
Abstract
It is increasingly recognized that immune development within mucosal tissues is under the control of environmental factors during early life. However, the cellular mechanisms that underlie such temporally and regionally restrictive governance of these processes are unclear. Here, we uncover an extrathymic pathway of immune development within the colon that is controlled by embryonic but not bone marrow–derived macrophages, which determines the ability of these organs to receive invariant natural killer T (iNKT) cells and allow them to establish local residency. Consequently, early-life perturbations of fetal-derived macrophages result in persistent decreases of mucosal iNKT cells and is associated with later-life susceptibility or resistance to iNKT cell–associated mucosal disorders. These studies uncover a host developmental program orchestrated by ontogenically distinct macrophages that is regulated by microbiota, and they reveal an important postnatal function of macrophages that emerge in fetal life.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request. Raw FASTQ files and processed reads of the transcriptional analyses can be accessed at the Gene Expression Omnibus under accession no. GSE167975.
References
Gensollen, T., Iyer, S. S., Kasper, D. L. & Blumberg, R. S. How colonization by microbiota in early life shapes the immune system. Science 352, 539–544 (2016).
Schuijs, M. J. et al. Farm dust and endotoxin protect against allergy through A20 induction in lung epithelial cells. Science 349, 1106–1110 (2015).
Shaw, S. Y., Blanchard, J. F. & Bernstein, C. N. Association between the use of antibiotics in the first year of life and pediatric inflammatory bowel disease. Am. J. Gastroenterol. 105, 2687–2692 (2010).
Cahenzli, J., Köller, Y., Wyss, M., Geuking, M. B. & McCoy, K. D. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe 14, 559–570 (2013).
Elahi, S. et al. Immunosuppressive CD71+ erythroid cells compromise neonatal host defence against infection. Nature 504, 158–162 (2013).
Gollwitzer, E. S. et al. Lung microbiota promotes tolerance to allergens in neonates via PD-L1. Nat. Med. 20, 642–647 (2014).
Olszak, T. et al. Microbial exposure during early life has persistent effects on natural killer T cell function. Science 336, 489–493 (2012).
An, D. et al. Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell 156, 123–133 (2014).
Nieuwenhuis, E. E. S. et al. Cd1d-dependent regulation of bacterial colonization in the intestine of mice. J. Clin. Invest. 119, 1241–1250 (2009).
Heller, F., Fuss, I. J., Nieuwenhuis, E. E., Blumberg, R. S. & Strober, W. Oxazolone colitis, a Th2 colitis model resembling ulcerative colitis, is mediated by IL-13-producing NK-T cells. Immunity 17, 629–638 (2002).
Ginhoux, F. & Guilliams, M. Tissue-resident macrophage ontogeny and homeostasis. Immunity 44, 439–449 (2016).
Bain, C. C. et al. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 15, 929–937 (2014).
Tamoutounour, S. et al. Origins and functional specialization of macrophages and of conventional and monocyte-derived dendritic cells in mouse skin. Immunity 39, 925–938 (2013).
Thomas, S. Y. et al. PLZF induces an intravascular surveillance program mediated by long-lived LFA-1-ICAM-1 interactions. J. Exp. Med. 208, 1179–1188 (2011).
Lynch, L. et al. Regulatory iNKT cells lack expression of the transcription factor PLZF and control the homeostasis of Treg cells and macrophages in adipose tissue. Nat. Immunol. 16, 85–95 (2015).
Pellicci, D. G. et al. A natural killer T (NKT) cell developmental pathway involving a thymus-dependent NK1.1−CD4+ CD1d-dependent precursor stage. J. Exp. Med. 195, 835–844 (2002).
Schreiber, H. A. et al. Intestinal monocytes and macrophages are required for T cell polarization in response to Citrobacter rodentium. J. Exp. Med. 210, 2025–2039 (2013).
Sudo, T. et al. Functional hierarchy of c-kit and c-fms in intramarrow production of CFU-M. Oncogene 11, 2469–2476 (1995).
Squarzoni, P. et al. Microglia modulate wiring of the embryonic forebrain. Cell Rep. 8, 1271–1279 (2014).
Serbina, N. V. & Pamer, E. G. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat. Immunol. 7, 311–317 (2006).
Rantakari, P. et al. Fetal liver endothelium regulates the seeding of tissue-resident macrophages. Nature 538, 392–396 (2016).
Johnston, B., Kim, C. H., Soler, D., Emoto, M. & Butcher, E. C. Differential chemokine responses and homing patterns of murine TCRαβ NKT cell subsets. J. Immunol. 171, 2960–2969 (2003).
Chakravarti, S. et al. Lumican regulates collagen fibril assembly: skin fragility and corneal opacity in the absence of lumican. J. Cell Biol. 141, 1277–1286 (1998).
Bleul, C. C., Fuhlbrigge, R. C., Casasnovas, J. M., Aiuti, A. & Springer, T. A. A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1). J. Exp. Med. 184, 1101–1109 (1996).
Haimon, Z. et al. Re-evaluating microglia expression profiles using RiboTag and cell isolation strategies. Nat. Immunol. 19, 636–644 (2018).
Boisset, J.-C. et al. Mapping the physical network of cellular interactions. Nat. Methods 15, 547–553 (2018).
Olszak, T. et al. Protective mucosal immunity mediated by epithelial CD1d and IL-10. Nature 509, 497–502 (2014).
Constantinides, M. G. & Bendelac, A. Transcriptional regulation of the NKT cell lineage. Curr. Opin. Immunol. 25, 161–167 (2013).
Iyer, S. S. et al. Dietary and microbial oxazoles induce intestinal inflammation by modulating aryl hydrocarbon receptor responses. Cell 173, 1123–1134.e11 (2018).
Boirivant, M., Fuss, I. J., Chu, A. & Strober, W. Oxazolone colitis: a murine model of T helper cell type 2 colitis treatable with antibodies to interleukin 4. J. Exp. Med. 188, 1929–1939 (1998).
Arrunategui-Correa, V. & Kim, H. S. The role of CD1d in the immune response against Listeria infection. Cell. Immunol. 227, 109–120 (2004).
Kamran, P. et al. Parabiosis in mice: a detailed protocol. J. Vis. Exp. 80, 50556 (2013).
Tamoutounour, S. et al. CD64 distinguishes macrophages from dendritic cells in the gut and reveals the Th1-inducing role of mesenteric lymph node macrophages during colitis. Eur. J. Immunol. 42, 3150–3166 (2012).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq: a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Zhou, Y. et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 10, 1523 (2019).
Zhang, T. et al. Deciphering the landscape of host barriers to Listeria monocytogenes infection. Proc. Natl Acad. Sci. USA 114, 6334–6339 (2017).
Wollert, T. et al. Extending the host range of Listeria monocytogenes by rational protein design. Cell 129, 891–902 (2007).
Acknowledgements
R.S.B. is supported by National Institutes of Health grant nos. DK044319, DK053056, DK051362, DK088199 and 5P01AI073748 and the Harvard Digestive Diseases Center (no. P30DK034854). T.G. is supported by the Crohn’s and Colitis Foundation of America Research Fellow Award (no. 418509). We thank Blumberg laboratory members for their assistance in manuscript preparation. We thank S. S. Iyer for organizing the RNA sequencing at the core facility and for helping with the oxazolone colitis. We thank H. Gerke for her assistance in performing the Plvap−/− experiments and R. Baron for assistance with the isolation of lung cells.
Author information
Authors and Affiliations
Contributions
T.G. and R.S.B. conceived, designed and interpreted the experiments. T.G., T.Z. and M.P. carried out the experiments. M.S. and P.R. provided the Plvap−/− mice; the Plvap−/− mice experiments were performed in their laboratories. T.G. and X.L. performed the transcriptional analyses. J.N.G., X.L., F.G., T.Z. and M.W. aided with the interpretation of the data. F.G. and P.S. provided the AFS98 antibodies. T.G. and R.S.B. wrote the manuscript. All authors were involved in the critical revision of the manuscript for important intellectual content.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Immunology thanks Gerard Eberl and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available. Zoltan Fehervari was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1
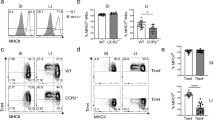
a, Circulatory exchange of CD45.1 (black) or CD45.2 (grey) TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) and iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) cells in the blood of surgically joined CD45.1 (left) and CD45.2 (right) congenic animals (n = 2) determine by flow cytometry, 3 weeks after surgery. Circles are representative of average cell frequency. b, Representative plot of the circulatory exchange of CD45.1 or CD45.2 TCR-αβ+ T and iNKT cells in the colon of surgically joined congenic animals (n = 2) 3 weeks after surgery. c, Schematic of adoptive transfer strategy. d, Adoptive transfer of CD45.1 adult thymic cells into a 4 day old CD45.2 host (n = 1) followed by quantitative analyses of colonic CD45.1 or CD45.2 TCR-αβ+ T and iNKT cells by flow cytometry on day 42.
Extended Data Fig. 2
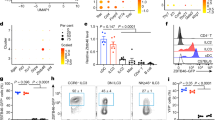
Diphtheria toxin (DT) administered every two days from day 8 to 14 (DT8-14) after birth followed by quantitative analyses on day 15 (H15) of the absolute count of macrophages (CD45+ Lin−F4/80+ CD64+) in the skin (LysCre+/-: n = 3, MMDTR n = 4) (a) or spleen (LysCre+/-: n = 5, MMDTR n = 5) (b) and the absolute count of iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) and TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) cells in the skin (LysCre+/-: n = 8, MMDTR n = 7) (c) of control littermates LysCre+/- or MMDTR animals. DT administered from day 8 to 10 (DT8-10) after birth followed by quantitative analyses on day 11 (H11) of the absolute count of iNKT and TCR-αβ+ T cells in the small intestine (d) and lung (e) of control littermates LysCre+/- (n = 3) or MMDTR (n = 12) animals. DT administered from day 8 to 10 (DT8-10) after birth followed by quantitative analyses on day 11 (H11) of the absolute count of splenic macrophages (f) of control littermates LysCre+/- (n = 5) or MMDTR (n = 3) animals. DT administered from day 12 to 14 (DT12-14) after birth followed by quantitative analyses on day 15 (H15) of the absolute count of splenic macrophages (g) of control littermates LysCre+/- (n = 9) or MMDTR (n = 10) animals. Absolute counts were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Extended Data Fig. 3
Diphtheria toxin (DT) administered every two days from day 8 to 14 after birth followed by quantitative analyses on day 15 of the absolute count of B cells (CD45+ CD19+, LysCre+/-: n = 5, MMDTR n = 5), neutrophils (CD45+ Ly6G+, LysCre+/-: n = 5, MMDTR n = 4), dendritic cells (DC) (CD45+, Lin−, CD11chi, MHCII+, CD64−, LysCre+/-: n = 5, MMDTR n = 4), eosinophils (CD45 + , SiglecF+, LysCre+/-: n = 5, MMDTR n = 4) in the colon of control littermates LysCre+/- or MMDTR animals (a). DT administered every two days from day 8 to 14 after birth followed by quantitative analyses on day 42 of the absolute count of MAIT cells (CD45+ CD3ε+ TCRβ+ MR1/5-OP-RU Tetramer+) in the colon of control littermates LysCre+/- (n = 5) or MMDTR (n = 5) animals (b). Absolute count of macrophages (CD45+ Lin− F4/80+ CD64+) (c), iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) and TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) cells (d) in the colon in the absence of DT treatment in control littermates LysCre+/- (n = 4) and MMDTR (n = 4) animals at 2 weeks old. Absolute counts were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. ns: not-significant.
Extended Data Fig. 4
Diphtheria toxin (DT) administered from day 8 to 10 (DT8-10) after birth followed by quantitative analyses on day 11 (H11) of the absolute count of macrophages (CD45+ Lin− F4/80+ CD64+) in the colon (a) or spleen (b) and the absolute count of iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) and TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) cells in the colon (c) and spleen (d) of control littermates Cx3cr1+/- (n = 4) or Cx3cr1DTR (n = 7) mice. e) Schematic of macrophage depletion model with AFS98 antibody. AFS98 or Isotype control antibody administered from day 4 to 10 (AFS4-10) after birth followed by quantitative analyses on day 11 (H11) of the absolute count of macrophages (n = 6 per group) (f), and the absolute count of iNKT and TCR-αβ+ T cells (n = 12 per group) (g) in the colon of injected animals. Absolute counts were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not-significant.
Extended Data Fig. 5
a, Schematic of macrophage depletion model. Diphtheria toxin (DT) administered every two days from day 56 to 62 (DT Adult) after birth followed by quantitative analyses at day 63 of the absolute count of macrophages (CD45+ Lin− F4/80+ CD64+) in the colon (b) and spleen (c), and the absolute count of iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) and TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) cells in the colon (d) and spleen (e) of control littermates LysCre+/- (colon n = 10, spleen n = 5) or MMDTR (colon n = 10, spleen n = 5) animals. f, Schematic of macrophage depletion model. DT administered from day 15 to 21 (DT15-21) after birth followed by quantitative analyses on day 22 (H22) of the absolute count of macrophages (g) and the absolute count of iNKT and TCR-αβ+ T cells in the colon (h) of control littermates LysCre+/- (n = 8) or MMDTR (n = 6) animals. Absolute counts were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. ***P < 0.001, ****P < 0.0001, ns: not-significant.
Extended Data Fig. 6
a, Absolute count of iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) and TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) cells in the colon of wild type (WT, n = 5) littermates or Ccr2-/- (n = 5) animals at day 56 after birth. b, Diphtheria Toxin (DT) administered at day 1 and (DT1-2) after birth followed by quantitative analyses on day 10 (H10) of the absolute count of F4/80hi/CD11blo and F4/80lo/CD11bhi macrophages (CD45+ Lin− F4/80+ CD64+) in the colon of control littermates LysCre+/- (n = 8) or MMDTR (n = 4) animals. c, Absolute count of iNKT and TCR-αβ+ T cells in the colon of germ-free (GF, n = 12) and GF conventionalized with specific pathogen free (SPF) microbiota prior to birth (GFCV, n = 10) animals at 35 days of life. d, Representative plot of F4/80hi/CD11blo and F4/80lo/CD11bhi macrophages in the colon of SPF, GF or GFCV animals at 15 days old. e, DT administered from day 5 to 7 after birth followed by the analysis of Cxcl16, Cd1d, Cxcl12, Lum, Dcn, Mfap5, Angptl1 and Col6a2 transcript expression by quantitative polymerase chain reaction in the colon of control littermates LysCre+/- (n = 3) or MMDTR (n = 4) animals. Numbers in the representative plots indicate cell frequency. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. *P < 0.05, **P < 0.01, ****P < 0.0001. ns: not-significant.
Extended Data Fig. 7
a, Percentage of Ki67 positive TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) and iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) cells on day 8 (H8) in the colon of control littermates LysCre+/- (n = 8) or MMDTR (n = 8) animals treated with diphtheria toxin (DT) from day 5 to 7 (DT5-7) after birth. b, DT administered from day 56 to 62 (DT Adult) after birth followed by analyses at day 63 of the Ki67 mean fluorescent intensity (MFI) of TCR-αβ+ T and iNKT cells in the spleen of control littermates LysCre+/- (n = 8) or MMDTR (n = 8) animals. c) Percentage of Ki67 positive TCR-αβ+ T and iNKT cells on day 63 in the colon of control littermates LysCre+/- (n = 8) or MMDTR (n = 8) animals treated DT from day 56 to 62 after birth. Representative plot of TCR-αβ+ T and iNKT cells from 11 day old CD45.2 control littermates LysCre+/- or MMDTR animals adoptively transferred with CD45.1 adult thymic cells at 4 days old and treated with DT from day 8 to 10 (DT8-10) after birth in the colon (d) or spleen (e). f, Schematic of adoptive transfer and macrophage depletion model. g, Adoptive transfer of CD45.1 adult thymic cells into 3 day old CD45.2 control littermates LysCre+/- (n = 3) or MMDTR (n = 5) animals followed by DT administration from day 3 to 7 (DT3-7) after birth and quantitative analyses on day 8 (H8) of the Ki67 MFI (left) and percentage (right) of CD45.1 expressing TCR-αβ+ T and iNKT cells in the colon. H) Representative plot (left) and cell percentage (right) of iNKT cell subsets (NKT1, NKT2, NKT 17) from 8 day old CD45.2 control littermates LysCre+/- (n = 3) or MMDTR (n = 5) animals adoptively transferred with CD45.1 adult thymic cells at 3 days old and treated with DT from day 3 to 7 (DT3-7) after birth in the colon. SSC-A, side scatter. Numbers in the representative plots indicate cell frequency and were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. *P < 0.05, **P < 0.01, ****P < 0.0001, ns: not-significant.
Extended Data Fig. 8
a, Schematic of macrophage depletion model ex vivo. Adult spleen from control littermates LysCre+/- or MMDTR animals, digested and cultured for 48 hours with Diphtheria toxin (DT) followed by quantitative analyses of the absolute count of macrophages (CD45+ Lin− F4/80+ CD64+, LysCre+/-: n = 4, MMDTR n = 5) (b), and the absolute count of iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) and TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) (LysCre+/-: n = 5, MMDTR n = 5) cells (c). d, Representative plot of macrophages on day 49 in the colon of control littermates LysCre+/- (n = 4) or MMDTR (n = 4) animals treated with DT from day 8 to 14 after birth. DT administered from day 8 to 14 (DT8-14) after birth followed by quantitative analyses on day 49 (H49) of the absolute count of macrophages (e), and the absolute count of iNKT and TCR-αβ+ T cells in the skin (F) of control littermates LysCre+/- or MMDTR animals. Absolute counts were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. *P < 0.05, ***P < 0.001, ns: not-significant. Representative plot of TCR-αβ+ T and iNKT cells on day 49 in the colon of control littermates LysCre+/- or MMDTR animals treated with DT from day 8 to 14 after birth in the colon (g) and spleen (h). Unl, Unloaded. Tet, Tetramer. Numbers in the representative plots indicate cell frequency.
Extended Data Fig. 9
Representative plot (left) and cell percentage (right) of iNKT cell subsets (NKT1, NKT2, NKT 17) from 42 day old control littermates LysCre+/- (n = 6) or MMDTR (n = 4) animals treated with DT from day 8 to 14 (DT8-14) after birth in the colon (a) or spleen (b). Mean fluorescent intensity (MFI) of Ki67, percentage of Ki67+ and CD69+ TCR-αβ+ T (CD45+ CD3ε+ TCRβ+) and iNKT (CD45+ CD3ε+ TCRβ+ CD1d Tetramer+) cells on day 42 (H42) in the colon (c,d,g) or spleen (E,F,H) of control littermates LysCre+/- (n = 6) or MMDTR (n = 4) animals treated with diphtheria toxin (DT) from day 8 to 14 after birth. I) Schematic of macrophage depletion model and αGalactosylceramide (αGal) treatment. DT administered from day 8 to 14 after birth followed by αGal regimen on day 49 and quantitative analyses 16 hours after, of the IFNγ protein level in the colon of control littermates LysCre+/- (n = 10) or MMDTR (n = 8) animals by enzyme-linked immunosorbent assay (ELISA) (J). DT administered from day 8 to 14 after birth followed by Listeria monocytogenes administration by oral gavage on day 49 and analyses of Ifnγ (K) or Il12p40 (L) mRNA expression in the colon of control littermates LysCre+/- (n = 6) or MMDTR (n = 6) animals 3 days after infection by quantitative polymerase chain reaction analysis. Numbers in the representative plots indicate cell frequency and were determined by flow cytometry. Error bars indicate standard error of mean. Each dot is representative of an individual mouse. P values were calculated by unpaired two-sided Student’s t-test. *P < 0.05, **P < 0.01, ns: not-significant. Numbers in the representative plots indicate cell frequency.
Extended Data Fig. 10
a, Gating strategy for iNKT (right panel) and TCR-αβ+ T (middle panel) cells identification by flow cytometry in the colon at day 12 (top) and day 49 (Adult) (bottom) after birth. b, Gating strategy for F4/80hi/CD11blo and F4/80lo/CD11bhi macrophages (right panel) identification by flow cytometry in the colon at day 12 (top) and day 49 (Adult) (bottom) after birth.
Supplementary information
Supplementary Tables
Supplementary Table 1: Differentially expressed transcripts comparing colonic macrophages from 8- and 14-day-old SPF animals by RNA-seq. DESeq analysis (Padj < 0.05). P values calculated by the Wald test are corrected for multiple testing using the Benjamini–Hochberg method. Supplementary Table 2: GO term analysis of the enriched transcripts in colonic macrophages at 8 days old as compared to 14 days old. Supplementary Table 3: Differentially expressed transcripts comparing colonic macrophages from 9-day-old SPF and germ-free animals by RNA-seq. DESeq analysis (Padj < 0.05). P values calculated by the Wald test are corrected for multiple testing using the Benjamini–Hochberg method. Supplementary Table 4: Differentially expressed transcripts comparing colonic iNKT cells from 14- and 56-day-old adult animals by RNA-seq. DESeq analysis (Padj < 0.05). P values calculated by the Wald test are corrected for multiple testing using the Benjamini–Hochberg method. Supplementary Table 5: GO term analysis of the enriched transcripts in colonic iNKT cells at 14 days old compared to 56 days old.
Rights and permissions
About this article

Cite this article
Gensollen, T., Lin, X., Zhang, T. et al. Embryonic macrophages function during early life to determine invariant natural killer T cell levels at barrier surfaces. Nat Immunol 22, 699–710 (2021). https://doi.org/10.1038/s41590-021-00934-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-021-00934-0
This article is cited by
-
Unconventional immune cells in the gut mucosal barrier: regulation by symbiotic microbiota
Experimental & Molecular Medicine (2023)
-
Macrophages in intestinal homeostasis and inflammatory bowel disease
Nature Reviews Gastroenterology & Hepatology (2023)
-
Developmentally programmed early-age skin localization of iNKT cells supports local tissue development and homeostasis
Nature Immunology (2023)
-
Macrophages make the bed for early iNKT cells
Nature Immunology (2021)
