
Abstract
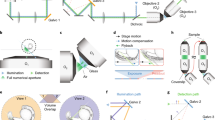
Here we present a fluorescence microscope light path that enables imaging, during free behavior, of thousands of neurons in mice and hundreds of neurons in juvenile songbirds. The light path eliminates traditional illumination optics, allowing for head-mounted microscopes that have both a lower weight and a larger field of view (FOV) than previously possible. Using this light path, we designed two microscopes: one optimized for FOV (~4 mm FOV; 1.4 g), and the other optimized for weight (1.0 mm FOV; 1.0 g).
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others


Data availability
The imaging data used to generate the figures in this paper are freely available at the following repository: https://academictorrents.com/details/aa2a3d2f5ff0b3974871db47db57bc3dabf3c192.
Code availability
The code used to perform the analysis and generate the figures in this paper is freely available under the MIT License at the following repository: https://github.com/FeeLab/Feescope-Paper-Code. The firmware source code along with build instructions is available under the CC BY-NC-SA 4.0 license at the following repository: https://github.com/FeeLab/Fee-Scopes.
References
Ghosh, K. K. et al. Miniaturized integration of a fluorescence microscope. Nat. Methods 8, 871–878 (2011).
Cai, D. J. et al. A shared neural ensemble links distinct contextual memories encoded close in time. Nature 534, 115–118 (2016).
Aghajan, Z. M. et al. Impaired spatial selectivity and intact phase precession in two-dimensional virtual reality. Nat. Neurosci. 18, 121–128 (2015).
Li, Y. et al. Neuronal representation of social information in the medial amygdala of awake behaving mice. Cell 171, 1176–1190.e17 (2017).
Roberts, T. F. et al. Identification of a motor-to-auditory pathway important for vocal learning. Nat. Neurosci. 20, 978–986 (2017).
Chen, K.-S. et al. A hypothalamic switch for REM and Non-REM sleep. Neuron 97, 1168–1176.e4 (2018).
Ziv, Y. et al. Long-term dynamics of CA1 hippocampal place codes. Nat. Neurosci. 16, 264–266 (2013).
Sofroniew, N. J., Flickinger, D., King, J. & Svoboda, K. A large field of view two-photon mesoscope with subcellular resolution for in vivo imaging. eLife 5, e14472 (2016).
Adams, J. K. et al. Single-frame 3D fluorescence microscopy with ultraminiature lensless FlatScope. Sci. Adv. 3, e1701548 (2017).
Xue, Y., Davison, I. G., Boas, D. A. & Tian, L. Single-shot 3D wide-field fluorescence imaging with a computational miniature mesoscope. Sci. Adv. 6, eabb7508 (2020).
Liberti, W. A. III, Perkins, L. N., Leman, D. P. & Gardner, T. J. An open source, wireless capable miniature microscope system. J. Neural Eng. 14, 045001 (2017).
de Groot, A. et al. NINscope, a versatile miniscope for multi-region circuit investigations. eLife 9, e49987 (2020).
Scott, B. B. et al. Imaging cortical dynamics in GCaMP transgenic rats with a head-mounted widefield macroscope. Neuron 100, 1045–1058.e5 (2018).
Rynes, M. L. et al. Miniaturized head-mounted microscope for whole-cortex mesoscale imaging in freely behaving mice. Nat. Methods 18, 417–425 (2021).
Switz, N. A., D’Ambrosio, M. V. & Fletcher, D. A. Low-cost mobile phone microscopy with a reversed mobile phone camera lens. PLoS ONE 9, 1–7 (2014).
Aronov, D., Andalman, A. S. & Fee, M. S. A specialized forebrain circuit for vocal babbling in the juvenile songbird. Science 320, 630–634 (2008).
Inan, H. et al. Fast and statistically robust cell extraction from large-scale neural calcium imaging datasets. Preprint at bioRxiv https://doi.org/10.1101/2021.03.24.436279 (2021).
Glas, A., Hübener, M., Bonhoeffer, T. & Goltstein, P. M. Benchmarking miniaturized microscopy against two-photon calcium imaging using single-cell orientation tuning in mouse visual cortex. PLoS ONE 14, 1–18 (2019).
Ölveczky, B. P., Andalman, A. S. & Fee, M. S. Vocal experimentation in the juvenile songbird requires a basal ganglia circuit. PLoS Biol. 3, e153 (2005).
Pnevmatikakis, E. A. & Giovannucci, A. NoRMCorre: an online algorithm for piecewise rigid motion correction of calcium imaging data. J. Neurosci. Methods 291, 83–94 (2017).
Payne, H. L., Lynch, G. F. & Aronov, D. Neural representations of space in the hippocampus of a food-caching bird. Science 373, 343–348 (2021).
Dimanico, M. M. et al. Aspects of tree shrew consolidated sleep structure resemble human sleep. Commun. Biol. 4, 722 (2021).
Okobi, D. E. Jr., Banerjee, A., Matheson, A. M. M., Phelps, S. M. & Long, M. A. Motor cortical control of vocal interaction in neotropical singing mice. Science 363, 983–988 (2019).
Finkelstein, A. et al. Three-dimensional head-direction coding in the bat brain. Nature 517, 159–164 (2015).
Tohru, I. Imaging lens. US patent 8934179 (2015).
Fee, M. S. & Leonardo, A. Miniature motorized microdrive and commutator system for chronic neural recording in small animals. J. Neurosci. Methods 112, 83–94 (2001).
Lopes, G. et al. Bonsai: an event-based framework for processing and controlling data streams. Front. Neuroinform. 9, 7 (2015).
Chen, T.-W. et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 499, 295–300 (2013).
Okubo, T. S., Mackevicius, E. L. & Fee, M. S. In vivo recording of single-unit activity during singing in zebra finches. Cold Spring Harb. Protoc. 2014, pdb.prot084624 (2014).
Dana, H. et al. High-performance calcium sensors for imaging activity in neuronal populations and microcompartments. Nat. Methods 16, 649–657 (2019).
Acknowledgements
We thank D. Aharoni and J. Newman for many helpful discussions and for their technical expertise. In addition, we thank M. Wilson, J. Newman, J. Voigts, N. Nikbakht, A. Bahle, M. Happ, A. Tanner and J. Kornfeld for comments on the manuscript. We thank W. Guo (MIT) for providing transgenic mice and Q. Xu (MIT) for baseplate designs and for running mice. M.S.F. acknowledges funding through the McKnight Foundation. J.R.S. acknowledges funding through the Harold and Ruth Newman Family Hertz Graduate Fellowship. G.F.L. acknowledges funding through the Simons Foundation (grant 542977ASPI). J.J.Z. acknowledges funding through the NIH (R21 EY028381-01).
Author information
Authors and Affiliations
Contributions
M.S.F. and J.R.S. designed the optics. G.F.L. and J.R.S. designed the circuitry. G.F.L. wrote the firmware. J.R.S. characterized optical performance. G.F.L. performed surgeries and acquired data in birds. J.J.Z. performed surgeries in mice. J.J.Z., G.F.L. and J.R.S. acquired data in mice. G.F.L. and J.R.S. processed the data. J.R.S. performed the decoding analysis. J.R.S., G.F.L. and M.S.F. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
J.R.S., G.F.L. and M.S.F. have an anticipated financial interest as part of a partnership with Open Ephys, a company working to sell open-source tools based on the technology in this paper. This project continues to be open source.
Peer review
Peer review information
Nature Methods thanks Tycho Hoogland and the other, anonymous, reviewers for their contribution to the peer review of this work. Primary Handling editor: Nina Vogt, in collaboration with the Nature Methods team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Simulation and measurement of optical performance.
a, Ray trace simulation of Featherscope lens assembly (above) and point spread function calculated at the image plane (below). The grid spacing is equal to the pixel width. The cross-shaped behavior of the PSF at the edges of the field of view is characteristic of astigmatic aberrations. b, Image taken with the Featherscope of a USAF 1951 resolution target. The line widths of the largest and smallest elements visible in the inset are 3.91 μm and 2.19 μm, respectively. c, measurements of Featherscope focal surface compared to simulations. Bead height at best focus is plotted in blue for the set of beads used in the PSF measurement in Fig. 1. Simulation results for a spherical surface of best focus is plotted in orange. d, Same as (a) but for the Kiloscope lens assembly. e, Same as in (b) but with the Kiloscope. The line widths of the largest and smallest elements visible in the inset are 6.96 μm and 4.38 μm, respectively. f, Same as in (c) but for the Kiloscope.
Extended Data Fig. 2 Illumination coupling optics.
a, optical assembly for adapting NA and mode scrambling the excitation light before coupling it into the microscope. The beam profile is illustrated below at various points in the assembly. Spacing between the axicons was adjusted for optimal microscope performance, and it was noted that the optimal configuration resulted in a central dark spot in the output beam. This spot is not present at the object plane due to imperfections in the fiber. b, Radial profile of the signal collected with the Featherscope while imaging a thin layer of fluorescent dye using different optical configurations. Optical vignetting (orange) illustrates the effects of light losses within the microscope optics and was measured by illuminating the dye with a diffuse light source opposite the objective. Total vignetting (yellow) illustrates the decrease in fluorescent signal caused by the combination of optical vignetting and a Gaussian-shaped excitation light profile. The curve with correction optics (blue) illustrates the flat signal intensity profile that can be achieved with an axicon pair that increases the excitation intensity at the edges of the field of view. c, same as in (b) but for the Kiloscope.
Extended Data Fig. 3 Analog-to-digital converter functionality.
a, Time series recording of two test signals at 100 Hz (orange) and 1000 Hz (blue) using the onboard analog-to-digital converter, sampling at 25.5 kS/s for each signal. b, Power spectral density of the test signals. The spectrum was calculated using Thomson’s multitaper estimate with Slepian tapers. The time series deviates from a pure tone with a root-mean-square residual of 0.2% of the full scale, calculated using the full bandwidth of the recorded signal.
Extended Data Fig. 4 Additional performance characterization of Kiloscope optics.
a, Image taken with the Kiloscope of a fixed coronal brain slice from one Tg(Camk2a-cre)T29-1Stl x Ai95(RCL-GCaMP6f)-D mouse. The image is corrected for vignetting. b, Image of USAF resolution target using Kiloscope optics and a Sony IMX260 image sensor with 1.4 μm pixel width. The smallest element in group 7 is resolvable and has a linewidth of 2.2 μm.
Extended Data Fig. 5 Extraction results from the three birds presented in the paper.
Footprints for each extraction are plotted as colored outlines against a grayscale background of maximum intensity of median filtered data over the course of a recording session. Footprints were extracted using EXTRACT and manually curated based on videos of each footprint’s activity. The high variability in the number of extracted footprints is likely due to variations in virus expression levels between individuals. All scale bars are 100 μm. a, 46 footprints after curation. b, 177 footprints after curation. c, 225 footprints after curation.
Extended Data Fig. 6 Extraction results from the four mice presented in the paper.
Footprints for each extraction are plotted as colored outlines against a grayscale background of the maximum intensity of median filtered data over the course of a recording session. Footprints were extracted using EXTRACT and manually curated for panel (a) based on videos of each footprint’s activity. For panels (b–d), we inspected each footprint by eye and removed visually anomalous footprints and footprints overlapping with blood vessels. The high variability in the number of extracted footprints is likely due to variations in virus expression levels between individuals. All scale bars are 1 mm. a, 4 mm cranial window, 2382 footprints after curation. b, 3 mm cranial window, 1029 footprints. c, 4 mm cranial window, 401 footprints. d, 3 mm cranial window, 112 footprints.
Extended Data Fig. 7 Movement observed using the Kiloscope.
Time series of horizontal and vertical displacement of the FOV, calculated using NoRMCorre, during the recording shown in Fig. 2.
Extended Data Fig. 8 Spatial distribution of coactive neural populations in the Kiloscope dataset shown in Figure 3.
a, Ridgeline plot from Fig. 2d; highlighted in green is a single large correlated event for each NMF component. b, Locations of footprints that participate in each highlighted event. Pixels containing one active footprint are colored blue, two or more overlapping footprints colored red, and inactive footprints colored gray. Footprints are thresholded at the 10% level. Listed in each panel are the number of footprints active in the associated event and the total number of footprints in the NMF component. All scale bars are 750 μm.
Extended Data Fig. 9 Decoding position within the maze with octant resolution.
a, Circular maze with octants labeled; in this case, direction is not coded. b, Decoding matrix for a set of binary SVM models trained to predict the labels in (a) based on the activity in Fig. 2h. White is set at chance level. c, Map of decoder accuracies using the 32 closest footprints less than 270 μm to a given point. Pixels that do not meet a significance threshold (Benjamini-Hockberg false discovery rate = 5%) are plotted in black.
Extended Data Fig. 10 Receptive fields of selected footprints from the Kiloscope.
a, the circular maze that the mouse explored during the recordings in Fig. 2e–h. b, receptive fields plotted for a collection of footprints from Fig. 2e–h. The plotted intensity is the fraction of the time a given footprint was active at a given location. Footprints were selected by eye for large-scale structure in their receptive fields. c, locations of the selected footprints within the FOV. Footprints were selected blindly without knowledge of their location within the FOV. Regions of overlap between footprints are plotted in red. Scale bar is 750 μm.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Scherrer, J.R., Lynch, G.F., Zhang, J.J. et al. An optical design enabling lightweight and large field-of-view head-mounted microscopes. Nat Methods 20, 546–549 (2023). https://doi.org/10.1038/s41592-023-01806-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41592-023-01806-1
This article is cited by
-
Mesoscopic calcium imaging in a head-unrestrained male non-human primate using a lensless microscope
Nature Communications (2024)
-
Large depth-of-field ultra-compact microscope by progressive optimization and deep learning
Nature Communications (2023)
