
Abstract
The medial prefrontal cortex (mPFC) has been implicated in the extinction of emotional memories, including conditioned fear. We found that ventral hippocampal (vHPC) projections to the infralimbic (IL) cortex recruited parvalbumin-expressing interneurons to counter the expression of extinguished fear and promote fear relapse. Whole-cell recordings ex vivo revealed that optogenetic activation of vHPC input to amygdala-projecting pyramidal neurons in the IL was dominated by feed-forward inhibition. Selectively silencing parvalbumin-expressing, but not somatostatin-expressing, interneurons in the IL eliminated vHPC-mediated inhibition. In behaving rats, pharmacogenetic activation of vHPC→IL projections impaired extinction recall, whereas silencing IL projectors diminished fear renewal. Intra-IL infusion of GABA receptor agonists or antagonists, respectively, reproduced these effects. Together, our findings describe a previously unknown circuit mechanism for the contextual control of fear, and indicate that vHPC-mediated inhibition of IL is an essential neural substrate for fear relapse.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others
Change history
09 July 2018
In the version of this article initially published, the traces in Fig. 1j and in Fig. 1k, right, were duplicated from the corresponding traces in Fig. 1c, bottom, and Fig. 1d, bottom right. The error has been corrected in the HTML and PDF versions of the article.
References
Quirk, G. J. & Milad, M. R. Neuroscience: editing out fear. Nature 463, 36–37 (2010).
Milad, M. R. & Quirk, G. J. Fear extinction as a model for translational neuroscience: ten years of progress. Annu. Rev. Psychol. 63, 129–151 (2012).
Goode, T. D. & Maren, S. Animal models of fear relapse. ILAR J. 55, 246–258 (2014).
Vervliet, B., Craske, M. G. & Hermans, D. Fear extinction and relapse: state of the art. Annu. Rev. Clin. Psychol. 9, 215–248 (2013).
Bouton, M. E. Context and behavioral processes in extinction. Learn. Mem. 11, 485–494 (2004).
Bouton, M. E. & Bolles, R. C. Contextual control of the extinction of conditioned fear. Learn. Motiv. 10, 445–466 (1979).
Laurent, V. & Westbrook, R. F. Inactivation of the infralimbic but not the prelimbic cortex impairs consolidation and retrieval of fear extinction. Learn. Mem. 16, 520–529 (2009).
Milad, M. R. & Quirk, G. J. Neurons in medial prefrontal cortex signal memory for fear extinction. Nature 420, 70–74 (2002).
Morgan, M. A., Romanski, L. M. & LeDoux, J. E. Extinction of emotional learning: contribution of medial prefrontal cortex. Neurosci. Lett. 163, 109–113 (1993).
Likhtik, E., Pelletier, J. G., Paz, R. & Paré, D. Prefrontal control of the amygdala. J. Neurosci. 25, 7429–7437 (2005).
Cho, J.-H., Deisseroth, K. & Bolshakov, V. Y. Synaptic encoding of fear extinction in mPFC-amygdala circuits. Neuron 80, 1491–1507 (2013).
Knapska, E. & Maren, S. Reciprocal patterns of c-Fos expression in the medial prefrontal cortex and amygdala after extinction and renewal of conditioned fear. Learn. Mem. 16, 486–493 (2009).
Orsini, C. A., Kim, J. H., Knapska, E. & Maren, S. Hippocampal and prefrontal projections to the basal amygdala mediate contextual regulation of fear after extinction. J. Neurosci. 31, 17269–17277 (2011).
Maren, S., Phan, K. L. & Liberzon, I. The contextual brain: implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 14, 417–428 (2013).
Corcoran, K. A. & Maren, S. Hippocampal inactivation disrupts contextual retrieval of fear memory after extinction. J. Neurosci. 21, 1720–1726 (2001).
Hobin, J. A., Ji, J. & Maren, S. Ventral hippocampal muscimol disrupts context-specific fear memory retrieval after extinction in rats. Hippocampus 16, 174–182 (2006).
Zelikowsky, M. et al. Prefrontal microcircuit underlies contextual learning after hippocampal loss. Proc. Natl. Acad. Sci. USA 110, 9938–9943 (2013).
Jin, J. & Maren, S. Fear renewal preferentially activates ventral hippocampal neurons projecting to both amygdala and prefrontal cortex in rats. Sci. Rep. 5, 8388 (2015).
Herry, C. et al. Switching on and off fear by distinct neuronal circuits. Nature 454, 600–606 (2008).
Knapska, E. et al. Functional anatomy of neural circuits regulating fear and extinction. Proc. Natl. Acad. Sci. USA 109, 17093–17098 (2012).
Xu, C. et al. Distinct hippocampal pathways mediate dissociable roles of context in memory retrieval. Cell 167, 961–972.e16 (2016).
Wang, Q., Jin, J. & Maren, S. Renewal of extinguished fear activates ventral hippocampal neurons projecting to the prelimbic and infralimbic cortices in rats. Neurobiol. Learn. Mem. 134, 38–43 (2016).
Parent, M. A., Wang, L., Su, J., Netoff, T. & Yuan, L.-L. Identification of the hippocampal input to medial prefrontal cortex in vitro. Cereb. Cortex 20, 393–403 (2010).
Hoover, W. B. & Vertes, R. P. Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct. Funct. 212, 149–179 (2007).
Ascoli, G. A. et al. Petilla Interneuron Nomenclature Group et al. Petilla terminology: nomenclature of features of GABAergic interneurons of the cerebral cortex. Nat. Rev. Neurosci. 9, 557–568 (2008).
Ferreira, A. N., Yousuf, H., Dalton, S. & Sheets, P. L. Highly differentiated cellular and circuit properties of infralimbic pyramidal neurons projecting to the periaqueductal gray and amygdala. Front. Cell. Neurosci. 9, 161 (2015).
Little, J. P. & Carter, A. G. Synaptic mechanisms underlying strong reciprocal connectivity between the medial prefrontal cortex and basolateral amygdala. J. Neurosci. 33, 15333–15342 (2013).
Kawaguchi, Y. & Kubota, Y. Neurochemical features and synaptic connections of large physiologically-identified GABAergic cells in the rat frontal cortex. Neuroscience 85, 677–701 (1998).
Tamás, G., Lorincz, A., Simon, A. & Szabadics, J. Identified sources and targets of slow inhibition in the neocortex. Science 299, 1902–1905 (2003).
Wozny, C. & Williams, S. R. Specificity of synaptic connectivity between layer 1 inhibitory interneurons and layer 2/3 pyramidal neurons in the rat neocortex. Cereb. Cortex 21, 1818–1826 (2011).
Kawaguchi, Y. & Kondo, S. Parvalbumin, somatostatin and cholecystokinin as chemical markers for specific GABAergic interneuron types in the rat frontal cortex. J. Neurocytol. 31, 277–287 (2002).
Bukalo, O. et al. Prefrontal inputs to the amygdala instruct fear extinction memory formation. Sci. Adv. 1, e1500251 (2015).
Duvarci, S. & Pare, D. Amygdala microcircuits controlling learned fear. Neuron 82, 966–980 (2014).
Gomez, J. L. et al. Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 357, 503–507 (2017).
Do-Monte, F. H., Manzano-Nieves, G., Quiñones-Laracuente, K., Ramos-Medina, L. & Quirk, G. J. Revisiting the role of infralimbic cortex in fear extinction with optogenetics. J. Neurosci. 35, 3607–3615 (2015).
Ishikawa, A. & Nakamura, S. Convergence and interaction of hippocampal and amygdalar projections within the prefrontal cortex in the rat. J. Neurosci. 23, 9987–9995 (2003).
Tierney, P. L., Dégenètais, E., Thierry, A.-M., Glowinski, J. & Gioanni, Y. Influence of the hippocampus on interneurons of the rat prefrontal cortex. Eur. J. Neurosci. 20, 514–524 (2004).
Sotres-Bayon, F., Sierra-Mercado, D., Pardilla-Delgado, E. & Quirk, G. J. Gating of fear in prelimbic cortex by hippocampal and amygdala inputs. Neuron 76, 804–812 (2012).
Padilla-Coreano, N. et al. Direct ventral hippocampal-prefrontal input is required for anxiety-related neural activity and behavior. Neuron 89, 857–866 (2016).
Jin, J. & Maren, S. Prefrontal-hippocampal interactions in memory and emotion. Front. Syst. Neurosci. 9, 170 (2015).
Shin, L. M., Rauch, S. L. & Pitman, R. K. Amygdala, medial prefrontal cortex, and hippocampal function in PTSD. Ann. NY Acad. Sci. 1071, 67–79 (2006).
Herry, C. et al. Neuronal circuits of fear extinction. Eur. J. Neurosci. 31, 599–612 (2010).
Giustino, T. F. & Maren, S. The role of the medial prefrontal cortex in the conditioning and extinction of fear. Front. Behav. Neurosci. 9, 298 (2015).
Marek, R., Strobel, C., Bredy, T. W. & Sah, P. The amygdala and medial prefrontal cortex: partners in the fear circuit. J. Physiol. (Lond.) 591, 2381–2391 (2013).
Adhikari, A. et al. Basomedial amygdala mediates top-down control of anxiety and fear. Nature 527, 179–185 (2015).
Kim, H.-S., Cho, H.-Y., Augustine, G. J. & Han, J.-H. Selective control of fear expression by optogenetic manipulation of infralimbic cortex after extinction. Neuropsychopharmacology 41, 1261–1273 (2016).
Maren, S. Fear of the unexpected: hippocampus mediates novelty-induced return of extinguished fear in rats. Neurobiol. Learn. Mem. 108, 88–95 (2014).
Knight, R. Contribution of human hippocampal region to novelty detection. Nature 383, 256–259 (1996).
Eichenbaum, H. Prefrontal-hippocampal interactions in episodic memory. Nat. Rev. Neurosci. 18, 547–558 (2017).
Adhikari, A., Topiwala, M. A. & Gordon, J. A. Synchronized activity between the ventral hippocampus and the medial prefrontal cortex during anxiety. Neuron 65, 257–269 (2010).
Armbruster, B. N., Li, X., Pausch, M. H., Herlitze, S. & Roth, B. L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 104, 5163–5168 (2007).
Lerchner, W. et al. Reversible silencing of neuronal excitability in behaving mice by a genetically targeted, ivermectin-gated Cl- channel. Neuron 54, 35–49 (2007).
Lynagh, T. & Lynch, J. W. An improved ivermectin-activated chloride channel receptor for inhibiting electrical activity in defined neuronal populations. J. Biol. Chem. 285, 14890–14897 (2010).
Klapoetke, N. C. et al. Independent optical excitation of distinct neural populations. Nat. Methods 11, 338–346 (2014).
Han, X. et al. A high-light sensitivity optical neural silencer: development and application to optogenetic control of non-human primate cortex. Front. Syst. Neurosci. 5, 18 (2011).
Strobel, C., Marek, R., Gooch, H.M., Sullivan, R.K.P. & Sah, P. Prefrontal and auditory input to intercalated neurons of the amygdala. Cell Reports https://doi.org/10.1016/j.celrep.2015.02.008 (2015).
Fitzgerald, P. J., Giustino, T. F., Seemann, J. R. & Maren, S. Noradrenergic blockade stabilizes prefrontal activity and enables fear extinction under stress. Proc. Natl. Acad. Sci. USA 112, E3729–E3737 (2015).
Corcoran, K. A., Desmond, T. J., Frey, K. A. & Maren, S. Hippocampal inactivation disrupts the acquisition and contextual encoding of fear extinction. J. Neurosci. 25, 8978–8987 (2005).
Giustino, T. F. et al. β-adrenoceptor blockade in the basolateral amygdala, but not the medial prefrontal cortex, rescues the immediate extinction deficit. Neuropsychopharmacology 42, 2537–2544 (2017).
Acknowledgements
We thank A. Woodruff for comments on the manuscript. We thank L. Xu, University of North Carolina Vector Core, University of Pennsylvania Vector Core, and the Institute of Molecular Genetics of Montpellier for producing viruses. This work was supported by the US National Institutes of Health (R01MH065961 to S.M.; F31MH107113 to T.D.G.; F31MH112208 to T.F.G.), a McKnight Memory and Cognitive Disorders Award to S.M., and Australian Research Council (CE140100007) and National Health and Medical Research Grants to P.S.
Author information
Authors and Affiliations
Contributions
S.M. and P.S. supervised all of the experiments. S.M., P.S. and R.M. designed the experiments. R.M., J.J., T.D.G., T.F.G., Q.W., G.M.A. and P.J.F. collected the data. R.M., J.J., T.D.G., T.F.G., Q.W., G.M.A., P.J.F., S.M. and P.S. analyzed the data. R.H. and J.E.P. generated and provided AAVDJ/8 viral vectors. T.L. and J.W.L. generated and provided the ivermectin construct. R.M., J.J., T.D.G., S.M. and P.S. wrote the manuscript. All of the authors read and edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
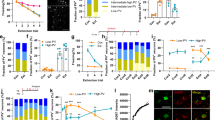
Supplementary Figure 1 HPC projections predominantly target IL neurons in the mPFC.
Comparison of hippocampal inputs onto principal neurons in the IL and PL reveal significantly larger EPSCs in IL neurons compared to caudal or rostral PL responses (rostral PL: n = 12; caudal PL: n = 5; IL: n = 21 from 7 animals; one-way ANOVA with Dunnett’s multiple comparison test, F2,35 = 6.62, *P = 0.037). Dots represent data from individual cells. Error bars indicate means ± s.e.m.
Supplementary Figure 2 Hippocampus-driven IL responses are monosynaptic, time-locked responses.
a, Example of a HPC-driven response in a pyramidal neuron, containing the initial excitatory response, followed by an inhibitory component. The 5-fold enlarged inset shows the time-locked onset of individual responses (grey) to the light stimulation (blue) without any response-failures. Averaged response shown in black. Analysis of EPSCs show very low response jitter (b) (individual jitter of neurons shown as grey dots; n = 19) and response latency (c) (n = 14), both typical parameters for monosynaptic responses. Error bars indicate means ± s.e.m.
Supplementary Figure 3 Quantification of vHPC-evoked inhibitory conductances in IL principal cells.
a, Schematic showing experimental setup. Optical terminal stimulation of ventral hippocampal inputs and whole-cell recording of pyramidal neurons in the IL. For the spiking suppression experiments (d), local electrical stimulation (battery symbol) was used. b, Current-clamp responses to hippocampal terminal release before (black trace) and after (red trace) the application of the GABAB-receptor antagonist CGP55845 (n = 3), revealing the fast (yellow) and slow IPSCs (green). c, Bar graphs with response patterns of IL principal neurons to optical hippocampal stimulation containing fast (top, yellow) and slow (bottom, green) inhibitory conductances (percentage of the total). d, Investigation of spiking suppression of IL principal neurons by the hippocampus. Electrical suprathreshold stimulation was used to evoke spiking (left) while presenting optical hippocampal stimulation 150 ms before the spiking event (n = 8). Blue bars represent optical stimulation.
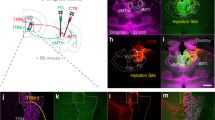
Supplementary Figure 4 Feed-forward inhibition onto pyramidal neurons is mediated by local IL interneurons.
a, Schematic for electrical stimulation (battery symbol) of IL tissue ex vivo in the presence of AMPA- and NMDA-receptor antagonists NBQX and APV, respectively, to isolate GABAergic transmission. b, Voltage clamp (left) and current-clamp recordings (right) revealed inhibitory responses that contain both fast (yellow) and slow (green) inhibitory components. c, Application of the GABAB-receptor antagonist CGP55845 (red trace) isolated the fast, inhibitory conductance, which was blocked by the GABAA/C-receptor antagonist picrotoxin (green trace). Holding voltage: −60 mV.
Supplementary Figure 5 CNO- and virus-dependent silencing of IL neurons in freely moving rats.
a, Schematic design of experimental approach alongside a representative image depicting DREADD-expressing neurons of animals implanted with multichannel recording arrays into the IL (40 μm coronal section; white bar inset = 250 μm). b, Vehicle (VEH) injections did not cause a significant change in the spontaneous activity (20 s bins) of IL neurons (n = 15 for hM4Di; n = 27 for mCherry control) (left). When the hM4D(Gi)-expressing animal was injected with 1 mg/kg (middle) or 3 mg/kg (right) of CNO, IL neurons (n = 18 for 1 mg/kg; n = 16 for 3 mg/kg) exhibited a significant reduction in spontaneous firing relative to neurons of control virus-infected animals (n = 25 for 1 mg/kg; n = 23 for 3 mg/kg; repeated measures ANOVA, main effects of virus: F1,41 = 10.912, **P = 0.0020 for 1 mg/kg; F1,37 = 24.375, ***P < 0.0001 for 3 mg/kg).
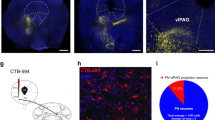
Supplementary Figure 6 Pharmacological inactivation of the IL impedes retrieval of extinguished fear.
Test data show mean baseline freezing (3 min), mean freezing during nine 5-ITI blocks (30-sec ITIs) and a during a post-trial period (150 sec) following infusions of muscimol or vehicle into the IL (MUSC, n = 6; VEH, n = 10; repeated measures ANOVA, main effect of drug: F1,14 = 35.78, ***P < 0.0001). Corresponding conditioning and extinction data are shown in Fig. 4. Error bars indicate means ± s.e.m.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–6
Rights and permissions
About this article

Cite this article
Marek, R., Jin, J., Goode, T.D. et al. Hippocampus-driven feed-forward inhibition of the prefrontal cortex mediates relapse of extinguished fear. Nat Neurosci 21, 384–392 (2018). https://doi.org/10.1038/s41593-018-0073-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-018-0073-9
This article is cited by
-
Dorsal hippocampus to nucleus accumbens projections drive reinforcement via activation of accumbal dynorphin neurons
Nature Communications (2024)
-
Dynorphin / kappa-opioid receptor regulation of excitation-inhibition balance toggles afferent control of prefrontal cortical circuits in a pathway-specific manner
Molecular Psychiatry (2023)
-
Central medial thalamic nucleus dynamically participates in acute itch sensation and chronic itch-induced anxiety-like behavior in male mice
Nature Communications (2023)
-
Thalamic nucleus reuniens coordinates prefrontal-hippocampal synchrony to suppress extinguished fear
Nature Communications (2023)
-
A prolonged stress rat model recapitulates some PTSD-like changes in sleep and neuronal connectivity
Communications Biology (2023)
