
Abstract
Older people are particularly susceptible to infectious and neoplastic diseases of the lung and it is unclear how lifelong exposure to environmental pollutants affects respiratory immune function. In an analysis of human lymph nodes (LNs) from 84 organ donors aged 11–93 years, we found a specific age-related decline in lung-associated, but not gut-associated, LN immune function linked to the accumulation of inhaled atmospheric particulate matter. Increasing densities of particulates were found in lung-associated LNs with age, but not in the corresponding gut-associated LNs. Particulates were specifically contained within CD68+CD169− macrophages, which exhibited decreased activation, phagocytic capacity, and altered cytokine production compared with non-particulate-containing macrophages. The structures of B cell follicles and lymphatic drainage were also disrupted in lung-associated LNs with particulates. Our results reveal that the cumulative effects of environmental exposure and age may compromise immune surveillance of the lung via direct effects on immune cell function and lymphoid architecture.
Similar content being viewed by others


Main
The demographics of the world population are rapidly changing, such that it has been projected that individuals 65 years or older will represent over 20% of the population by 20501. As the majority of healthcare costs, morbidity and mortality from diseases are experienced by individuals over 55 years of age2, there is a need to better understand the mechanisms by which aging increases disease susceptibility. In particular, there is a significant and striking increase in both the incidence and severity of diseases of the lung and respiratory tract with age. Older people are at increased risk for lung damage, as well as severe outcomes from infection with respiratory viruses such influenza3 and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), for which mortality from infection in individuals >75 years is more than 80-fold greater than in younger adults4,5,6. Moreover, neoplastic disease of the lung, including small cell lung cancer, mostly affects individuals older than 60 years7.
Senescent changes in the immune system have been implicated in the increased disease burden in older people. With age, immune cells and functional mediators undergo intrinsic alterations leading to decreased adaptive immune responses, increased inflammation8,9 and decreased regulation10, thus impairing anti-pathogen and anti-tumor immunity. However, the mechanisms for the biased decline in respiratory immunity over age are not known. In addition, aging and its effects on the respiratory tract are shaped by prolonged exposure to the environment through inhalation11,12, although the role of environmental insults in age-associated impairments of the immune system is not well understood.
Studies of immunosenescence in humans sample blood as the most accessible site. However, immune responses occur in mucosal and barrier sites of infection and associated lymphoid organs. For responses to respiratory infections, lung-associated lymph nodes (LLNs) are crucial for adaptive immune responses to new and recurring pathogens, as demonstrated in mouse models13,14. Antigens encountered in the lungs enter LLNs via lymphatics where adaptive immune responses are initiated, including T cell priming and interactions with B cells in specialized lymphoid follicles to promote humoral immunity. Age-associated effects on LNs have been identified in mice15 and morphological changes with age have been reported in human LNs16,17; however, analysis of human LN aging and its effect on immune responses and functionality has not been performed. As part of their role in immune surveillance, LNs also filter impurities from tissues through lymphatics18,19, although the impact of this broader role for LNs on human immune responses remains unexplored. Here, we took an anatomical approach to investigate the cellular, structural and functional niches of tissue-draining LNs in samples from human organ donors of different ages, revealing localized, age-associated changes in LLNs due to the accumulation of inhaled atmospheric particulates from environmental pollutants.
Results
Atmospheric particulates accumulate in LLNs
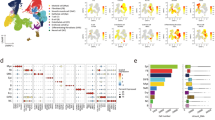
We obtained multiple LN samples associated with lungs and intestines from human organ donors (see Methods), through a human tissue resource we established involving collaborations with organ procurement organizations, as previously described20. Based on our acquisition of tissues from hundreds of organ donors of all ages (with no history of smoking) over the past decade21,22,23,24,25, we consistently observed differences in the appearance of LNs associated with the lung and gut, respectively. Here, we show that LLNs are black in color, while mesenteric LNs (MLNs) from the same individual are beige or translucent in color (Fig. 1a), similar to mouse LNs. Comparing LNs from donors of different ages revealed that black LLNs were observed in the majority of adult donors after the third decade of life but were less prevalent in younger donors (<30 years) (Fig. 1b). Black particulates are present in the atmosphere and consist of different polycyclic aromatic hydrocarbons derived from environmental pollutants such as motor vehicle exhausts, heating and wildfires12,26. We therefore hypothesized that inhaled atmospheric particulates and their accumulation with age may result in specific alterations to the immune function and architecture of LLNs.

a, Photos showing the gross anatomy of LLNs (top) and gut-associated MLNs (bottom) dissected from lung and intestine samples. b, Appearance of whole LLNs by age. c, Graphs of the donors whose samples were used in this study, stratified by age (top) and sex for each site (bottom). d, Brightfield confocal images of human LLNs (left) and MLNs (right). For each site, the left column shows whole LN sections stitched together from 70–400 single 20× images and the right column shows magnified sections, as outlined by the white boxes in the images to the left. The ages of the organ donors are indicated. The black regions show particulate contents. Representative images were taken from 5–7 donors (LLNs) and 4–6 donors (MLNs) per age group (≤39, 40–64 and ≥65 years). Scale bars, 1,500 µm (whole sections except for MLN from 20 yr old donor which is 500 µm) and 100 or 50 µm (single images). e, Particulate content in LNs, quantified by measuring the area (μm2) of each LN that contained black particulates. Imaging data were quantified using Imaris software. Linear regression was performed with Pearson’s correlation (two tailed). P values and Pearson’s R values are provided (**, p<0.01). The data are presented as means ± 95% CIs. The data are from 16–19 donors per site.
We took a quantitative imaging approach to assess the impact of age and inhaled particulates on LN structure and function in LLNs and MLNs from a cohort of 84 organ donors aged 11–93 years, distributed equally between males and females, with no documented history of heavy smoking (>20 packs for 1 year or longer; see Methods for criteria) (Fig. 1c and Extended Data Table 1). We imaged whole LN sections by confocal microscopy, stitching together multiple (70–400) 20× single images (see Methods) to quantify the structural and cellular changes between sites and with age. Brightfield images showed increased particulate matter with age, specifically in LLNs, which was most notable at 40 years of age and older, while MLNs from each corresponding donor did not exhibit particulates at any age (Fig. 1d,e). This age-related accumulation of atmospheric particulates in LLNs was similar between males and females (Extended Data Fig. 1), suggesting environmental effects localized to the LLNs draining the respiratory tract, irrespective of sex.
Uptakes of particulates by CD68+CD169− macrophages in LLNs
LNs comprise densely packed immune cells that are organized in defined structural niches. B cells are organized in follicles that in humans are arranged in the periphery and throughout the organ (Fig. 2a, magenta), while CD8+ T cells are situated around and outside follicles delineating the T cell zone (Fig. 2a, cyan). LN macrophages, as defined in mouse models, are subdivided into different subsets based on their localization within the T cell zone, the subcapsular region or follicles27,28. To define macrophage populations and their localization in human LNs, we stained LN sections with CD68 (a scavenger receptor expressed by tissue or migratory macrophages29) and CD169 (a sialic acid receptor expressed by tissue-resident and subcapsular macrophages in mucosal and lymphoid sites, respectively30,31,32). In human LNs, we observed CD68+ macrophages distributed within the T cell zone and in the subcapsular region around follicles (Fig. 2a, regions 1 and 2), while CD169 expression distinguished subcapsular and medullary from T cell zone macrophages (TCZMs) (Fig. 2a; compare regions 1, 2 and 3). Three subsets of macrophages could be distinguished based on coordinate expression of CD68 and CD169: CD68+CD169− macrophages were mostly found in the T cell zone (Fig. 2a, region 1, green); CD68+CD169+ macrophages were in the subcapsular sinus (region 2, yellow); and CD68−CD169+ macrophages were in the medullary sinus (region 3, red). Human LNs therefore contain macrophages distributed in all regions, with specific subsets in T cell zones and subcapsular regions.

a, Confocal images of LLNs immunostained for CD68 (green), CD169 (red), CD8 (cyan) and CD19 (magenta). Left, whole LN section stitched together from 70–400 single 20× images. Right, magnified sections as outlined by the white boxes to the left. Each row of magnified images shows the localization of a different macrophage subset (1 = CD68+CD169−; 2 = CD68+CD169+; 3 = CD68−CD169+). Representative cells for each subset indicated with yellow arrow. Scale bars, 2,000 µm (whole LN section) and 200 µm (single images). b, Confocal images of human LLNs stained for CD68 and CD169 to show macrophage subsets and black particulate matter localization (brightfield microscopy; white). Yellow arrows indicate the representative CD68+CD169- macrophages containing particulate matter. Representative images were taken from six donors. Scale bars, 50 µm. c, Confocal images of human LLNs stained for CD68 and CD169 to show black particulate matter accumulation in different macrophage subsets of the indicated ages. The left two columns show whole LN sections stitched together from 70–400 single 20× images. The right two columns show magnified sections. Within these sets of images, the left columns show a brightfield imaging overlay to indicate the localization of particulates (white). Yellow arrows indicate the representative CD68+CD169- macrophages containing particulate matter. Representative images were taken from 6–9 donors per age group (≤39, 40–64 and ≥65 years). Scale bars, 1,000 µm (whole LN sections) and 200 µm (single images). d, Bar graph showing the macrophage subsets that contained particulates (<3 µm) for different age groups, as calculated using Imaris software. The data are presented as means ± s.d. and are from 18 donors. Statistical significance was determined by two-way ANOVA with Tukey’s post-hoc test. Statistically significant differences were found for the following comparisons: P = 0.0012 (**, p<0.01) for CD68+CD169− versus CD68+CD169+ at 40–64 years, P = 0.0083 (**) for CD68+CD169− versus CD68−CD169+ at 40–64 years, P < 0.0001(****) for CD68+CD169− versus CD68+CD169+ at ≥65 years and P < 0.0001(****) for CD68+CD169− versus CD68−CD169+ at ≥65 years.
We hypothesized that particulates would be contained within innate immune cells with the capacity to engulf foreign antigens through phagocytosis. In LNs, phagocytic cells mostly comprise macrophages and/or dendritic cells, as neutrophils and monocytes are not significantly represented in LNs21,23. Comparison of brightfield images (particulates) with fluorescence staining of lineage markers in LNs revealed that particulates are contained within macrophages (CD68+ cells) and not within dendritic cells (CD11c+HLA-DR+CD141+ cells) (Fig. 2b and Extended Data Fig. 2a). We further investigated whether particulates were contained within specific macrophages subsets. Using quantitative imaging analysis of whole LNs, we found that the majority of particulates were contained within CD68+CD169− macrophages (Fig. 2c,d), which also expressed CD11c (Extended Data Fig. 2b,c)—a phenotypic feature of TCZMs in mice27,33. We did not find significant particulates in subcapsular (CD68+CD169+) and medullary (CD68−CD169+) macrophages. Together, these results suggest that TCZMs are the predominant subset containing particulate matter within LLNs, consistent with the role of this subset as scavengers for dying and dead cells33.
Particulates impair macrophage phagocytosis and turnover
In tissues, macrophages are strategically localized to serve as a gatekeeper to phagocytose pathogens, dead cells or debris. We investigated whether the increased presence of particulates with age in TCZMs in LLNs would result in altered activation, phagocytic capacity and/or turnover. We used flow cytometry to assess markers of activation and phagocytosis for macrophage subsets in LLNs and MLNs of organ donors aged 18–92 years, to control for the effects of subset, site, age and particulate content. The flow cytometry panel for analysis contained markers for lineage (CD64, CD68 (refs. 29,34), CD11c and CD11b) tissue residence (CD163 (ref. 35), CD169 (refs. 30,31,32) and CX3CR1 (refs. 36,37)), activation (HLA-DR, CD80 and CD86) and phagocytosis, including the scavenger receptor CD36, which is important for the uptake of apoptotic cells and bacteria38, and CD209 (DC-SIGN1), a marker of phagocytic capacity for pathogens39.
Consistent with our imaging data, three major macrophage subsets were delineated based on CD68 and CD169 expression (Fig. 3a and Extended Data Fig. 3 (gating strategy)); each subset differed in the expression of phenotypic markers for macrophage subsets (CD64, CD11c, CD11b, CD163 and CX3CR1), localization (CX3CR1) and function (CD209, HLA-DR, CD36 and CD80/86) (Fig. 3b). We identified differences in the composition of macrophage subsets between LLNs and MLNs; there was an increased frequency of CD68+CD169− macrophages in LLNs compared with MLNs and the frequency of these subsets at each site did not change with age (Fig. 3c,d and Extended Data Fig. 4a). However, expression of the key activation markers CD80 and CD86 and the phagocytic marker CD36 decreased with age specifically in CD68+CD169− macrophages in LLNs but not in MLNs (Fig. 3e), nor in CD68+CD169+ subcapsular and CD68−CD169+ medullary sinus macrophages in either LLNs or MLNs (Fig. 3e and Extended Data Fig. 4b). In contrast, CD209 expression was not altered significantly with age in any macrophage subset at either site (Fig. 3e and Extended Data Fig. 4b). These results show that while the frequencies of LN macrophage subsets are largely maintained as age progresses, the expression of functional markers specifically in the CD68+CD169− subset within LLNs decreases with age, suggesting that particulates may have specific effects on macrophage function.

a, Expression of CD68 and CD169 gated on live, singlet and CD45+CD3− cells, showing CD68+CD169− (black rectangle), CD68−CD169+ (red rectangle) and CD68+CD169+ (blue rectangle) macrophage subsets. b, Heat map showing the differential expression of the indicated markers by the three macrophage subsets, compiled from LLNs. c, Frequency of macrophage subsets in LLNs (n = 21) and MLNs (n = 19). Statistical significance was determined by mixed-effects analysis with Tukey’s post-hoc test. The data are presented as means ± s.d. Statistically significant differences between macrophage subset frequencies in LLNs versus MLNs were found for CD68+CD169− (P=0.0004,*** for p<0.001) and CD68−CD169+(<0.0001,****) cells. There was no significant difference (P = 0.9458) for CD68+CD169+cells. d, Frequency of macrophage subsets in LLNs (n = 21) and MLNs (n = 19) by age. Linear regression was performed with Pearson’s correlation (two tailed). P values and Pearson’s R values are provided (n.s.). The data are presented as means ± 95% CIs. e, Expression of the functional markers CD80/86, CD36 and CD209 by CD68+CD169− and CD68+CD169+ macrophage subsets in LLNs (n = 22) and MLNs (n = 21) by age. Linear regression was performed with Pearson’s correlation (two tailed). P values and Pearson’s R values are provided (*, p<0.05, ***, p<0.001). The data are presented as means ± 95% CIs.
We investigated the direct effect of particulates on phagocytic capacity by imaging and functional assays. The decrease in CD36 expression with age in LLN CD68+CD169− macrophages, as measured by flow cytometry, was also observed by imaging (Fig. 4a), and specifically in CD68+CD169− macrophages containing particulates but not in CD68+CD169− macrophages without particulates (Fig. 4b). To directly assess whether particulate uptake by macrophages inhibits phagocytosis, we performed phagocytic assays using the THP-1 human macrophage cell line exposed to carbon particulates isolated from urban environmental sources (see Methods). THP-1 cells readily take up these atmospheric black particulates within 6 h (Fig. 4c), demonstrating the efficiency of macrophage-mediated surveillance. Using an in vitro phagocytic assay for the uptake of fluorescent bioparticles consisting of bacteria with a pH-sensitive dye that fluoresces when phagocytosed (see Methods), we found a significant decrease in phagocytosis by particulate-containing compared with non-particulate-containing THP-1 cells (Fig. 4d and Extended Data Fig. 5a). These results show that particulate uptake by macrophages can directly impact the phagocytic function of macrophages that are important for scavenging and the maintenance of tissue homeostasis.

a, Confocal images of human LLNs stained for CD68, CD36 and CD169 from donors of the indicated ages. The white arrows show CD68+CD169− macrophages and CD36 expression in LNs from a representative young and older donor. Representative images were taken from 5–6 donors per age group (≤65 and ≥65 years). Scale bars, 15 µm. b, Graphs showing the frequency of CD36+ CD68+CD169− macrophages by age and stratified by those with (left) or without (right) black particulates (n = 11). The calculations were made using Imaris software. Linear regression was performed with Pearson correlation (two tailed). P values and Pearson’s R values are provided (*, p<0.05). The data are presented as means ± 95% CIs. NS, not significant. c, THP-1 macrophages phagocytose black particulates. Differentiated THP-1 cells were incubated with or without black particulates for 6 h. The representative images show black particulates (≥4 μm size) within macrophages (n = 3 wells per condition; three images per well). d, Particulate-containing THP-1 macrophages exhibit decreased phagocytosis. Differentiated THP-1 cells previously incubated with or without black particulates were cultured with pHrodo beads at 37 or 4 °C for 90 min and phagocytic uptake was determined by confocal microscopy (2–3 images per well). The graph shows the frequency of THP-1 cells containing beads for each well and condition, representative of two experiments (n = 9–12 wells per condition). Statistical significance between phagocytosis by THP-1 cells without or with particulates was determined by one-way ANOVA with Tukey’s post-hoc test (****P < 0.0001). e, Differential proliferation of macrophages in LLNs by age. Confocal images of human LLNs stained for CD68, Ki-67 and CD169 are presented. The white arrows show Ki-67+ macrophages. Representative images were taken from 6–7 donors (2–4 images per donor) per age group (≤65 and ≥65 years). Scale bars, 100 μm. f, Graph showing the frequency of proliferating (Ki-67+) cells for each macrophage subset and for different age groups, as calculated using Imaris software. Statistical significance was determined by mixed-effects analysis with Tukey’s post-hoc test. Statistically significant differences were found for the following comparisons: P = 0.004 (**) for CD68+CD169− at ≤39 years versus CD68+CD169− at 40–64 years, P = 0.0095 (**) for CD68+CD169− at ≤39 years versus CD68+CD169− at ≥65 years, P = 0.0373(*) for CD68+CD169– at 40–64 years versus CD68−CD169+ at 40−64 years, P = 0.0023 (**) for CD68+CD169− at ≤39 years versus CD68−CD169+ at ≤39 years, P = 0.0071(**) for CD68−CD169+ at ≤39 years versus CD68+CD169+ at ≤39 years and P = 0.0384 (*) for CD68+CD169+ at ≤39 years versus CD68+CD169+ at 40–64 years. The data are presented as means ± s.d. and are from 13 donors (2–7 single images per donor).
The decrease in phagocytic function and recycling of the membrane suggested that particulates may also affect the ability of tissue macrophages to undergo proliferative turnover for replenishment or maintenance40. We assessed Ki-67 expression in situ as a marker of proliferating cells (Fig. 4e,f). In LN samples from younger donors (≤39 years), the numbers of proliferating CD68+CD169− and CD68+CD169+ macrophages were greater than the number of proliferating CD68−CD169+ macrophages. In contrast, CD68+CD169− macrophages from older donors (≥65 years) had significantly decreased proliferation independent of particulate content (Fig. 4f). CD68+CD169− macrophages with or without particulates did not show a direct correlation in Ki67 expression (Extended Data Fig. 5b). The decreased proliferation of CD68+CD169− macrophage subsets in older individuals (independent of particulates) suggests that particulates do not affect turnover, but rather decreased turnover is a potential mechanism for the accumulation of particulates within macrophages at older ages.
Particulates alter cytokine production by macrophages
Macrophages also elicit critical innate immune functions through the secretion of multiple proinflammatory and regulatory cytokines and mediators41. To define how particulate content affects macrophage-derived cytokine production, we stained LLN sections with antibodies to macrophage-derived factors including the antiviral cytokine interferon-α (IFN-α), the proinflammatory cytokines tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) and the anti-inflammatory mediator arginase42. We previously found that human LLNs exhibit ongoing immune activity compared with other LN sites23, enabling in situ examination of cytokine production. To dissect the individual contributions of particulates and age in macrophage function, we measured cytokine production by CD68+CD169− macrophages with and without particulates in LLNs across all adult ages (Fig. 5 and Extended Data Fig. 6). For each parameter measured, we performed a multivariable regression analysis to control for particulate and age effects (Supplementary Table 1).

a, Confocal images of human LLNs stained for CD68, CD169 and TNF-α from donors of the indicated ages. The arrows show representative CD68+CD169− macrophages (magenta, CD68+CD169− macrophages with particulates; orange, CD68+CD169− macrophages without particulates). The left column shows brightfield imaging overlays (particulates are white). Representative images were taken from 2–5 donors (3–4 images per donor) per age group (≤39, 40–64 and ≥65 years). Scale bars, 20 µm. b, Graph showing the paired frequencies of TNF-α+ CD68+CD169− macrophages with or without black particulates, as calculated using Imaris software. Statistical significance was assessed using repeated measures ANOVA. Statistically significant differences were found for the following comparisons of with versus without particulates: P = 0.0021(**) for ≤39 years, P = 0.0018(**) for 40–64 years and P = 0.0092(**) for ≥65 years. The data are from ten donors (3–4 single images per donor). c, Graphs showing the frequencies of TNF-α+ CD68+CD169− macrophages with or without particulates by age, as calculated using Imaris software (n = 10). Linear regression was performed with Pearson’s correlation (two tailed). P values and Pearson’s R values are provided (*, p<0.05). The data are presented as means ± 95% CIs. d, As in a, but with staining for IFN-α instead of TNF-α. Representative images were taken from 4–6 donors (2–4 images per donor) per age group (≤39, 40–64 and ≥65 years). Scale bars, 20 µm. e, As in b, but for IFN-α+ CD68+CD169− macrophages. The P values for comparisons of with versus without black particulates were P = 0.0021(**) for ≤39 years, P = 0.0036(**) for 40–64 years and P = 0.4170(n.s.) for ≥65 years. The data are from 15 donors (2–4 single images per donor). f, As in c, but for IFN-α+ CD68+CD169− macrophages (n = 15). (**, p < 0.01).
Across ages, macrophages containing particulates exhibited decreased frequencies of cytokine production compared with macrophages without particulates for IFN-α, TNF-α and IL-6 (Fig. 5a–f, Extended Data Fig. 6a–c and Supplementary Table 1). This finding is consistent with particulates having inhibitory effects on macrophage function. In contrast, arginase was produced comparably by macrophages containing or lacking particulates (Extended Data Fig. 6d–f). When comparing functional capacity across age, we found three patterns showing differential effects of particulates and age for various mediators. For TNF-α, there was an age-associated increase in the frequency of TNF-α+ macrophages independent of particulates, although the frequency of TNF-α+ macrophages with particulates was lower at all ages (Fig. 5a–c). Multivariate analysis showed significant yet independent effects of particulates and age for TNF-α (Supplementary Table 1). For IFN-α expression, we found a significant increase with age only in the particulate-containing macrophage subset, and multivariate analysis further revealed independent effects of particulates, with decreased IFN-α expression at younger ages (Fig. 5d–f and Supplementary Table 1). In contrast, both arginase and IL-6 did not show age-associated changes in their expression, although IL-6 expression was decreased in particulate- compared with non-particulate-containing macrophages (Extended Data Fig. 6a–f and Supplementary Table 1). Together, these results show independent and synergistic effects of particulates and age on macrophage function in LLNs.
Particulates disrupt LN architecture and lymphatic drainage
Adaptive immune responses are primed within LN follicles, where T cells interact with B cells to promote the production of antibody-secreting plasma cells and memory B cells43. Disruptions of these follicles due to cytokines, chemokines or infections can lead to a decreased adaptive immune response44,45. In addition, structural connections with lymphatic vessels are important for the transit of immune cells throughout tissues and for migration of dendritic cells from tissues to LNs for T cell priming46. We investigated whether the accumulation of carbon particulates in aging LLNs affects LN architecture and immunosurveillance. We observed that B cell follicles in LLNs became more dispersed with age, resulting in a loss of B cell follicle integrity, as quantified using a computational measure of follicle circularity, while follicles within MLNs did not exhibit significant structural changes with age (Fig. 6a,b). Similarly, an assessment of lymphatics by staining human LN sections with podoplanin47 (Fig. 6c) revealed a slight decrease in the total lymphatics area in both LLNs and MLNs with age, but this did not achieve significance (Fig. 6d). However, in regions containing particulate matter, there were significantly fewer lymphatic vessels in older compared with younger adults (Fig. 6d). The accumulation of carbon particulates is therefore accompanied by a disruption in LN architecture, affecting follicles and lymphatic drainage, with potential impacts on the priming of adaptive immunity and immune surveillance.

a, Representative confocal images of whole human LN sections from donors of the indicated ages, stained for CD20 to show B cell follicles (green) and with brightfield imaging overlays (white) to show black particulate matter. Representative images were taken from 3−7 donors (LLNs) and 2–8 donors (MLNs) per age group (≤39, 40–64 and ≥65 years). Scale bars, 1,000 µm. b, B cell circularity measured using an open-source code (see Methods) to quantify B cell follicle integrity in MLNs (n = 14) and LLNs (n = 17). The graphs show the circularity measurement above a threshold (>0.5 µm) for each whole LN section obtained from organ donors of the indicated ages. Linear regression was performed with Pearson’s correlation (two tailed). P values and Pearson’s R values are provided (** for p<0.01). The data are presented as means ± 95% CIs. c, Representative confocal images of human LLNs (from donors of the indicated ages) stained for podoplanin to show the lymphatics (red; middle). Brightfield imaging overlays (white; left) show the black particulate matter. In the right column, they are pseudocolored green to show the overlay with podoplanin (the particulate matter and lymphatics were isosurfaced using Imaris software). Representative images were taken from 5–12 donors per age group (≤65 and ≥65 years). Scale bars, 1,000 µm. d, Left, graph showing the percentage of the whole LN section covered by lymphatics. Right, graph showing the area covered by lymphatics close to the particulate area (<3 µm), as determined using Imaris software. Statistical significance was determined by one-way ANOVA with Tukey’s post-hoc test. Statistically significant differences were found for the following comparisons of podoplanin+ area (right graph): ≤39 versus 40–64 years (P = 0.0257, *) and ≤39 versus ≥65 years (P = 0.0279, *). The data are presented as means ± s.d. and are from 17 donors.
Discussion
Diseases of the lungs and respiratory tract disproportionately affect older people, as evidenced by the dramatically increased susceptibility to respiratory infections that has been observed during the SARS-CoV-2 pandemic. Here, we reveal a new mechanism for compromised respiratory immunity with increasing age due to exposure to inhaled particulates from the environment that have specific effects on LLNs, which provide critical immunosurveillance functions. We show that particulates are contained within TCZMs in LLNs but are not present in TCZMs in MLNs within the same individual. Particulate-containing macrophages exhibit decreased phagocytosis over age, probably due to the direct effects of particulates, which also decreases cytokine production and can exacerbate age-associated inflammation. Moreover, increased particulate content after the age of 40 years results in a disruption of LN structure and lymphatic connections. Our findings provide evidence for individual and cumulative effects of environmental insults and senescent changes on lung-localized immunity.
Macrophages orchestrate the innate immune response through their phagocytic uptake of pathogens and the production of cytokines for immune cell recruitment and the initiation of adaptive immunity. They also maintain tissue homeostasis through the uptake and elimination of dead cells and debris48,49. Here, we provide direct evidence that macrophages take up atmospheric particulates that lodge in LLNs. Whether these macrophages originate in the lungs is not known, although the increased prevalence of CD68+CD169− macrophages in LLNs versus MLNs suggests recruitment to LLNs from the lungs. Our results also indicate that the fate of particulate-containing macrophages may differ with age. Macrophages in the LNs of younger individuals exhibited higher turnover than those from older individuals, which may facilitate particulate clearance. In this way, young macrophages may be more resilient to the detrimental effects of particulate accumulation.
Particulates also have adverse effects on macrophage function. We showed that a high concentration of particulates can be engulfed by macrophages, which results in impaired phagocytic capacity mediated by direct recognition via scavenger receptors or other non-opsonized pathways. Whether phagocytosis of pathogens and cellular debris or Fc receptor-mediated opsonization are affected remains to be established. We also detected independent effects of particulates on cytokine production by macrophages. Particulate-containing macrophages exhibited lower production of key proinflammatory cytokines including TNF-α, IFN-α and IL-6, which are essential for innate responses to pathogens. We therefore propose a direct effect of particulates in multiple aspects of macrophage function and turnover. The resultant persistence of functionally impaired macrophages in LLNs may contribute to the dysregulated innate responses to respiratory pathogens known to occur in older people5,50.
We also found disrupted follicular structure and decreased lymphatic connections in LLNs with high particulate content, mostly found in individuals older than 50 years. These results suggest impaired priming of adaptive immune responses for newly encountered respiratory pathogens, which is known to occur with age5. However, the ability of particulate-containing LNs to support priming needs to be specifically investigated. Older individuals are intrinsically compromised in their ability to respond to new pathogens due to diminished numbers of naive T cells and a lack of thymic output51. The loss of structural niches for priming in LLNs further exacerbates the impact of these senescent changes on respiratory immunity. In our previous studies, we showed that the proportions of tissue-resident memory T cells and influenza virus-specific tissue-resident memory T cells were maintained over age in human lungs, but decreased in frequency in LLNs24,52,53, suggesting that altered LLN architecture also impacts the maintenance of memory T cells. Together, our findings indicate that age-associated changes to the immune system are local and anatomical, as well as intrinsic to specific cell types.
Pollution from carbon-based sources is an ongoing and growing threat to the health and livelihood of the world’s population54. The specific effects of pollutants on lung inflammation and asthma in certain individuals or within certain geographic regions have been documented55,56,57. Here, through examination of lymphoid tissues, we demonstrate a chronic and ubiquitous impact of pollution on our ability to mount critical immune defense and surveillance of the lungs. In this way, older people are highly vulnerable to pathogens that infect the respiratory tract, as has been tragically demonstrated with the COVID-19 pandemic. The effects of pollutants on neurodegenerative disease are also well documented58,59 and neuroinflammation is implicated in this process60. We therefore propose that policies to limit carbon emissions will not only improve the global climate, but also preserve our immune systems and their ability to protect against current and emerging pathogens and to maintain tissue health and integrity.
In conclusion, our results provide direct evidence that the environment can have cumulative and adverse effects on our immune system with age. We show how environmental pollutants specifically target immune cells within lymphoid organs, which carry out essential immune surveillance functions. These findings can inform how we monitor and study our immune system—in health and disease and with age.
Methods
Human samples
LN tissues were obtained from brain-dead organ donors through an approved protocol and material transfer agreement with LiveOnNY, the local organ procurement organization for the New York metropolitan area, as previously described21,24. Tissues for this study were obtained from donors with no history of asthma and who were negative for SARS-CoV-2, cancer, hepatitis B and C and human immunodeficiency virus. We also selected for donors who were indicated in the donor information sheet as being non-smokers and/or with no history of heavy smoking (>20 packs for >1 year or more) where indicated (82/84 donors). A list of the donors used in this study, along with information on their age and sex, is provided in Extended Data Table 1. This study does not qualify as having performed research on human participants because the tissues were obtained from deceased (brain-dead) organ donors, as confirmed by the Institutional review board at Columbia University.
Preparation of cell suspensions from tissue samples
Following procurement, organs were transported to the laboratory and maintained in cold media supplemented with 5% fetal bovine serum (FBS) and penicillin/streptomycin with glutamine. LN tissues were dissected out from the lungs or intestines, cleaned of fat and connective tissue, chopped into pieces and incubated with RPMI media (Thermo Fisher Scientific) containing collagenase D (Sigma–Aldrich) and DNase (Sigma–Aldrich) for 60 min at 37 °C. Cells were isolated with additional mechanical digestion and density gradient centrifugation, with high yields of live cells, as previously described23,25.
Tissue preparation for confocal imaging
Dissected LN tissues were fixed in paraformaldehyde, lysine and periodate buffer (PLP; 0.05 M phosphate buffer, 0.1 M l-lysine (pH 7.4), 2 mg ml−1 NaIO4 and 10 mg ml−1 paraformaldehyde) overnight at 4 °C (Supplementary Table 3). The following day, tissues were dehydrated in 30% sucrose overnight at 4 °C and subsequently embedded in Optimal Cutting Temperature compound. Donor LNs bearing identification numbers of <410 were fixed in phosphate-buffered saline (PBS) solution with 1% paraformaldehyde and 0.1 M l-lysine, incubated in 20% sucrose at 4 °C and subsequently embedded in Optimal Cutting Temperature compound. Frozen tissues were sectioned using a Leica 3050 S cryostat at 20 μm thickness. Intracellular staining media was prepared with PBS containing 2% goat serum, 2% FBS, 0.05% Tween-20 and 0.3% Triton-X. Tissues were blocked with Human TruStain FcX (BioLegend; 1:100 dilution) in intracellular staining media for 1 h at room temperature. Sections were washed with the intracellular staining buffer and stained with the indicated antibodies (Supplementary Table 2) at 1:25 dilution for 1 h at room temperature. Cytokine staining was performed with their corresponding isotypes to eliminate any signal due to non-specific binding. Images were acquired with a Nikon Eclipse Ti inverted confocal microscope using the dry 20× objective. For fluorescence detection, the following lasers were used: 405, 488,561 and 638 nm. For imaging of whole LN sections, 70–400 20× images were acquired depending on the size of the LN, then computationally stitched using NIS-Elements software (Nikon). Single 20× images were imaged in 2 μm 3× Z-steps. Images were analyzed using Imaris software (Bitplane, Oxford Instruments; version 9.5/9.6), including spot, surface, shortest distance, circularity, isosurfacing and pseudocoloring functions.
Flow cytometry
LN cells were enriched for CD3− cells using biotin-conjugated anti-CD3 and an EasySep Human Biotin Positive Selection Kit II (STEMCELL Technologies; Supplementary Table 3). Following enrichment, LN cells were resuspended in FACS buffer (PBS, 5% FBS and 0.5% sodium azide) and stained for surface markers at a 1:100 dilution for 20 min at 4 °C (Supplementary Table 2). For intracellular staining, cells were resuspended with Fixation/Permeabilization Concentrate and incubated for 60 min at room temperature as indicated in the Tonbo Biosciences Transcription Factor Staining Buffer Kit (TNB-0607-KIT). Cells were washed with permeabilization buffer, resuspended in the permeabilization buffer with antibodies at a 1:100 dilution and incubated for 60 min at room temperature. Cells were then washed and resuspended in FACS buffer, then analyzed by flow cytometry using a BD LSR II cytometer (Becton Dickinson) and the data were analyzed using FlowJo software (Tree Star version 10.7.1). To generate the heat map of marker expression, geometric mean fluorescence intensities for each surface marker were exported from FlowJo, normalized to the average expression of each marker across the subsets and plotted in Python using the seaborn package.
Phagocytosis assay
The human macrophage line THP-1 (kindly provided by S. Ghosh) was maintained in RPMI 1640 medium supplemented with 10% FBS and 10,000 IU penicillin (per ml), 10,000 µg ml−1 streptomycin and 29.2 mg ml−1 l-glutamine (Thermo Fisher Scientific). THP-1 cells (4 × 104) were differentiated using (400 nM) Phorbol 12-myristate 13-acetate (Sigma–Aldrich) for 3 d in 6-well plates and the cells were rested overnight in fresh media (RPMI 1640 medium supplemented with 10% FBS and 10,000 IU penicillin (per ml), 10,000 µg ml−1 streptomycin and 29.2 mg ml−1 l-glutamine). Atmospheric particulates purified from air filters and analyzed by the National Institute of Standards and Technology26 were purchased from Sigma–Aldrich. To test the uptake of particulates, differentiated THP-1 cells were incubated in fresh media with black particulates (0.0006 g ml−1) for 6 h at 37 °C. Following incubation, cells were washed twice to remove all of the free-floating particulates and were rested overnight in fresh media. To assess phagocytosis, differentiated THP-1 cells without particulates (control) and with particulates were cultured in live imaging solution with pHrodo Red E. coli BioParticles, according to the manufacturer‘s protocol, at 4 °C (control condition) or 37 °C (experimental condition) for 90 min. Fresh live imaging solution was added to the cells and they were imaged immediately using confocal microscopy.
Statistical analysis
Statistical analysis was performed using Prism 8.2.0 (GraphPad) with the exception of the multivariable regression analysis, which was done using Excel version 16.54. For correlations with age, Pearson’s correlation was used, and Pearson’s R values and their associated P values are included in the figures. Statistical comparisons between two groups were performed using a Student’s t-test. To assess differences between cytokine production and Ki-67 in macrophages with and without particulates, we used repeated measures two-way analysis of variance (ANOVA). To further assess the differences in cytokine production, multivariable regression analysis was used to control for particulate- and age-related effects. Where noted, three or more groups were analyzed using one-way ANOVA, two-way ANOVA or mixed-effects analysis with Tukey’s post-hoc test. As indicated in the captions, the use of asterisks to indicate significance as determined in Prisms was as follows: ns: p>0.05; * p<0.05; ** p<0.01; ***p<0.001; ****p<0.0001.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Source data are provided with this paper.
Code availability
The Python code used to analyze B cell circularity is available from GitHub (https://github.com/Ironhorse1618/Python3.7-Imaris-XTensions).
References
Schneider, J. L. et al. The aging lung: physiology, disease, and immunity. Cell 184, 1990–2019 (2021).
Why Population Aging Matters: A Global Perspective (National Institute on Aging et al., 2007).
Pop-Vicas, A. & Gravenstein, S. Influenza in the elderly: a mini-review. Gerontology 57, 397–404 (2011).
O’Driscoll, M. et al. Age-specific mortality and immunity patterns of SARS-CoV-2. Nature 590, 140–145 (2021).
Akbar, A. N. & Gilroy, D. W. Aging immunity may exacerbate COVID-19. Science 369, 256–257 (2020).
Metcalf, C. J. E. et al. Comparing the age and sex trajectories of SARS-CoV-2 morbidity and mortality with other respiratory pathogens. R. Soc. Open Sci. 9, 211498 (2022).
Presley, C. J., Reynolds, C. H. & Langer, C. J. Caring for the older population with advanced lung cancer. Am. Soc. Clin. Oncol. Educ. Book 37, 587–596 (2017).
Lopez-Otin, C., Blasco, M. A., Partridge, L., Serrano, M. & Kroemer, G. The hallmarks of aging. Cell 153, 1194–1217 (2013).
Frasca, D. & Blomberg, B. B. Inflammaging decreases adaptive and innate immune responses in mice and humans. Biogerontology 17, 7–19 (2016).
Williams, A. E., Jose, R. J., Brown, J. S. & Chambers, R. C. Enhanced inflammation in aged mice following infection with Streptococcus pneumoniae is associated with decreased IL-10 and augmented chemokine production. Am. J. Physiol. Lung Cell. Mol. Physiol. 308, L539–L549 (2015).
Nilsson Sommar, J. et al. Long-term exposure to particulate air pollution and black carbon in relation to natural and cause-specific mortality: a multicohort study in Sweden. BMJ Open 11, e046040 (2021).
Weisberg, S. P., Ural, B. B. & Farber, D. L. Tissue-specific immunity for a changing world. Cell 184, 1517–1529 (2021).
Legge, K. L. & Braciale, T. J. Lymph node dendritic cells control CD8+ T cell responses through regulated FasL expression. Immunity 23, 649–659 (2005).
Paik, D. H. & Farber, D. L. Influenza infection fortifies local lymph nodes to promote lung-resident heterosubtypic immunity. J. Exp. Med. 218, e20200218 (2021).
Masters, A. R. et al. Assessment of lymph node stromal cells as an underlying factor in age-related immune impairment. J. Gerontol. A Biol. Sci. Med. Sci. 74, 1734–1743 (2019).
Luscieti, P., Hubschmid, T., Cottier, H., Hess, M. W. & Sobin, L. H. Human lymph node morphology as a function of age and site. J. Clin. Pathol. 33, 454–461 (1980).
Hadamitzky, C. et al. Age-dependent histoarchitectural changes in human lymph nodes: an underestimated process with clinical relevance? J. Anat. 216, 556–562 (2010).
Buettner, M. & Bode, U. Lymph node dissection—understanding the immunological function of lymph nodes. Clin. Exp. Immunol. 169, 205–212 (2012).
Davies, M. L. et al. A systemic macrophage response is required to contain a peripheral poxvirus infection. PLoS Pathog. 13, e1006435 (2017).
Farber, D. L. Tissues, not blood, are where immune cells act. Nature 593, 506–509 (2021).
Carpenter, D. J. et al. Human immunology studies using organ donors: impact of clinical variations on immune parameters in tissues and circulation. Am. J. Transpl. 18, 74–88 (2018).
Dogra, P. et al. Tissue determinants of human NK cell development, function, and residence. Cell 180, 749–763 (2020).
Granot, T. et al. Dendritic cells display subset and tissue-specific maturation dynamics over human life. Immunity 46, 504–515 (2017).
Thome, J. J. et al. Spatial map of human T cell compartmentalization and maintenance over decades of life. Cell 159, 814–828 (2014).
Miron, M. et al. Human lymph nodes maintain TCF-1hi memory T cells with high functional potential and clonal diversity throughout life. J. Immunol. 201, 2132–2140 (2018).
Schantz, M. M. et al. Development of two fine particulate matter standard reference materials (<4 μm and <10 μm) for the determination of organic and inorganic constituents. Anal. Bioanal. Chem. 408, 4257–4266 (2016).
Bellomo, A., Gentek, R., Bajenoff, M. & Baratin, M. Lymph node macrophages: scavengers, immune sentinels and trophic effectors. Cell Immunol. 330, 168–174 (2018).
Gray, E. E. & Cyster, J. G. Lymph node macrophages. J. Innate Immun. 4, 424–436 (2012).
Davies, L. C., Jenkins, S. J., Allen, J. E. & Taylor, P. R. Tissue-resident macrophages. Nat. Immunol. 14, 986–995 (2013).
Ural, B. B. et al. Identification of a nerve-associated, lung-resident interstitial macrophage subset with distinct localization and immunoregulatory properties. Sci. Immunol. 5, eaax8756 (2020).
O’Neill, A. S., van den Berg, T. K. & Mullen, G. E. Sialoadhesin—a macrophage-restricted marker of immunoregulation and inflammation. Immunology 138, 198–207 (2013).
Martinez-Pomares, L. & Gordon, S. CD169+ macrophages at the crossroads of antigen presentation. Trends Immunol. 33, 66–70 (2012).
Baratin, M. et al. T cell zone resident macrophages silently dispose of apoptotic cells in the lymph node. Immunity 47, 349–362 (2017).
Tremble, L. F. et al. Differential association of CD68+ and CD163+ macrophages with macrophage enzymes, whole tumour gene expression and overall survival in advanced melanoma. Br. J. Cancer 123, 1553–1561 (2020).
Etzerodt, A. & Moestrup, S. K. CD163 and inflammation: biological, diagnostic, and therapeutic aspects. Antioxid. Redox Signal. 18, 2352–2363 (2013).
Medina-Contreras, O. et al. CX3CR1 regulates intestinal macrophage homeostasis, bacterial translocation, and colitogenic Th17 responses in mice. J. Clin. Invest. 121, 4787–4795 (2011).
Ensan, S. et al. Self-renewing resident arterial macrophages arise from embryonic CX3CR1+ precursors and circulating monocytes immediately after birth. Nat. Immunol. 17, 159–168 (2016).
Erdman, L. K. et al. CD36 and TLR interactions in inflammation and phagocytosis: implications for malaria. J. Immunol. 183, 6452–6459 (2009).
Schulz, D., Severin, Y., Zanotelli, V. R. T. & Bodenmiller, B. In-depth characterization of monocyte-derived macrophages using a mass cytometry-based phagocytosis assay. Sci. Rep. 9, 1925 (2019).
Hashimoto, D. et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity 38, 792–804 (2013).
Cox, N., Pokrovskii, M., Vicario, R. & Geissmann, F. Origins, biology, and diseases of tissue macrophages. Annu. Rev. Immunol. 39, 313–344 (2021).
Sica, A. & Mantovani, A. Macrophage plasticity and polarization: in vivo veritas. J. Clin. Invest. 122, 787–795 (2012).
Huang, Q., Xu, L. & Ye, L. T cell immune response within B-cell follicles. Adv. Immunol. 144, 155–171 (2019).
Popescu, M., Cabrera-Martinez, B. & Winslow, G. M. TNF-α contributes to lymphoid tissue disorganization and germinal center B cell suppression during intracellular bacterial infection. J. Immunol. 203, 2415–2424 (2019).
Kaneko, N. et al. Loss of Bcl-6-expressing T follicular helper cells and germinal centers in COVID-19. Cell 183, 143–157 (2020).
Petrova, T. V. & Koh, G. Y. Biological functions of lymphatic vessels. Science 369, eaax4063 (2020).
Schacht, V. et al. T1α/podoplanin deficiency disrupts normal lymphatic vasculature formation and causes lymphedema. EMBO J. 22, 3546–3556 (2003).
Arandjelovic, S. & Ravichandran, K. S. Phagocytosis of apoptotic cells in homeostasis. Nat. Immunol. 16, 907–917 (2015).
Kurosaka, K., Takahashi, M., Watanabe, N. & Kobayashi, Y. Silent cleanup of very early apoptotic cells by macrophages. J. Immunol. 171, 4672–4679 (2003).
Boe, D. M., Boule, L. A. & Kovacs, E. J. Innate immune responses in the ageing lung. Clin. Exp. Immunol. 187, 16–25 (2017).
Nikolich-Zugich, J. Aging of the T cell compartment in mice and humans: from no naive expectations to foggy memories. J. Immunol. 193, 2622–2629 (2014).
Kumar, B. V., Connors, T. J. & Farber, D. L. Human T cell development, localization, and function throughout life. Immunity 48, 202–213 (2018).
Poon, M. M. L. et al. Heterogeneity of human anti-viral immunity shaped by virus, tissue, age, and sex. Cell Rep. 37, 110071 (2021).
Cohen, A. J. et al. Estimates and 25-year trends of the global burden of disease attributable to ambient air pollution: an analysis of data from the Global Burden of Diseases Study 2015. Lancet 389, 1907–1918 (2017).
Matthews, N. C. et al. Urban particulate matter-activated human dendritic cells induce the expansion of potent inflammatory Th1, Th2, and Th17 effector cells. Am. J. Respir. Cell Mol. Biol. 54, 250–262 (2016).
Brandt, E. B. et al. Exposure to allergen and diesel exhaust particles potentiates secondary allergen-specific memory responses, promoting asthma susceptibility. J. Allergy Clin. Immunol. 136, 295–303 (2015).
Guarnieri, M. & Balmes, J. R. Outdoor air pollution and asthma. Lancet 383, 1581–1592 (2014).
Haghani, A., Morgan, T. E., Forman, H. J. & Finch, C. E. Air pollution neurotoxicity in the adult brain: emerging concepts from experimental findings. J. Alzheimers Dis. 76, 773–797 (2020).
Haghani, A., Thorwald, M., Morgan, T. E. & Finch, C. E. The APOE gene cluster responds to air pollution factors in mice with coordinated expression of genes that differs by age in humans. Alzheimers Dement. 17, 175–190 (2021).
Scieszka, D. et al. Neuroinflammatory and neurometabolomic consequences from inhaled wildfire smoke-derived particulate matter in the Western United States. Toxicol. Sci. 186, 149–162 (2022).
Acknowledgements
This work was supported by National Institutes of Health (NIH) grants HL145547, AI106697 and AI128949 awarded to D.L.F. B.B.U. was supported by NIH T32HL105323. P.D. was supported by a Cancer Research Institute Irvington Postdoctoral Fellowship. N.L. is supported by the National Science Foundation Graduate Research Fellowship Program. P.A.S. is supported by the Canadian Institutes of Health Research Fellowship. This study also used the Confocal and Specialized Microscopy Shared Resource core supported by NIH P30 CA013696 and the Columbia Center for Translational Immunology Flow Cytometry Core supported in part by NIH S10RR027050. We thank M. J. Gastinger (Imaris) for the Python code used in the B cell imaging analysis, A. Urso for previous help with confocal microscopy, S. Ghosh for the THP-1 cell line, and the transplant coordinators at LiveOnNY and the donor families for making this study possible.
Author information
Authors and Affiliations
Contributions
B.B.U. designed the experiments, processed the tissues, performed the imaging and flow cytometry, analyzed the data, produced the figures and wrote the paper. P.D., S.B.W., D.P.C. and P.A.S. helped with data analysis and visualization. T.G. made the initial observations of LN particulates and generated the whole-tissue photos. T.S. prepared the tissue samples. P.D., S.B.W., D.P.C., P.A.S., M.M.L.P., N.L., P.T. and Y.S.L. assisted with the tissue processing. T.S., M.K. and R.M. obtained the donor tissues. D.L.F. planned the experiments, coordinated the tissue acquisition and data acquisition/analysis, analyzed the data and wrote and edited the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Medicine thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editors: Saheli Sadanand and Joao Monteiro, in collaboration with the Nature Medicine team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Carbon particulate content in different lymph nodes is similar between males and females.
Carbon particulate content in the lung-associated lymph node (LLN) and mesenteric lymph node (MLN) of donors from Fig. 1 stratified by sex. Data from 16–19 donors per site. Significance calculated by student T-test (two-tailed). Data presented as the mean ± SEM. N.S.: not significant by student’s t-test.
Extended Data Fig. 2 Particulate uptake occurs within CD11c+ macrophages but not dendritic cells (DC).
a. Confocal image of human LLNs stained for MHCII (HLA-DR), CD11c, and CD141 expression to identify DC (CD11c+HLA-DR+CD141+) shown with overlay of brightfield (left columns) for visualizing particulates (middle and right columns). Representative images were taken from 5 donors. Scale bar: 50 um. b. Confocal image of human LLNs stained for macrophage markers CD68 and CD169 along with CD11c. The image on the left show with overlay of brightfield (left columns) to show localization of particulates (white). The image on the right shows the CD68 and CD11c expression on macrophages. Representative images were taken from 6–9 donors per age group (≤39,40–64 and ≥65 yrs). Scale bar: 50–100 um. c. Graphs shows particulate content in specific macrophage subsets for different age groups, quantitated using Imaris software. Significance calculated by 2-way ANOVA with Tukey’s posttest, *P < 0.05, ****P < 0.0001. Significance for 40–64: CD68+CD169−CD11c+ vs. CD68+CD169+CD11c+ P = 0.0144, CD68+CD169−CD11c+ vs. CD68−CD169+CD11c+ P = 0.0316; 65 ≤ : CD68+CD169−CD11c+ vs. CD68+CD169+CD11c+ P = < 0.0001, CD68+CD169−CD11c+ vs. CD68−CD169+CD11c+ P = < 0.0001. Data presented as the mean ± SD. Data are from 17 donors.
Extended Data Fig. 3 Gating strategy.
Flow cytometry gating for CD68+CD169−, CD68−CD169+, and CD68+CD169+ macrophage subsets following gating on live, singlet, and CD45+ cells.
Extended Data Fig. 4 CD68−CD169+ macrophages in LN do not exhibit alterations in functional marker expression with age.
a. Frequency of CD68−CD169+ macrophages in LLN (n = 21) and MLN (n = 19) over age. Linear regression with Pearson correlation (two-tailed). Pearson r, *P < 0.033, **P < 0.005 and ***P < 0.001. Data presented as the mean ± 95% CI. b. Expression of functional markers CD80/86, CD36, and CD209 expression on CD68-CD169+ macrophages over age by flow cytometric analysis of LLNs (n = 21, left column) and MLNs (n = 19, right column). Linear regression with Pearson correlation (two-tailed). Pearson r, *P < 0.033. Data presented as the mean ± 95% CI.
Extended Data Fig. 5 Particulates inhibit phagocytosis but do not alter age-associated effects on proliferation.
a. Particulate containing THP-1 macrophages cannot phagocytose fluorescent pHrodo beads. Representative images of differentiated THP-1 cells previously incubated with (+) or without (-) black particulates subsequently cultured with pHrodo beads at 37 °C or 4 °C for 90 minutes. Confocal image of THP-1 cells imaged for pHrodo beads and brightfield. Yellow arrows indicate THP-1 cells. Scale bar: 50um. Image is representative of two experiments. (n = 9−12 wells/condition). b. Imaging data from Fig. 3g analyzed for Ki67+ CD68+CD169− macrophages that are within or outside the black particulate area. Significance assessed with paired T-test (two-tailed). Data are from 13 donors (2–7 single images for each donor).
Extended Data Fig. 6 Particulate-containing CD68+CD169− macrophages induce an immunoregulatory environment in the LLNs.
a. Confocal image of human LLNs stained for CD68, CD169, and Arginase-1 from donors of indicated ages. Arrows show representative CD68+CD169− macrophages with particulates (magenta) or without particulates (orange). Representative images were taken from 2–6 donors (2–4 images/donor) per age group (≤39,40–64 and ≥65 yrs). Scale bar: 20 um. b. Graph shows paired frequencies of Arg-1+CD68+CD169− macrophages with or without particulates calculated using Imaris software. Significance assessed with repeated measures ANOVA. Significance for ≤39 yrs P = 0.059, 40–64 years P = 0.0088, ≥65 years P = 0.9969. Data are from 13 donors (2–4 single images for each donor). c. Graph shows frequency of Arg-1+CD68+CD169− macrophages with or without particulates over age calculated using Imaris software. (n = 13). Linear regression with Pearson correlation (two-tailed). Pearson r, n.s. Data presented as the mean ± 95% CI. d. Confocal image of human LLNs stained for CD68, CD169, and IL-6 from donors of indicated ages. Arrows show representative CD68+CD169− macrophages with particulates (magenta) or without particulates (orange). Representative images were taken from 4–5 donors (2–4 images/donor) per age group (≤39,40–64 and ≥65 yrs). Scale bar: 20 um. e. Graph shows paired frequencies of IL-6+CD68+CD169− macrophages with or without particulates calculated using Imaris software. Significance assessed with repeated measures ANOVA. Significance for ≤39 yrs P = < 0.0001, 40–64 years P = 0.0037, ≥65 years P = 0.0002. Data are from 14 donors (2–4 single images for each donor) f. Graph shows frequency IL-6+CD68+CD169− macrophages with or without particulates over age calculated using Imaris software. (n = 14). Linear regression with Pearson correlation (two-tailed). Pearson r, n.s. Data presented as the mean ± 95% CI.
Supplementary information
Supplementary Information
Supplementary Tables 1–3.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ural, B.B., Caron, D.P., Dogra, P. et al. Inhaled particulate accumulation with age impairs immune function and architecture in human lung lymph nodes. Nat Med 28, 2622–2632 (2022). https://doi.org/10.1038/s41591-022-02073-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41591-022-02073-x
This article is cited by
-
Pollution impairs lung defences with age
Nature Reviews Immunology (2023)
