
Abstract
Immune checkpoint blockade has provided a paradigm shift in cancer therapy, but the success of this approach is very variable; therefore, biomarkers predictive of clinical efficacy are urgently required. Here, we show that the frequency of PD-1+CD8+ T cells relative to that of PD-1+ regulatory T (Treg) cells in the tumor microenvironment can predict the clinical efficacy of programmed cell death protein 1 (PD-1) blockade therapies and is superior to other predictors, including PD ligand 1 (PD-L1) expression or tumor mutational burden. PD-1 expression by CD8+ T cells and Treg cells negatively impacts effector and immunosuppressive functions, respectively. PD-1 blockade induces both recovery of dysfunctional PD-1+CD8+ T cells and enhanced PD-1+ Treg cell–mediated immunosuppression. A profound reactivation of effector PD-1+CD8+ T cells rather than PD-1+ Treg cells by PD-1 blockade is necessary for tumor regression. These findings provide a promising predictive biomarker for PD-1 blockade therapies.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
The data that support the findings of this study are available from the corresponding author upon request. The exome sequencing data of patients’ tumor samples were deposited in the Japanese Genotype-phenotype Archive (accession number: JGAS00000000244).
References
Schreiber, R. D., Old, L. J. & Smyth, M. J. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science 331, 1565–1570 (2011).
Zou, W., Wolchok, J. D. & Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 8, 328rv324 (2016).
Dong, H. et al. Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat. Med. 8, 793–800 (2002).
Topalian, S. L. et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N. Engl. J. Med. 366, 2443–2454 (2012).
Carbone, D. P. et al. First-line nivolumab in stage IV or recurrent non-small-cell lung cancer. N. Engl. J. Med. 376, 2415–2426 (2017).
Rizvi, N. A. et al. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science 348, 124–128 (2015).
Rooney, M. S., Shukla, S. A., Wu, C. J., Getz, G. & Hacohen, N. Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell 160, 48–61 (2015).
Sharma, P., Hu-Lieskovan, S., Wargo, J. A. & Ribas, A. Primary, adaptive, and acquired resistance to cancer immunotherapy. Cell 168, 707–723 (2017).
Kansy, B. A. et al. PD-1 status in CD8+ T cells associates with survival and anti-PD-1 therapeutic outcomes in head and neck cancer. Cancer Res. 77, 6353–6364 (2017).
Gros, A. et al. PD-1 identifies the patient-specific CD8+ tumor-reactive repertoire infiltrating human tumors. J. Clin. Invest. 124, 2246–2259 (2014).
Tada, Y. et al. Targeting VEGFR2 with ramucirumab strongly impacts effector/activated regulatory T cells and CD8. J. Immunother. Cancer 6, 106 (2018).
Shitara, K. et al. Pembrolizumab versus paclitaxel for previously treated, advanced gastric or gastro-oesophageal junction cancer (KEYNOTE-061): a randomised, open-label, controlled, phase 3 trial. Lancet 392, 123–133 (2018).
Kim, S. T. et al. Comprehensive molecular characterization of clinical responses to PD-1 inhibition in metastatic gastric cancer. Nat. Med. 24, 1449–1458 (2018).
Thommen, D. S. et al. A transcriptionally and functionally distinct PD-1. Nat. Med. 24, 994–1004 (2018).
Daud, A. I. et al. Tumor immune profiling predicts response to anti-PD-1 therapy in human melanoma. J. Clin. Invest. 126, 3447–3452 (2016).
Wherry, E. J. & Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 15, 486–499 (2015).
Zehn, D., Lee, S. Y. & Bevan, M. J. Complete but curtailed T-cell response to very low-affinity antigen. Nature 458, 211–214 (2009).
Kamada, T. et al. PD-1+ regulatory T cells amplified by PD-1 blockade promote hyperprogression of cancer. Proc. Natl Acad. Sci. USA 116, 9999–10008 (2019).
Hsieh, C. S. et al. Recognition of the peripheral self by naturally arising CD25+CD4+ T cell receptors. Immunity 21, 267–277 (2004).
Tran, D. Q., Ramsey, H. & Shevach, E. M. Induction of FOXP3 expression in naive human CD4+FOXP3− T cells by T-cell receptor stimulation is transforming growth factor-β dependent but does not confer a regulatory phenotype. Blood 110, 2983–2990 (2007).
Saito, T. et al. Two FOXP3+CD4+ T cell subpopulations distinctly control the prognosis of colorectal cancers. Nat. Med. 22, 679–684 (2016).
Miyara, M. et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 30, 899–911 (2009).
Togashi, Y., Shitara, K. & Nishikawa, H. Regulatory T cells in cancer immunosuppression—implications for anticancer therapy. Nat. Rev. Clin. Oncol. 16, 356–371 (2019).
Yokosuka, T. et al. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J. Exp. Med. 209, 1201–1217 (2012).
Hui, E. et al. T cell costimulatory receptor CD28 is a primary target for PD-1-mediated inhibition. Science 355, 1428–1433 (2017).
Zhang, R. et al. An obligate cell-intrinsic function for CD28 in Tregs. J. Clin. Invest. 123, 580–593 (2013).
Levine, A. G., Arvey, A., Jin, W. & Rudensky, A. Y. Continuous requirement for the TCR in regulatory T cell function. Nat. Immunol. 15, 1070–1078 (2014).
Salmond, R. J., Huyer, G., Kotsoni, A., Clements, L. & Alexander, D. R. The src homology 2 domain-containing tyrosine phosphatase 2 regulates primary T-dependent immune responses and Th cell differentiation. J. Immunol. 175, 6498–6508 (2005).
Nguyen, T. V., Ke, Y., Zhang, E. E. & Feng, G. S. Conditional deletion of Shp2 tyrosine phosphatase in thymocytes suppresses both pre-TCR and TCR signals. J. Immunol. 177, 5990–5996 (2006).
Frearson, J. A. & Alexander, D. R. The phosphotyrosine phosphatase SHP-2 participates in a multimeric signaling complex and regulates T cell receptor (TCR) coupling to the Ras/mitogen-activated protein kinase (MAPK) pathway in Jurkat T cells. J. Exp. Med. 187, 1417–1426 (1998).
Huang, A. C. et al. T-cell invigoration to tumour burden ratio associated with anti-PD-1 response. Nature 545, 60–65 (2017).
Krieg, C. et al. High-dimensional single-cell analysis predicts response to anti-PD-1 immunotherapy. Nat. Med. 24, 144–153 (2018).
Subrahmanyam, P. B. et al. Distinct predictive biomarker candidates for response to anti-CTLA-4 and anti-PD-1 immunotherapy in melanoma patients. J. Immunother. Cancer 6, 18 (2018).
Takeuchi, Y. et al. Clinical response to PD-1 blockade correlates with a sub-fraction of peripheral central memory CD4+ T cells in patients with malignant melanoma. Int. Immunol. 30, 13–22 (2018).
Tumeh, P. C. et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature 515, 568–571 (2014).
Prasad, K. V. et al. T-cell antigen CD28 interacts with the lipid kinase phosphatidylinositol 3-kinase by a cytoplasmic Tyr(P)-Met-Xaa-Met motif. Proc. Natl Acad. Sci. USA 91, 2834–2838 (1994).
Stein, P. H., Fraser, J. D. & Weiss, A. The cytoplasmic domain of CD28 is both necessary and sufficient for costimulation of interleukin-2 secretion and association with phosphatidylinositol 3′-kinase. Mol. Cell Biol. 14, 3392–3402 (1994).
Truitt, K. E., Hicks, C. M. & Imboden, J. B. Stimulation of CD28 triggers an association between CD28 and phosphatidylinositol 3-kinase in Jurkat T cells. J. Exp. Med. 179, 1071–1076 (1994).
Hawse, W. F., Boggess, W. C. & Morel, P. A. TCR signal strength regulates Akt substrate specificity to induce alternate murine Th and T regulatory cell differentiation programs. J. Immunol. 199, 589–597 (2017).
Kamphorst, A. O. et al. Rescue of exhausted CD8 T cells by PD-1-targeted therapies is CD28-dependent. Science 355, 1423–1427 (2017).
Soond, D. R., Slack, E. C., Garden, O. A., Patton, D. T. & Okkenhaug, K. Does the PI3K pathway promote or antagonize regulatory T cell development and function? Front. Immunol. 3, 244 (2012).
Patton, D. T. et al. Cutting edge: the phosphoinositide 3-kinase p110δ is critical for the function of CD4+CD25+Foxp3+ regulatory T cells. J. Immunol. 177, 6598–6602 (2006).
Haxhinasto, S., Mathis, D. & Benoist, C. The AKT–mTOR axis regulates de novo differentiation of CD4+Foxp3+ cells. J. Exp. Med. 205, 565–574 (2008).
Strauss, L. et al. Selective survival of naturally occurring human CD4+CD25+Foxp3+ regulatory T cells cultured with rapamycin. J. Immunol. 178, 320–329 (2007).
Delgoffe, G. M. et al. The mTOR kinase differentially regulates effector and regulatory T cell lineage commitment. Immunity 30, 832–844 (2009).
Passerini, L. et al. Treatment with rapamycin can restore regulatory T-cell function in IPEX patients. J. Allergy Clin. Immunol. 145, 1262–1271 (2020).
Abu-Eid, R. et al. Selective inhibition of regulatory T cells by targeting the PI3K–Akt pathway. Cancer Immunol. Res. 2, 1080–1089 (2014).
Ali, K. et al. Inactivation of PI3K p110δ breaks regulatory T-cell-mediated immune tolerance to cancer. Nature 510, 407–411 (2014).
Sugiyama, E. et al. Blockade of EGFR improves responsiveness to PD-1 blockade in EGFR-mutated non-small cell lung cancer. Sci. Immunol. 5 eaav3937 (2020).
Van Gassen, S. et al. FlowSOM: using self-organizing maps for visualization and interpretation of cytometry data. Cytometry 87, 636–645 (2015).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
DePristo, M. A. et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 43, 491–498 (2011).
Hoof, I. et al. NetMHCpan, a method for MHC class I binding prediction beyond humans. Immunogenetics 61, 1 (2009).
Nielsen, M. et al. NetMHCpan, a method for quantitative predictions of peptide binding to any HLA-A and -B locus protein of known sequence. PLoS ONE 2, e796 (2007).
Sugiyama, D. et al. Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans. Proc. Natl Acad. Sci. USA 110, 17945–17950 (2013).
Hogquist, K. A. et al. T cell receptor antagonist peptides induce positive selection. Cell 76, 17–27 (1994).
Wang, L., Chu, F. & Xie, W. Accurate cancer classification using expressions of very few genes. IEEE/ACM Trans. Comput. Biol. Bioinform. 4, 40–53 (2007).
Wang, S. General constructive representations for continuous piecewise-linear functions. IEEE Trans. Circuits Syst. I Regul. Pap. 51, 1889–1896 (2004).
Acknowledgements
This article is dedicated to the memory of T. Kamada. We thank T. Kobayashi, Y. Tada, T. Takaku, M. Nakai, K. Onagawa, M. Takemura, C. Haijima, M. Hoshino, K. Yoshida, M. Sugaya, Y. Ishige, E. Tanji, M. Ozawa and C. Notake for their technical assistance. This study was supported by Grants-in-Aid for Scientific Research (S grant 17H06162 (H. Nishikawa), Challenging Exploratory Research grant 16K15551 (H. Nishikawa), Young Scientists grant 17J09900 (Y. Togashi), JSPS Research Fellowship 17K18388 (Y. Togashi) and JSPS Research Fellowship 18J21161 (T. Kamada) and 18J21551 (S.K.)) from the Ministry of Education, Culture, Sports, Science and Technology of Japan; by the Project for Cancer Research and Therapeutic Evolution (P-CREATE, 16cm0106301h0001 (H. Nishikawa), 16cm0106301h0001 (H.M.), 18cm0106502s0403 (M.K.) and 18cm0106340h0001 (Y. Togashi)); by a Development of Technology for Patient Stratification Biomarker Discovery grant (19ae0101074s0401 (H. Nishikawa)) and Leading Advanced Projects for Medical Innovation (LEAP, 18am0001001h9905 (H.M.)) from the Japan Agency for Medical Research and Development (AMED); by the National Cancer Center Research and Development Fund (28-A-7 and 31-A-7 (H. Nishikawa)); by the Naito Foundation (Y. Togashi and H. Nishikawa); by the Takeda Science Foundation (Y. Togashi); by the Kobayashi Foundation for Cancer Research (Y. Togashi); by a Novartis research grant (Y. Togashi); by a Bristol-Myers Squibb research grant (Y. Togashi) and by the SGH Foundation (Y. Togashi). This study was executed in part as a research program supported by Ono Pharmaceutical Co. Ltd.
Author information
Authors and Affiliations
Contributions
S.K., Y. Togashi, T. Kamada, E.S., H. Nishinakamura, Y. Takeuchi, K.V., K.I., Y.M., S.M., T.I., A.T., S.F., G.I., T. Kuwata, M.K., T.U., H.M. and H. Nishikawa performed the experiments and analyzed the data. A.K., H.U., K.K., K.A., K.N., N.Y., K.G., M.T., T.D. and K.S. collected the clinical specimens and performed the analyses of the clinical data. T.S. and Y.Y. designed the architecture of the statistical models. S.K., Y. Togashi, T. Kamada and H. Nishikawa conceived the project and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
Y. Togashi has received honoraria from Bristol-Myers Squibb, Ono Pharmaceutical, Chugai Pharmaceutical, MSD and Boehringer Ingelheim and has received research funding from AstraZeneca and KOTAI Biotechnologies outside the scope of this work. T.S. and Y.Y. are employees of Hitachi, Ltd. (Hitachi provided support in the form of salaries for authors T.S. and Y.Y. but did not have any additional role in the study). T. Kuwata has received research funding from Ono Pharmaceutical and has received honoraria from Chugai Pharmaceutical outside this work. K.G. has received honoraria and research funding from Bristol-Myers Squibb, Chugai Pharmaceutical and Ono Pharmaceutical and has received research funding from AstraZeneca, MSD, Merck Serono and F. Hoffmann–La Roche outside the scope of this work. H.M. has served as a member of the scientific advisory board for Chugai Pharmaceutical and has received research funding from Ono Pharmaceutical outside the scope of this work. T.D. has received research funding from Merck Serono, MSD, Chugai Pharmaceutical and Bristol-Myers Squibb outside the scope of this work. K.S. has served as a member of the scientific advisory board for Ono Pharmaceutical, Eli Lilly, Bristol-Myers Squibb, Astellas Pharma, Takeda and Pfizer; has received research funding from Ono Pharmaceutical, Eli Lilly, Sumitomo Dainippon Pharma, Daiichi Sankyo, Taiho Pharmaceutical, Chugai Pharmaceutical and MSD; and has received honoraria from Novartis, AbbVie and Yakult outside the scope of this work. H. Nishikawa received research funding from Ono Pharmaceutical and BD Japan for this work and has received honoraria and research funding from Chugai Pharmaceutical, MSD and Bristol-Myers Squibb; honoraria from Ono Pharmaceutical; and research funding from Taiho Pharmaceutical, Daiichi Sankyo, Kyowa Kirin, Zenyaku Kogyo, Oncolys BioPharma, Debiopharm, Asahi-Kasei, SRL, Fujifilm, Astellas Pharma, Sumitomo Dainippon Pharma and Sysmex outside the scope of this study. H. Nishikawa is the primary inventor on pending patents PCT/JP2019/02163 belonging to the National Cancer Center Japan and Ono Pharmaceutical and PCT/JP2020/0059919 belonging to the National Cancer Center Japan and BD Biosciences. T.S. has a pending US patent 16/117260 belonging to Hitachi, Ltd. All other authors declare no competing interests.
Additional information
Peer review information Zoltan Fehervari was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 CyTof tSNE plots of TILs and the relation between PD-1 expression and antigen affinity are shown.
(a) Fresh tumor samples obtained from primary or metastatic tumors by endoscopic or needle biopsy within two weeks before the initial drug administration were subjected to CyTOF. (b) Binding affinities of each peptide restricted by HLA−A*0201 were predicted with NetMHCpan (version 2.8). Sequences and predicted IC50 values of peptides used in this study are shown. (c) Predicted IC50 values for Melan A and Flu are shown. (d) Representative FCM staining of PD-1 expression (left) and summary for expression levels detected by Mean Fluorescence Intensity (MFI) of PD-1 in tetramer+CD8+ T cells (right). 1 × 105 HLA−A*0201 CD8+ T cells prepared from healthy individuals (n = 3) were cultured with antigen-presenting cells pulsed with natural Melan A or Flu peptides. Seven to 10 days later, PD-1 expression by Melan A or Flu-specific CD8+ T cells detected by MHC/peptide tetramers was examined. Paired t test was used. MelA, Melan A; Bars, mean; error bars, s.e.m.
Extended Data Fig. 2 Cytokine production by CD8+ T cells is attenuated by PD-L1 and recovered by anti-PD-1 mAb.
(a, c) Representative FCM staining (left) and summary (right) of IFN-γ production by CD8+ T cells are shown. CD8+ T cells purified from splenocytes of C57BL/6J (a) or Pdcd1-/-C57BL/6 (c) mice were stimulated with anti-CD3 mAb and anti-CD28 mAb, PD-L1-Ig and/or anti-PD-1 mAb as indicated for 48 hours (n = 3 per group). IFN-γ production by CD8+ T cells was analyzed with FCM. One-way ANOVA test with Bonferroni corrections was used. (b) MC-38 cells (5×105) were inoculated subcutaneously into C57BL/6J mice on day 0 and anti-PD-1 mAb or control mAb was administered on days 3, 6 (n = 6 tumors per group). On day 7, mice were sacrificed, and TILs were extracted from tumors and analyzed with FCM. Representative FCM staining (left) and summary (right) of IFN-γ and TNF-α production by CD8+ T cells from TILs are shown. Unpaired t test was used. These experiments were performed at least twice with similar results. Bars, mean; error bars, s.e.m.
Extended Data Fig. 3 Treg cell activation markers are down-regulated by PD-1 signaling and recovered by anti-PD-1 mAb.
(a, c) Representative FCM staining (left) and summary (right) of GITR, ICOS, and CTLA−4 expression by Foxp3+CD4+ T cells are shown. CD25+CD4+ T cells purified from splenocytes of C57BL/6J (wild-type; WT) (a) or Pdcd1-/-C57BL/6 (PD-1 KO) (c) mice were stimulated with anti-CD3 mAb and anti-CD28 mAb, PD-L1-Ig and/or anti-PD-1 mAb as indicated for 48 hours (n = 3 per group). The expression of GITR, ICOS and CTLA-4 in Foxp3+CD4+ T cells was analyzed with FCM. One-way ANOVA test with Bonferroni corrections was used. (b) MC-38 cells (5 × 105) were inoculated subcutaneously into C57BL/6J mice on day 0 and anti-PD-1 mAb or control mAb was administered on days 3, 6 (n = 5 per group). On day 7, mice were sacrificed, and TILs were extracted from tumors and analyzed with FCM. Representative FCM staining (left) and summary (right) of GITR, ICOS and CTLA-4 expression in Foxp3+CD4+ T cells from TILs are shown. Unpaired t-test was used. The experiments were performed at least twice with similar results. Bars, mean; error bars, s.e.m..
Extended Data Fig. 4 The activation of TCR and CD28 signals in human T cells is affected by PD-1 blockade.
(a, b) Representative FCM staining (left) and summary (right) of pZAP70, pAKT and pSHP2 expression by CD8+ T cells (a) and CD45RA-FoxP3hiCD4+ T cells (b) are shown. CD8+ T cells (a) or CD45RA-FoxP3hiCD4+ T cells (b) sorted from PBMCs of healthy individuals (n = 4 per group) were stimulated with anti-CD3 mAb and anti-CD28 mAb, PD-L1-Ig and/or anti-PD-1 mAb as indicated for 48 hours. The expression of pZAP70, pAKT and pSHP2 by CD8+ T cells and CD45RA-FoxP3hiCD4+ T cells was analyzed with FCM. One-way ANOVA test with Bonferroni corrections was used. (c) Representative FCM staining (left) and summary (right) for the fold changes of Treg cell activation markers after treatment with anti-PD-1 mAb are shown. The expression of pZAP70, pAKT and pSHP2 by eTreg cells from TILs was analyzed with FCM after 48 hours of stimulation by anti-CD3 mAb and anti-CD28 mAb in the presence/absence of anti-PD-1 mAb (n = 5 per group). Paired t-test was used. Bars, mean; error bars, s.e.m.
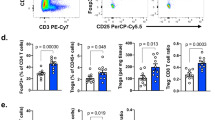
Extended Data Fig. 5 PD-1 expression balance of T cells in the TME, not in PBMCs, correlates with PD-1 blockade efficacy.
(a) The boundaries classified by the piece-wise linear model was generated by training with the discovery cohort. (b) The boundaries were applied into the validation cohort. The red color shading in each panel represents the responder probability. The AUC values of the piece-wise linear model in the discovery cohort (a) and the validation cohort (b) were 0.991 and 0.968, respectively. (c-g) Fresh tumor samples obtained from primary or metastatic tumors by endoscopic or needle biopsy and PBMCs within 2 weeks before the initial drug administration were subjected to FCM. Correlation of PD-1+ (%) in CD8+ T cells (c) and eTreg cells (d) in PBMCs and TILs in the validation cohort is shown. (e, f) PD-1 expression by CD8+ T cells (e) or eTreg cells (f) in PBMCs from responders and non-responders is shown. Unpaired t test was used (All, R, n = 13, NR, n = 31; NSCLC, R, n = 5, NR, n = 7; GC, R, n = 3, NR, n = 19; MM, R, n = 5, NR, n = 5). (g) Scatter plots using %PD-1+ in CD8+ T cells and eTreg cells in PBMCs is shown.
Extended Data Fig. 6 tSNE plots are shown for representative markers in CyTOF analysis for PBMCs.
Blood samples obtained from patients in the validation cohort within two weeks before the initial drug administration were subjected to CyTOF. Representative expression patterns of markers are shown.
Extended Data Fig. 7 Any biomarkers with PBMCs do not predict clinical responses to PD-1 blockade therapies.
(a–d) CD45RA+CCR7+ (%) (a), CD45RA+CCR7−(%) (b), CD45RA−CCR7+ (%) (c), and CD45RA−CCR7− (%) (d) in CD4+ cells in PBMCs from responders and non-responders are shown. (e-h) CD45RA+CCR7+ (%) (e), CD45RA+CCR7− (%) (f), CD45RA−CCR7+ (%) (g), and CD45RA−CCR7− (%) (h) in CD8+ cells in PBMCs from responders and non-responders are shown. (k) Ki-67+ (%) in CD8+ cells in PBMCs from responders and non-responders is shown. (j) CD14+CD16−HLA-DRhigh (%) in CD45+ cells in PBMCs from responders and non-responders is shown. (k) CD69+ (%) in CD3−CD56+ cells in PBMCs from responders and non-responders is shown. Unpaired t-test was used (All, R, n = 12, NR, n = 29; NSCLC, R, n = 5, NR, n = 7; GC, R, n = 2, NR, n = 17; MM, R, n = 5, NR, n = 5).
Extended Data Fig. 8 TMB and PD-L1 expression show a slight correlation with efficacy of PD-1 blockade therapies.
(a) Kaplan–Meier curves for PFS of GC patients treated with anti-PD-1 mAb according to MMR status in the validation cohort. (b, c) Eight NSCLC, nine GC and five MM samples in the validation cohort were subjected to whole-exome sequencing. ROC curves for TMB (missense variants) were developed by plotting the true positive rate against the false positive rate at each threshold setting. AUC (0.64, 95%CI: 0.38-0.91) shown in each plot summarizes the performance of TMB (b). Kaplan–Meier curves for PFS of patients (the validation cohort) treated with PD-1 blockade therapies in the whole (upper left), NSCLC (upper right), GC (lower left) and MM (lower right) according to TMB. To compare PFS, the cut-off of TMB derived from ROC analysis was used (c). (d) Kaplan–Meier curves for PFS of patients (the validation cohort) treated with anti-PD-1 mAb in the whole (left), NSCLC (middle) and GC (right) according to PD-L1 expression. To compare PFS, PD-L1 expression by tumor cells of ≥ 1% was defined as positive. (a, c, d) Log-rank test was employed.
Supplementary information
Supplementary Information
Supplementary Figs. 1–3.
Supplementary Tables
Supplementary Tables 1–12.
Supplementary Data
Point-wise linear models and feature importance are shown.
Rights and permissions
About this article

Cite this article
Kumagai, S., Togashi, Y., Kamada, T. et al. The PD-1 expression balance between effector and regulatory T cells predicts the clinical efficacy of PD-1 blockade therapies. Nat Immunol 21, 1346–1358 (2020). https://doi.org/10.1038/s41590-020-0769-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-020-0769-3
This article is cited by
-
Hypothesis of a CD137/Eomes activating axis for effector T cells in HPV oropharyngeal cancers
Molecular Medicine (2024)
-
Potential anti-tumor effects of regulatory T cells in the tumor microenvironment: a review
Journal of Translational Medicine (2024)
-
Regulatory T cell-mediated immunosuppression orchestrated by cancer: towards an immuno-genomic paradigm for precision medicine
Nature Reviews Clinical Oncology (2024)
-
Insights into vaccines for elderly individuals: from the impacts of immunosenescence to delivery strategies
npj Vaccines (2024)
-
New prospects of cancer therapy based on pyroptosis and pyroptosis inducers
Apoptosis (2024)
