
Abstract
Innate immune cells recruited to inflammatory sites have short life spans and originate from the marrow, which is distributed throughout the long and flat bones. While bone marrow production and release of leukocyte increases after stroke, it is currently unknown whether its activity rises homogeneously throughout the entire hematopoietic system. To address this question, we employed spectrally resolved in vivo cell labeling in the murine skull and tibia. We show that in murine models of stroke and aseptic meningitis, skull bone marrow-derived neutrophils are more likely to migrate to the adjacent brain tissue than cells that reside in the tibia. Confocal microscopy of the skull–dura interface revealed myeloid cell migration through microscopic vascular channels crossing the inner skull cortex. These observations point to a direct local interaction between the brain and the skull bone marrow through the meninges.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

References
Swirski, F. K. & Nahrendorf, M. Leukocyte behavior in atherosclerosis, myocardial infarction, and heart failure. Science 339, 161–166 (2013).
Jickling, G. C. et al. Targeting neutrophils in ischemic stroke: translational insights from experimental studies. J. Cereb. Blood Flow Metab. 35, 888–901 (2015).
Kamel, H. & Iadecola, C. Brain-immune interactions and ischemic stroke: clinical implications. Arch. Neurol. 69, 576–581 (2012).
Offner, H. et al. Experimental stroke induces massive, rapid activation of the peripheral immune system. J. Cereb. Blood Flow Metab. 26, 654–665 (2006).
Courties, G., Moskowitz, M. A. & Nahrendorf, M. The innate immune system after ischemic injury: lessons to be learned from the heart and brain. JAMA Neurol. 71, 233–236 (2014).
Gelderblom, M., Sobey, C. G., Kleinschnitz, C. & Magnus, T. Danger signals in stroke. Ageing Res. Rev. 24, 77–82 (2015).
Courties, G. et al. Ischemic stroke activates hematopoietic bone marrow stem cells. Circ. Res. 116, 407–417 (2015).
Morrison, S. J. & Scadden, D. T. The bone marrow niche for haematopoietic stem cells. Nature 505, 327–334 (2014).
Itkin, T. et al. Distinct bone marrow blood vessels differentially regulate haematopoiesis. Nature 532, 323–328 (2016).
Gamache, D. A., Povlishock, J. T. & Ellis, E. F. Carrageenan-induced brain inflammation. Characterization of the model. J. Neurosurg. 65, 679–685 (1986).
Ueda, Y., Kondo, M. & Kelsoe, G. Inflammation and the reciprocal production of granulocytes and lymphocytes in bone marrow. J. Exp. Med. 201, 1771–1780 (2005).
Hill, W. D. et al. SDF-1 (CXCL12) is upregulated in the ischemic penumbra following stroke: association with bone marrow cell homing to injury. J. Neuropathol. Exp. Neurol. 63, 84–96 (2004).
Heidt, T. et al. Chronic variable stress activates hematopoietic stem cells. Nat. Med. 20, 754–758 (2014).
Zenker, W. & Kubik, S. Brain cooling in humans–anatomical considerations. Anat. Embryol. (Berl.) 193, 1–13 (1996).
Adeeb, N., Mortazavi, M. M., Tubbs, R. S. & Cohen-Gadol, A. A. The cranial dura mater: a review of its history, embryology, and anatomy. Childs Nerv. Syst. 28, 827–837 (2012).
Hershkovitz, I. et al. The elusive diploic veins: anthropological and anatomical perspective. Am. J. Phys. Anthropol. 108, 345–358 (1999).
Rangel de Lázaro, G., de la Cuétara, J. M., Píšová, H., Lorenzo, C. & Bruner, E. Diploic vessels and computed tomography: segmentation and comparison in modern humans and fossil hominids. Am. J. Phys. Anthropol. 159, 313–324 (2016).
Lucchinetti, C. F. et al. Inflammatory cortical demyelination in early multiple sclerosis. N. Engl. J. Med. 365, 2188–2197 (2011).
Michaud, J. P., Bellavance, M. A., Préfontaine, P. & Rivest, S. Real-time in vivo imaging reveals the ability of monocytes to clear vascular amyloid beta. Cell Rep. 5, 646–653 (2013).
Prokop, S. et al. Impact of peripheral myeloid cells on amyloid-β pathology in Alzheimer’s disease-like mice. J. Exp. Med. 212, 1811–1818 (2015).
Zenaro, E. et al. Neutrophils promote Alzheimer’s disease-like pathology and cognitive decline via LFA-1 integrin. Nat. Med. 21, 880–886 (2015).
Park, L. et al. Brain perivascular macrophages initiate the neurovascular dysfunction of Alzheimer Aβ peptides. Circ. Res. 121, 258–269 (2017).
Faraco, G., Park, L., Anrather, J. & Iadecola, C. Brain perivascular macrophages: characterization and functional roles in health and disease. J. Mol. Med. (Berl.) 95, 1143–1152 (2017).
Abtin, A. et al. Perivascular macrophages mediate neutrophil recruitment during bacterial skin infection. Nat. Immunol. 15, 45–53 (2014).
Perez-de-Puig, I. et al. Neutrophil recruitment to the brain in mouse and human ischemic stroke. Acta Neuropathol. 129, 239–257 (2015).
Bartholomäus, I. et al. Effector T cell interactions with meningeal vascular structures in nascent autoimmune CNS lesions. Nature 462, 94–98 (2009).
Coles, J. A., Myburgh, E., Brewer, J. M. & McMenamin, P. G. Where are we? The anatomy of the murine cortical meninges revisited for intravital imaging, immunology, and clearance of waste from the brain. Prog. Neurobiol. 156, 107–148 (2017).
Louveau, A. et al. Structural and functional features of central nervous system lymphatic vessels. Nature 523, 337–341 (2015).
Aspelund, A. et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 212, 991–999 (2015).
McKittrick, C. M., Lawrence, C. E. & Carswell, H. V. Mast cells promote blood brain barrier breakdown and neutrophil infiltration in a mouse model of focal cerebral ischemia. J. Cereb. Blood Flow Metab. 35, 638–647 (2015).
Arac, A. et al. Evidence that meningeal mast cells can worsen stroke pathology in mice. Am. J. Pathol. 184, 2493–2504 (2014).
Sellner, J. & Leib, S. L. In bacterial meningitis cortical brain damage is associated with changes in parenchymal MMP-9/TIMP-1 ratio and increased collagen type IV degradation. Neurobiol. Dis. 21, 647–656 (2006).
Kruger, P. et al. Neutrophils: between host defence, immune modulation, and tissue injury. PLoS Pathog. 11, e1004651 (2015).
Shi, Y., Leak, R. K., Keep, R. F. & Chen, J. Translational stroke research on blood-brain barrier damage: challenges, perspectives, and goals. Transl. Stroke Res. 7, 89–92 (2016).
Keenan, T. M. & Folch, A. Biomolecular gradients in cell culture systems. Lab Chip 8, 34–57 (2008).
Lelios, I. & Greter, M. Isolation of leukocytes from mouse central nervous system. Methods Mol. Biol. 1193, 15–19 (2014).
Kim, J. Y. et al. Direct imaging of cerebral thromboemboli using computed tomography and fibrin-targeted gold nanoparticles. Theranostics 5, 1098–1114 (2015).
Acknowledgements
The authors thank M. Ericsson (HMS Electron Microscopy Facility) for skull sample preparation, sectioning, and assistance with EM imaging. We acknowledge D. Capen (Center for Systems Biology and Program in Membrane Biology/Division of Nephrology, MGH) for help with interpretation of electron microscopy data. The authors thank the MGH mouse imaging program and the Center for Skeletal Research Core (NIH P30 AR066261) for assistance with imaging. This work was funded in part by grants from the National Institutes of Health (NS084863 and HL139598), the American Heart Association (16SDG30190009), the Cure Alzheimer’s Fund, the Global Research Lab (GRL) program (NRF-2015K1A1A2028228) of the National Research Foundation by the Korean government, and by fellowships from the Netherlands Organisation for Scientific Research (NWO, Rubicon Grant: 835.15.014), the Deutsche Forschungsgemeinschaft (RO5071/1-1), and the MGH Research Scholar program.
Author information
Authors and Affiliations
Contributions
F.H. conceived the study, designed, performed, and analyzed most experiments and contributed to writing the manuscript. V.F., G.C., D.R., Y.S., K.V., G.R.W., J. K., C.V., and G.S.M. performed flow cytometry and imaging experiments. D.E.K., R.W., F.K.S., and M.A.M. discussed and interpreted data, provided strategic input, raised funding, and edited the manuscript. M.N. supervised the study and wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Bone marrow cell-tagging effect on cell viability or function.
a, Neutrophil recruitment to the brain after intravenous or marrow injection of cell tracker. Two-tailed Mann-Whitney test, naive (n = 7 IV, n = 6 skull tag, 4 experiments), P = 0.52; stroke 6hrs (n = 3 IV, n = 8 skull tag, 3 experiments), P = 0.28; stroke 24hrs (n = 3 IV, n = 6 skull tag, 3 experiments), P = 0.71; b, Viability (upper panel) and cellularity (lower panel) of bone marrow after intravenous or marrow injection of the red (skull) and green (tibia) dyes in naive mice (n = 7 IV, n = 8 Tag, 6 experiments). Two-tailed Mann Whitney test, viability skull, P = 0.01; viability tibia, P = 0.02; cellularity skull P > 0.99; cellularity tibia, P = 0.61. c, Gating for microglia and uptake of red cell tracker 4 hrs after cisternal carrageenan injection (single experiment), after subdural injection of 10 µl (upper left), no tracker (bottom left), 10 µl in the calvarium (upper right), IV injection of 10 µl (bottom right). Data are mean ± s.e.m.. Mann-Whitney test, ns indicates not significant. d, Brain sections of a naive Cx3cr1GFP mouse, after 10 µl red cell tracker injected in the sub-dural area (upper panel, arrow shows injection site) or locally in the skull marrow (lower panel), single experiment.
Supplementary Figure 2 Flow cytometry gating for brain.
Bottom right panel shows gates for cells originating from tibia (green tracker, FITC channel) and skull (red tracker, APC channel) based on the signal obtained in the circulation after IV injection (bottom left panel).
Supplementary Figure 3 Additional cell-tracking analyses.
a-c, Frequency of cells tracked from skull and tibia marrow in (a) stroke, 6hrs, n = 11, 5 experiments; 1 day, n = 13 for brain and spleen and n = 12 for blood, 5 experiments; 2 days, n = 7, 2 experiments; skull ***P = 0.001 and **P = 0.002 at 6hrs, *P = 0.022, *P = 0.03 at day 1; *P = 0.03, ns P = 0.22 at 2 days; tibia ns P = 0.46 at 6hrs, ns P = 0.69 at day 1, (b) carrageenan, n = 6, 4 experiments, Kruskall Wallis test, skull P = 0.51, tibia ns P = 0.65, (c) myocardial infarction, n = 5, 1 experiment, Kruskall Wallis test skull ns P = 0.11, tibia ns P = 0.32. d-f, Frequency of tracked cells in respective organs relative to the circulation after (d) stroke, at 6 hrs, n = 11, 5 experiments, ***P = 0.002, 1 day, n = 12, 5 experiments, P = 0.064 and 2 days, n = 5, 2 experiments, ns P = 0.13, (e) carrageenan injection, n = 6, 4 experiments and (f) after myocardial infarction, n = 5, 1 experiment. Data are mean ± s.e.m., ns indicates not significant, two-tailed Wilcoxon test unless otherwise specified.
Supplementary Figure 4 Location of tracked cells from skull and tibia after aseptic meningitis.
Cells originating from skull (red, arrow) and tibia (green, arrow head) in brain one day after induction of aseptic meningitis (n = 2 mice). Cells are outside (a,b,d) or inside (c) the vasculature. Speckles in c are present in all channels and present autofluorescence. Collapsed Z stacks show cells at a depth of 20-100 µm below the brain tissue surface.
Supplementary Figure 5 The spine marrow during acute stroke.
N = 12 per group, 3 experiments. Data are mean ± s.e.m. Two-tailed Mann-Whitney test, neutrophils, P = 0.29 and Ly6Chi monocytes, P = 0.052.
Supplementary Figure 6 Competitive in vitro transmigration assay through activated endothelium comparing skull and tibial neutrophils.
Cells from 6 mice (6 skull-tibia pairs, red or green fluorescence staining for location-specific tracking) were subjected to migration through a TNFα-activated brain endothelium towards fMLP. Results are migrated cells as % of initial population. Data are mean ± s.e.m.. P = 0.69, two-tailed Wilcoxon test.
Supplementary Figure 7 Histology of the dural vasculature 1 d after stroke.
Dural vasculature one day after stroke induced by permanent occlusion (representative of two experiments). Arrows show cells inside vasculature, arrow heads indicate cells outside of vasculature.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–7
Supplementary Video 1 - Neutrophils exiting a channel in vitro after intracisternal carrageenan injection.
Representative of 4 experiments. Channel exits are indicated with arrows. Dashed arrows indicate meningeal blood vessels linked to the channel. The bone marrow cavity is indicated by an asterisk. Scale bar indicates 50µm.
Supplementary Video 2 - Neutrophils exiting a channel in vivo.
In vivo imaging of a channel crossing the interior skull cortex in a 4 week old mouse one day after stroke (single experiment). Arrow indicates the cell’s direction towards the brain. Scale bar indicates 50µm.
Supplementary Video 3 - Neutrophils exiting a channel in vivo.
As in supplementary movie 2, but 6hrs after permanent middle cerebral artery occlusion (single experiment). Scale bar indicates 50µm.
Supplementary Video 4 - Neutrophil migration after sham surgery.
This movie was obtained in vitro from a sham control and shows no cell exit. Representative of 4 experiments. Scale bar indicates 50µm. Arrow indicates the exit of the channel at the dural. The asterisk indicates skull bone marrow.
Supplementary Video 5 - Neutrophil migration through channel in vitro after stroke.
As Supplementary movie 4, recorded in vitro after stroke. Scale bar indicates 50µm.
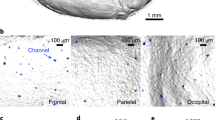
Supplementary Video 6 - Computed tomography of channels.
Animation of a high resolution data set shows mouse skull surface rendering (single experiment). Toward the end of the movie, arrows indicate microscopic channels.
Rights and permissions
About this article

Cite this article
Herisson, F., Frodermann, V., Courties, G. et al. Direct vascular channels connect skull bone marrow and the brain surface enabling myeloid cell migration. Nat Neurosci 21, 1209–1217 (2018). https://doi.org/10.1038/s41593-018-0213-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-018-0213-2
This article is cited by
-
Peripheral blood amyloid-β involved in the pathogenesis of Alzheimer’s disease via impacting on peripheral innate immune cells
Journal of Neuroinflammation (2024)
-
Targeting brain-peripheral immune responses for secondary brain injury after ischemic and hemorrhagic stroke
Journal of Neuroinflammation (2024)
-
Role of meningeal immunity in brain function and protection against pathogens
Journal of Inflammation (2024)
-
In vivo NIR-II fluorescence imaging for biology and medicine
Nature Photonics (2024)
-
Beyond the microcirculation: sequestration of infected red blood cells and reduced flow in large draining veins in experimental cerebral malaria
Nature Communications (2024)
