10.1101/410258v1 (2018).
Busch, S. A., Horn, K. P., Silver, D. J. & Silver, J. Overcoming macrophage-mediated axonal dieback following CNS injury. J. Neurosci. 29, 9967–9976 (2009).
Descalzi, G. et al. Neuropathic pain promotes adaptive changes in gene expression in brain networks involved in stress and depression. Sci. Signal. 10, eaaj1549 (2017).
Mitsi, V. et al. RGS9-2—controlled adaptations in the striatum determine the onset of action and efficacy of antidepressants in neuropathic pain states. Proc. Natl Acad. Sci. USA 112, E5088–E5097 (2015).
Andrews, S. FastQC: a quality control tool for high throughput sequence data. Babraham Bioinformatics https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (2010).
Bonfert, T., Kirner, E., Csaba, G., Zimmer, R. & Friedel, C. C. ContextMap 2: fast and accurate context-based RNA-seq mapping. BMC Bioinf. 16, 122 (2015).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. Roy. Stat. Soc. B 57, 289–300 (1995).
Chen, E. Y. et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics 14, 128 (2013).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–W97 (2016).
Jensen, E. C. Quantitative analysis of histological staining and fluorescence using ImageJ. Anat. Rec. 296, 378–381 (2013).
Hintze, J. L. & Nelson, R. D. Violin plots: a box plot-density trace synergism. Am. Stat. 52, 181–184 (1998).
Longair, M. H., Baker, D. A. & Armstrong, J. D. Simple neurite tracer: open source software for reconstruction, visualization and analysis of neuronal processes. Bioinformatics 27, 2453–2454 (2011).
Morrison, H. W. & Filosa, J. A. A quantitative spatiotemporal analysis of microglia morphology during ischemic stroke and reperfusion. J. Neuroinflammation 10, 782 (2013).
Meijering, E., Dzyubachyk, O. & Smal, I. Methods for cell and particle tracking. Methods Enzymol. 504, 183–200 (2012).
Scarpa, E. et al. A novel method to study contact inhibition of locomotion using micropatterned substrates. Biol. Open 2, 901–906 (2013).
Finelli, M. J., Wong, J. K. & Zou, H. Epigenetic regulation of sensory axon regeneration after spinal cord injury. J. Neurosci. 33, 19664–19676 (2013).
Acknowledgements
We thank all members of the Zou and Friedel laboratories for constructive comments. The work was supported by grants from the following organizations: the NIH/NINDS (R01/R56 NS073596), the Craig H. Neilsen Foundation (#476516) and the New York State Spinal Cord Injury Research Board (DOH01-C32242GG, DOH01-C33268GG) to H.Z.; the NIH/NINDS (R01 NS092735) to R.H.F.; the NIH/NINDS (NS086444, NS093537) and the NIH/NIA (U01 AG046170) to B.Z.; and the National Institute on Drug Abuse (P01 DA08227) to V.Z. Additional fellowship support was provided by the Chinese Scholarship Council to X.Z. and the New York State Spinal Cord Injury Research Board (DOH01-C32634GG) to S.W. M.-S.F., M.K. and C.C.F. were supported by grants FR2938/7-1 and CRC 1123 (Z2) from the Deutsche Forschungsgemeinschaft (DFG) to C.C.F. We would also like to thank Z. Yue and Y. Zhang for assisting in the rotarod tests, and Mount Sinai Microscopy CoRE for assisting in the STED imaging.
Author information
Authors and Affiliations
Contributions
X.Z. and S.W. conducted experiments and collected data. X.H., R.H.F. and H.Z. designed the study. K.A. and V.Z. provided expertise for the von Frey filament sensory assays, and M.-S.F., M.K., C.C.F., L.G. and B.Z. performed the bioinformatics analyses. All authors participated in data analyses and contributed to the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Neuroscience thanks Jerry Silver, Pate Skene, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
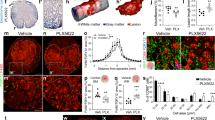
Extended Data Fig. 1 Upregulation of Plexin-B2 in activated microglia/macrophages after SCI.
a, X-Gal staining of sagittal section of thoracic spinal cords from WT mouse show negative lacZ activity in sham or after SCI at 7 dpi. Images from 3 independent mice for each condition showed similar results. b, IHC images show Plexin-B2 induction at the injury site at 7 dpi after SCI, which co-localizes with CD11b. Images from 4 independent mice for each condition showed similar results. c, IHC images of thoracic spinal cords from Cx3cr1GFP reporter mice show co-localization of Plexin-B2 and GFP at three time points after T8 transection. Images from 4 independent mice for each condition showed similar results. d, IHC images show minimal overlap between Plexin-B2 and the indicated neural markers at the injury site after SCI. Quantifications demonstrate that Plexin-B2 upregulation largely occurs in IBA1+ microglia/macrophages at three time points after SCI. Images from 4 independent mice showed similar results. e, Immunostaining of primary microglia (IBA1+) isolated from Cx3cr1CreER Plxnb2fl/B mice with no Tamoxifen exposure show Plexin-B2 induction upon LPS stimulation. Quantifications are shown on the right. n=100 cells counted from 3 independent cultures for each condition, unpaired two-tailed Student’s t test, ***p<0.0001. Data represent mean ± SEM. f, IHC images with long exposure show baseline expression of PB2 that colocalizes with IBA1 in uninjured spinal cord in control mice, but ablation of Plexin-B2 in Plxnb2 cKO mice. Images from 4 independent mice for each condition showed similar results.
Extended Data Fig. 2 Plexin-B2 ablation in IAM impairs wound healing.
a, Left, experimental scheme with continuous tamoxifen injection from 3 days before until 5 weeks after SCI. Right, graphs show no signification difference in body weight between cohorts during recovery after SCI. n=8 animals per group, two-way ANOVA with Bonferroni post hoc correction, n.s., p=0.11. Data represent mean ± SEM. b, IHC of sagittal thoracic spinal cords show much expanded distribution of IBA1+ cells and CSPGs, detected by CS-56 antibody, in Plxnb2 cKO as compared to control mice. Note the same spinal cord tissue was used for these co-staining as in Fig. 4b. c, IHC of sagittal thoracic spinal cords show expanded territory of pericytes (PDGFRβ+) together with IBA1+ immune cells as well as CSPG deposits in spatial disarray at the lesion site in Plxnb2 cKO as compared to control mice at 35 dpi. d, IHC of sagittal thoracic spinal cords show engorged blood vessels in expanded territory in cKO at 35 dpi, in contrast to the largely regressed capillary at the injury penumbra in control animal. R-rostral; C-caudal; D-dorsal; V-ventral, CC-central canal. For b, c, and d, n=3 independent mice per genotype with similar results.
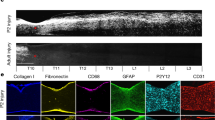
Extended Data Fig. 3 Inflammatory spillover in Plexin-B2 cKO mice after SCI.
a, Experimental scheme with continuous tamoxifen exposure from 3 days before until 5 weeks after SCI. b, IHC of sagittal thoracic spinal cords show high level of Vinculin at the lesion core, which is widespread in Plxnb2 cKO mice. Phalloidin staining also show engorged blood vessels in cKO but capillary regression in control. Images were obtained from two independent pairs of mice with similar results. c, Transcriptional profiling of ECM genes at the injury site in Plxnb2 cKO mice relative to control littermates at 14 dpi. Heatmap denotes log2 fold change (FC). Data from using two house keeping genes, Gapdh or Hsp90ab1, show similar results. The ECM genes validated in IHC are highlighted. d, Images of consecutive sagittal sections of thoracic spinal cords show widespread Oil Red O staining in Plxnb2 cKO mice at 35 dpi as compared to the confined lipid debris at the lesion core in littermate controls. Images obtained from three independent mice per group showed similar results. R-rostral; C-caudal; D-dorsal; V-ventral, CC-central canal.
Extended Data Fig. 4 Plexin-B2 ablation does not affect immune-related core function of IAM.
a, IHC images of thoracic spinal cords at 3 dpi after T8 dorsal transection (SCI-T). Mice also carry the Cx3cr1GFP reporter. EdU was delivered 3 hours before tissue harvest. Quantifications are shown on the right. Unpaired two-tailed Student’s t test. For 3 dpi, n=4 animals, n.s., p=0.62. For 7 dpi, n=4 animals, p=0.81. For 14 dpi, n=5 animals, p= 0.57. Data represent mean ± SEM. b, Images of phagocytosis assays show that both control (no hydroxy-tamoxifen) and Plxnb2 cKO (with hydroxy-tamoxifen) microglia are capable of phagocytosing fluorescent beads. n=3 independent cultures with similar results. c, Read coverage tracks from RNA-Seq data and IHC images illustrate similar upregulation of the two lipid metabolism genes in IAM in control and Plxnb2 cKO mice at three time points after SCI. Images from 3 independent mice per condition revealed similar results. d, qRT-PCR results show relative expression levels of the indicted cytokines in log2 fold change in mixed cortical cultures of the indicated genotypes. Gapdh was used as house keeping gene. n=4 independent culture samples, each performed in triplicates, unpaired two-tailed Student’s t test, p=0.43 for Tnf, **p=0.0050 for Il1b, and p=0.86 for C1qa.
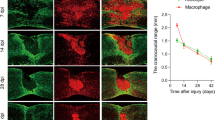
Extended Data Fig. 5 Plexin-B2 in IAM is required for corralling and vascular organization after SCI.
a, Experimental scheme. Tamoxifen was injected from day 3 before SCI until the time of analysis. (b-e) IHC images of sagittal thoracic spinal cords show early onset of the corralling process after T8 contusion injury. b, Note that in control mice at 3 dpi, dispersion of IBA1+ immune cells and intermingling with GFAP+ cells at the injury site, however, in Plxnb2 cKO mice, IBA1+ cells tended to cluster together (white arrows). c, Note the different orientation of the neovasculatures at the lesion site in Plxnb2 cKO mice, as compared to the concentric neovascular pattern in control at 7 dpi. Enlarged images of boxed area are shown on right. d, In control animal, by 7 dpi, pericytes (PDGFRβ+) had formed a ringed pattern at the injury site, in close alignment with IBA1+ cells and surrounded by CSPGs at the lesion penumbra, whereas in Plxnb2 cKO animal, pericytes had expanded their territories that mirrored the spatial disarray of IBA1+ cells, so were CSPGs. e, Note regression of capillaries at the injury site in control, but persistent engorged blood vessels in cKO animal at 14 dpi. White asterisks denote the lesion core. R-rostral; C-caudal; D-dorsal; V-ventral. Images from 2 (for d) or 3 (for b, c, and e) independent mice per genotype showed similar results.
Extended Data Fig. 6 Plexin-B2 in IAM is required for corralling and matrix reorganization after SCI.
a, Experimental scheme. Tamoxifen was injected from day 3 before until 2 weeks after SCI. (b–e) Representative IHC images of sagittal thoracic spinal cords at 14 dpi stained for the indicated genes show impaired corralling and matrix disorganization in Plxnb2 cKO mice after T8 contusion injury. Lesion site appeared enlarged and C1q widespread in cKO mice. White dashed lines denote lesion border. R-rostral; C-caudal; D-dorsal; V-ventral. For b, c, d and e, images obtained from 3 independent mice per genotype showed similar results.
Extended Data Fig. 7 Plexin-B2 in IAM is critical for corralling and injury resolution during the early phase of recovery from SCI.
a, Left, experimental scheme. Tamoxifen was injected from day 3 before until 3 weeks after SCI. Middle, images of sagittal section of thoracic spinal cords show confined Oil Red O staining in control but diffuse in Plexin-B2 cKO mice at 35 dpi. Quantifications are shown on the right, n=3 animals per group, unpaired two-tailed Students t-test, **p=0.0015. Graphs represent mean ± SEM. b–d, IHC of sagittal thoracic spinal cords at 35 dpi stained for the indicated markers show impaired corralling and matrix disorganization in Plxnb2 cKO mice. b, White dashed lines denote lesion border. c, Note widespread CD68+ cells (white arrows) in Plxnb2 cKO mice at 35 dpi. Enlarged images of boxed areas are shown on the right. d, Note with this tamoxifen regimen, in Plxnb2 cKO mice at 35 dpi, re-expression of Plexin-B2 in newly infiltrated macrophages at the injury site, while microglia continue to have Plexin-B2 ablation (white arrowhead). R-rostral; C-caudal; D-dorsal; V-ventral. For b, c, and d, n=3 independent mice per genotype with similar results.
Extended Data Fig. 8 Sema4C knockout minimally impacts functional recovery and wound healing after SCI.
a, Top, experimental scheme. b, BMS scores during the 5 week recovery after T8 contusion injury demonstrate comparably functional recovery in Sema4C KO mice as compared to littermate controls. n=6 for control cohort, n=4 for cKO cohort. Two-way ANOVA with Bonferroni post hoc correction for repeated measures. n.s., p=0.72. Data represent mean ± SEM. c, Rotarod test showed a modest decrease of hindlimb and tail balance in Sema4C KO mice as compared to littermate controls (n=5 for control cohort, n=4 for cKO cohort). Unpaired two-tailed Student’s t test. For duration, *p=0.046; for speed, *p=0.046. Von Frey filament test showed no significant difference in tactile sensory function between genotypes (n=5 for control cohort, n=4 for cKO cohort). For each animal, left and right hindpaws measured separately. Unpaired two-tailed Student’s t test. p=0.79. Data represent mean ± SEM. d, IHC images of sagittal thoracic spinal cord at 35 dpi stained for the indicated markers show successfully corralling and wound compaction in both cohorts after T8 contusion injury. Images from 3 animals per genotype showed similar results. R-rostral; C-caudal; D-dorsal; V-ventral.
Supplementary information
Supplementary Video 1
Open-field BMS scoring for control animals. Open-field walking test for BMS scoring of control animals at 35 days post injury. Animals were scored for plantar placement, ankle movement, angle of hindlimbs, trunk stability and other parameters as outlined in the BMS guidelines as previously described. One representative movie from 12 independent animals is shown.
Supplementary Video 2
Open-field BMS scoring for Plxnb2 cKO animals. Open-field walking test for BMS scoring of a Plxnb2 cKO animals at 35 days post injury. One representative movie from 12 independent animals is shown.
Supplementary Video 3
Phagocytosis assay for control microglia. pHrodo Red Zymosan Bioparticles were added to a lawn of microglia isolated from Cx3cr1CreER/GFPPlxnb2fl/B mice without tamoxifen treatment (control). Microglia comparably phagocytosed the beads in the control and the cKO condition. IncuCyte time-lapse movies were acquired at 1-h intervals for a period of 24 h. n = 3 independent cultures per condition, and similar results were obtained.
Supplementary Video 4
Phagocytosis assay for Plxnb2 cKO microglia. pHrodo Red Zymosan Bioparticles were added to a lawn of microglia isolated from Cx3cr1CreER/GFPPlxnb2fl/B mice with tamoxifen treatment (cKO). Microglia comparably phagocytosed the beads in the control and the cKO condition. IncuCyte time-lapse movies were acquired at 1-h intervals for a period of 24 h. n = 3 independent cultures per condition, and similar results were obtained.
Supplementary Video 5
Cell motility for control microglia. Control microglia (GFP+) are highly motile. Primary microglia from the cortex of neonatal Cx3cr1CreER/GFPPlxnb2fl/B pups were collected and cultured. Cx3cr1-GFP+ microglia were overlaid on a lawn of Cx3cr1-GFP– neural cells (labeled with CellTracker Red). IncuCyte time-lapse movie was acquired at 30-min intervals for a duration of 5 days. Microglia motility (red line) was tracked using the MTrackJ plugin from ImageJ. n = 3 independent cultures showed similar results.
Supplementary Video 6
Cell motility for Plxnb2 cKO microglia. Plxnb2 cKO microglia (GFP+) tracked on a lawn of neural cells labeled with CellTracker Red, as in Supplementary Video 5. n = 3 independent cultures showed similar results.
Supplementary Video 7
CIL of control microglia. An IncuCyte time-lapse movie was acquired at 5-min time intervals over a period of 2 h. Only a 30-min clip is shown here. Cortical cultures were obtained from neonatal Cx3cr1CreER/GFPPlxnb2fl/B mice and were not treated with OHT (control). Microglia were visualized using Cx3cr1-GFP reporter labels. Control microglia retracted and moved away within 5 min of contact with other cell types. n = 3 independent cultures showed similar results.
Supplementary Video 8
CIL of Plxnb2 cKO microglia. An IncuCyte time-lapse movie was acquired at 5-min time intervals over a period of 2 h. Only a 30-min clip is shown here. Cortical cultures were obtained from neonatal Cx3cr1CreER/GFPPlxnb2fl/B mice and treated with OHT (Plxnb2 cKO). Microglia were visualized using a Cx3cr1-GFP reporter label. Mutant cKO microglia stayed longer in contact with other cells than in control conditions. n = 3 independent cultures showed similar results.
Rights and permissions
About this article

Cite this article
Zhou, X., Wahane, S., Friedl, MS. et al. Microglia and macrophages promote corralling, wound compaction and recovery after spinal cord injury via Plexin-B2. Nat Neurosci 23, 337–350 (2020). https://doi.org/10.1038/s41593-020-0597-7
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-020-0597-7
This article is cited by
-
M2 microglia-derived exosome-loaded electroconductive hydrogel for enhancing neurological recovery after spinal cord injury
Journal of Nanobiotechnology (2024)
-
S100A9 Induces Macrophage M2 Polarization and Immunomodulatory Role in the Lesion Site After Spinal Cord Injury in Rats
Molecular Neurobiology (2024)
-
IFN-γ-STAT1-mediated CD8+ T-cell-neural stem cell cross talk controls astrogliogenesis after spinal cord injury
Inflammation and Regeneration (2023)
-
Microglial SIRT1 activation attenuates synapse loss in retinal inner plexiform layer via mTORC1 inhibition
Journal of Neuroinflammation (2023)
-
Microglia/macrophages are ultrastructurally altered by their proximity to spinal cord injury in adult female mice
Journal of Neuroinflammation (2023)
