
Abstract
Activation-induced cell death (AICD) of T lymphocytes can be exploited by cancers to escape immunological destruction. We demonstrated that tumor-specific cytotoxic T lymphocytes (CTLs) and type 1 helper T (TH1) cells, rather than type 2 helper T cells and regulatory T cells, were sensitive to AICD in breast and lung cancer microenvironments. NKILA, an NF-κB-interacting long noncoding RNA (lncRNA), regulates T cell sensitivity to AICD by inhibiting NF-κB activity. Mechanistically, calcium influx in stimulated T cells via T cell–receptor signaling activates calmodulin, thereby removing deacetylase from the NKILA promoter and enhancing STAT1-mediated transcription. Administering CTLs with NKILA knockdown effectively inhibited growth of breast cancer patient-derived xenografts in mice by increasing CTL infiltration. Clinically, NKILA overexpression in tumor-specific CTLs and TH1 cells correlated with their apoptosis and shorter patient survival. Our findings underscore the importance of lncRNAs in determining tumor-mediated T cell AICD and suggest that engineering lncRNAs in adoptively transferred T cells might provide a novel antitumor immunotherapy.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others


References
Schietinger, A. & Greenberg, P. D. Tolerance and exhaustion: defining mechanisms of T cell dysfunction. Trends Immunol. 35, 51–60 (2014).
Gati, A. et al. CD158 receptor controls cytotoxic T-lymphocyte susceptibility to tumor-mediated activation-induced cell death by interfering with Fas signaling. Cancer Res. 63, 7475–7482 (2003).
Grimm, M. et al. Tumour-mediated TRAIL-receptor expression indicates effective apoptotic depletion of infiltrating CD8+ immune cells in clinical colorectal cancer. Eur. J. Cancer 46, 2314–2323 (2010).
Kassouf, N. & Thornhill, M. H. Oral cancer cell lines can use multiple ligands, including Fas-L, TRAIL and TNF-alpha, to induce apoptosis in Jurkat T cells: possible mechanisms for immune escape by head and neck cancers. Oral Oncol. 44, 672–682 (2008).
Wang, K. C. & Chang, H. Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 43, 904–914 (2011).
Heward, J. A. & Lindsay, M. A. Long non-coding RNAs in the regulation of the immune response. Trends Immunol. 35, 408–419 (2014).
Atianand, M. K. et al. A long noncoding RNA lincRNA-EPS acts as a transcriptional brake to restrain inflammation. Cell 165, 1672–1685 (2016).
Wang, P. et al. The STAT3-binding long noncoding RNA lnc-DC controls human dendritic cell differentiation. Science 344, 310–313 (2014).
Ranzani, V. et al. The long intergenic noncoding RNA landscape of human lymphocytes highlights the regulation of T cell differentiation by linc-MAF-4. Nat. Immunol. 16, 318–325 (2015).
Hu, G. et al. Expression and regulation of intergenic long noncoding RNAs during T cell development and differentiation. Nat. Immunol. 14, 1190–1198 (2013).
Haanen, J. B. et al. In situ detection of virus- and tumor-specific T-cell immunity. Nat. Med. 6, 1056–1060 (2000).
Disis, M. L. et al. Concurrent trastuzumab and HER2/neu-specific vacactivated CTLs (n = 3 independentcination in patients with metastatic breast cancer. J. Clin. Oncol. 27, 4685–4692 (2009).
Lee, H. J. et al. Expression of NY-ESO-1 in triple-negative breast cancer is associated with tumor-infiltrating lymphocytes and a good prognosis. Oncology 89, 337–344 (2015).
Rentzsch, C. et al. Evaluation of pre-existent immunity in patients with primary breast cancer: molecular and cellular assays to quantify antigen-specific T lymphocytes in peripheral blood mononuclear cells. Clin. Cancer Res. 9, 4376–4386 (2003).
Zhu, J. & Paul, W. E. CD4 T cells: fates, functions, and faults. Blood 112, 1557–1569 (2008).
Tauriello, D. V. F. et al. TGFβ drives immune evasion in genetically reconstituted colon cancer metastasis. Nature 554, 538–543 (2018).
Mansfield, A. S. et al. Regional immunity in melanoma: immunosuppressive changes precede nodal metastasis. Mod. Pathol. 24, 487–494 (2011).
Su, S. et al. Blocking the recruitment of naive CD4+ T cells reverses immunosuppression in breast cancer. Cell Res. 27, 461–482 (2017).
Krammer, P. H., Arnold, R. & Lavrik, I. N. Life and death in peripheral T cells. Nat. Rev. Immunol. 7, 532–542 (2007).
Ankersmit, H. J. et al. Activation-induced T-cell death and immune dysfunction after implantation of left-ventricular assist device. Lancet 354, 550–555 (1999).
Li-Weber, M. et al. Vitamin E inhibits CD95 ligand expression and protects T cells from activation-induced cell death. J. Clin. Invest. 110, 681–690 (2002).
Rapoport, A. P. et al. NY-ESO-1-specific TCR-engineered T cells mediate sustained antigen-specific antitumor effects in myeloma. Nat. Med. 21, 914–921 (2015).
Norell, H. et al. Inhibition of superoxide generation upon T-cell receptor engagement rescues Mart-1(27-35)-reactive T cells from activation-induced cell death. Cancer Res. 69, 6282–6289 (2009).
Strønen, E. et al. Targeting of cancer neoantigens with donor-derived T cell receptor repertoires. Science 352, 1337–1341 (2016).
Feuerer, M. et al. Therapy of human tumors in NOD/SCID mice with patient-derived reactivated memory T cells from bone marrow. Nat. Med. 7, 452–458 (2001).
Han, W. et al. RNA-binding protein PCBP2 modulates glioma growth by regulating FHL3. J. Clin. Invest. 123, 2103–2118 (2013).
Beckhove, P. et al. Specifically activated memory T cell subsets from cancer patients recognize and reject xenotransplanted autologous tumors. J. Clin. Invest. 114, 67–76 (2004).
Rubio, V. et al. Ex vivo identification, isolation and analysis of tumor-cytolytic Tcells. Nat. Med. 9, 1377–1382 (2003).
Becker, C. et al. Adoptive tumor therapy with T lymphocytes enriched through an IFN-γ capture assay. Nat. Med. 7, 1159–1162 (2001).
Cappuzzo, F. et al. EGFR FISH assay predicts for response to cetuximab in chemotherapy refractory colorectal cancer patients. Ann. Oncol. 19, 717–723 (2008).
Liu, B. et al. A cytoplasmic NF-κB interacting long noncoding RNA blocks IκB phosphorylation and suppresses breast cancer metastasis. Cancer Cell 27, 370–381 (2015).
Fuertes, M. B. et al. Host type I IFN signals are required for antitumor CD8+ T cell responses through CD8alpha+ dendritic cells. J. Exp. Med. 208, 2005–2016 (2011).
Youn, H. D. & Liu, J. O. Cabin1 represses MEF2-dependent Nur77 expression and T cell apoptosis by controlling association of histone deacetylases and acetylases with MEF2. Immunity 13, 85–94 (2000).
Liu, J. O. Calmodulin-dependent phosphatase, kinases, and transcriptional corepressors involved in T-cell activation. Immunol. Rev. 228, 184–198 (2009).
Gu-Trantien, C. et al. CD4+ follicular helper T cell infiltration predicts breast cancer survival. J. Clin. Invest. 123, 2873–2892 (2013).
Ruffell, B., DeNardo, D. G., Affara, N. I. & Coussens, L. M. Lymphocytes in cancer development: polarization towards pro-tumor immunity. Cytokine Growth Factor Rev. 21, 3–10 (2010).
Palucka, A. K. & Coussens, L. M. The basis of oncoimmunology. Cell 164, 1233–1247 (2016).
Dong, H. et al. Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat. Med. 8, 793–800 (2002).
Motz, G. T. et al. Tumor endothelium FasL establishes a selective immune barrier promoting tolerance in tumors. Nat. Med. 20, 607–615 (2014).
Jiang, R. et al. The long noncoding RNA lnc-EGFR stimulates T-regulatory cells differentiation thus promoting hepatocellular carcinoma immune evasion. Nat. Commun. 8, 15129 (2017).
Stagg, J., Johnstone, R. W. & Smyth, M. J. From cancer immunosurveillance to cancer immunotherapy. Immunol. Rev. 220, 82–101 (2007).
Rosenberg, S. A. & Restifo, N. P. Adoptive cell transfer as personalized immunotherapy for human cancer. Science 348, 62–68 (2015).
Chhabra, A. TCR-engineered, customized, antitumor T cells for cancer immunotherapy: advantages and limitations. ScientificWorldJournal 11, 121–129 (2011).
Gutschner, T. et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 73, 1180–1189 (2013).
Beil-Wagner, J. et al. T cell-specific inactivation of mouse CD2 by CRISPR/Cas9. Sci. Rep. 6, 21377 (2016).
Yao, Y. D. et al. Targeted delivery of PLK1-siRNA by ScFv suppresses Her2+ breast cancer growth and metastasis. Sci. Transl. Med. 4, 130ra48 (2012).
Su, S. et al. CD10+GPR77+ cancer-associated fibroblasts promote cancer formation and chemoresistance by sustaining cancer stemness. Cell 172, 841–856.e816 (2018).
Chen, J. et al. CCL18 from tumor-associated macrophages promotes breast cancer metastasis via PITPNM3. Cancer Cell 19, 541–555 (2011).
Rossi, R. L. et al. Distinct microRNA signatures in human lymphocyte subsets and enforcement of the naive state in CD4+ T cells by the microRNA miR-125b. Nat. Immunol. 12, 796–803 (2011).
Dieu-Nosjean, M. C. et al. Long-term survival for patients with non-small-cell lung cancer with intratumoral lymphoid structures. J. Clin. Oncol. 26, 4410–4417 (2008).
Brenner, D., Golks, A., Kiefer, F., Krammer, P. H. & Arnold, R. Activation or suppression of NFκB by HPK1 determines sensitivity to activation-induced cell death. EMBO J. 24, 4279–4290 (2005).
Sun, T. et al. FASL -844C polymorphism is associated with increased activation-induced T cell death and risk of cervical cancer. J. Exp. Med. 202, 967–974 (2005).
Tang, J., Flomenberg, P., Harshyne, L., Kenyon, L. & Andrews, D. W. Glioblastoma patients exhibit circulating tumor-specific CD8+T cells. Clin. Cancer Res. 11, 5292–5299 (2005).
Su, S. et al. miR-142-5p and miR-130a-3p are regulated by IL-4 and IL-13 and control profibrogenic macrophage program. Nat. Commun. 6, 8523 (2015).
Wang, G. L. et al. Interaction between Cl- channels and CRAC-related Ca2+signaling during T lymphocyte activation and proliferation. Acta Pharmacol. Sin. 27, 437–446 (2006).
Acknowledgements
This work was supported by grants from the National Key Research and Development Program of China (2017YFA0106300 (S.S.) and 2016YFC1302300 (E.S.)), Natural Science Foundation of China (81720108029 (E.S.), 81622036 (S.S.),81472468 (S.S.), 81672614 (S.S.), 81621004 (E.S.), and 81490750 (E.S.)), Guangdong Science and Technology Department (2016A030306023 (S.S.), 2017A030313878 (S.S.), 2016B030229004 (E.S.), 2015B050501004 (E.S.), and 201710010083 (E.S.)), Tip-top Scientific and Technical Innovative Youth Talents of Guangdong Special Support Program (2016TQ03R553 (S.S.)), and Guangzhou Science Technology and Innovation Commission (201508020008 (E.S.) and 201508020249 (E.S.)).
Author information
Authors and Affiliations
Contributions
D.H., S.S., and E.S. conceived the ideas and designed the experiments. D.H., J.C., L.Y., Q.O., J.L., Y.L., J.Z., F.C., and Y.X. performed the experiments. H.Y., Q.L., F.S., and E.S. provided the patient samples for clinical data analysis and the PDX model. S.S., J.C., D.H., J.L., and L.L. analyzed the data. D.H., S.S., and E.S. wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 The identification and isolation of T cell subsets and the expression of apoptosis-inducing ligands and receptors in breast and lung cancer.
(a) Representative flow cytometric staining for HLA-A2 in the PBMC isolated from the peripheral blood of breast cancer patients (red: isotype, blue: anti-HLA-A2) (n = 576). (b) Representative images for immunohistochemical MUC1, HER2 and NY-ESO-1 staining in human breast cancer (n = 178) and normal breast tissue (n = 5). Scale bar, 50 μm. (c) Representative flow cytometric staining for MUC1-tetramer, HER2/neu-tetramer or NY-ESO-1-tetramer and anti-human CD8 antibody in the gated CD8+ T cells isolated from breast cancers (n = 5). The numbers indicate the percentages of tetramer+ CTLs. (d) Quantitation of FasL, TNF or TRAIL expression in the tumor cells of breast (n = 178) and lung cancer (n = 107) are shown. Boxes extend from the twenty-fifth to the seventy-fifth percentiles, and whiskers extend from the minimum values; lines indicate medians. (e) Representative images for immunohistochemical FasL, TRAIL and TNF staining in the breast tumor cells (n = 178) with indicated scoring quantification. Scale bar, 100 μm. (f-h) Tumor-specific CTLs (CD8+MUC1-tetramer+) and the paired naive (CD8+CD45RA+CD45RO-) and non-tumor-specific memory CTLs (CD8+CD45RA-CD45RO+MUC1-tetramer-) were isolated from 5 breast cancer patients. (f) Identification of tumor-specific CTLs and the phenotype of the isolated CD8+ T cells were determined by flow cytometric analysis for MUC1-tetramer, CD45RA and CD45RO staining. The numbers denote the percentages of CD45RO+CD45RA- (blue) and CD45RO-CD45RA+ (red) cells (mean±SEM, n = 5). (g) ELISPOT assay for IFN-γ was performed in the isolated CD8+ T cells (n = 4). (h) The isolated CD8+ T cells were incubated with deep red cell tracker-labeled autologous breast cancer cells at different E:T ratios for 12 h and tumor cell death was examined by PI uptake using flow cytometry. The numbers indicate the percentages of PI+ tumor cells (mean ± SEM, n = 4; ***, P<0.001 compared with naive CD8+ T cells). (i) Identification of TH1, TH2 and Treg by flow cytometry. Lymphocytes were gated as enriched lymphoid cell populations with low-granularity. Next, T cell subsets were identified and isolated on the basis of CD3, CD4 and respective surface marker (TH1: CD3+CD4+CXCR3+, TH2: CD3+CD4+CRTH2+CXCR3-, Treg: CD3+CD4+CD25+; n = 5). (j) The expression of IFNG, IL4, TGFB1 and IL10 in different T cell subsets isolated from peripheral blood of breast cancer patients was evaluated by qRT-PCR (mean ± SEM, n = 5,*** P<0.001 compared with TH1). (k) The mean fluorescence intensity (MFI) of Fas, TRAILR (DR4) and TNFR (TNFR2) expression was determined by FACS in various subsets of tumor-infiltrating T cells from breast (mean±SEM, n = 5, P>0.05) and lung cancer patients (mean±SEM, n = 5, P>0.05). P values by non-paired two tailed Student’s t-test (h, j, k).
Supplementary Figure 2 Tumor-antigen-activated CTLs are susceptible to FasL-mediated AICD as a result of NF-κB suppression.
(a) Scheme of tumor-antigen activated CTLs induction and the AICD sensitivity switch during T-cell activation. (b) The breast tumor-antigen activated CTLs were incubated with deep red cell tracker-labeled autologous breast cancer cells for 12 h. Tumor cell death was examined by PI uptake using flow cytometry. The numbers indicate the percentages of PI+ tumor cells (mean ± SEM, n = 4; **, P<0.01 compared with unprimed CTLs (day 0); d1: P=0.3905; d6: P=0026). (c) MUC1-specific CTLs were incubated with deep red cell tracker-labeled MUC1-loaded (T2/MUC1), HER2-loaded (T2/HER2) or unloaded T2 cells (T2) for 12 h. T2 cell death was examined by PI uptake using flow cytometry. The numbers indicate the percentages of PI+ T2 cells (mean ± SEM, n = 4; ***, P<0.001 compared with unprimed CTLs (day 0)). (d,e,i) Tumor-antigen activated CTLs were generated by co-culturing CD8+ T cells isolated from HLA-A2+ healthy donors with the autologous DCs pulsed with MDA-MB-231, NCI-H1703, MCF-7 or PC-9 lysates for indicated days. (d) The tumor-antigen activated CTLs were incubated with CFSE cell tracker-labeled relative cancer cell line and the tumor cell death was examined by PI uptake using flow cytometry. The curves correspond to the percentages of PI+ tumor cells (mean ± SEM, n = 3, **P < 0.01, ***P < 0.001, compared with the CD8+ T cells co-cultured with naive DCs (control)). (e, i) Apoptosis of tumor-antigen activated CTLs induced by relative tumor cells (e) or rFasL (i) was determined by Annexin V/PI staining and the curves correspond to specific-induced apoptotic rates (mean ± SEM, n = 3, **P < 0.01, ***P < 0.001, compared with the CD8+ T cells co-cultured with naive DCs (control)). (f) Western blotting was performed for FasL expression in various breast and lung cancer cell lines (n = 3). (g, h) The tumor-antigen activated CD8+ T cells were retrieved from the co-cultures with MDA-MB-231 lysate-pulsed DCs for 6 days. Apoptosis of the primed CTLs induced by tumor cells (g) or anti-CD3 (h) with or without anti-FasL or TNF-α neutralizing antibody was determined by Annexin V/PI staining. The numbers indicate the percentages of Annexin V+ cells (mean ± SEM, n = 3, ***, P<0.001 compared with untreated tumor-antigen activated CTLs that had been primed for 6 days (-)). (j-m) The tumor-antigen activated CTLs were generated by co-culturing CD8+ T cells with DCs pulsed with MDA-MB-231 lysates for indicated days. (j) The expression of BCL2, BCL2L1, IER3 and GADD45B in the CTLs was evaluated by qRT-PCR (mean ±SEM, n = 3, ** P < 0.01, *** P < 0.001 compared with unactivated CD8+ T cells (day 0)). (k) P65 nuclear translocation in indicated CTLs was assessed by immunofluorescent staining (n = 3). Scale bar, 10μm. (l) NF-κB activities of indicated CTLs were examined by EMSA (n = 3). (m) Western blotting was performed for total and phosphorylated IKK and IκBα in the primed CTLs (n = 3). P-values, unpaired two-tailed Student’s t-test (b-e, g-j).
Supplementary Figure 3 lncRNA NKILA is upregulated in tumor-specific T cells and interacts with the NF-κB–IκB complex via direct binding to p65.
(a) The expression of lncRNA XLOC_011899, RP11-854K16.3, RP11-315I20.3 and NKILA in CTLs primed by MDA-MB-231 lysate-pulsed DCs was evaluated by qRT-PCR (mean±SEM, n = 3; *, P<0.05, **, P<0.01, ***P< 0.001). (b) Binding of lncRNA XLOC_011899, RP11-854K16.3, RP11-315I20.3 and NKILA to p65/IκBα complex in d0 and d6 CTLs primed by MDA-MB-231 lysate-pulsed DCs, shown by RNA immunoprecipitation and qRT-PCR (mean ± SEM, n = 3, *P < 0.05, **P < 0.01, ***P< 0.001 versus IgG). (c) CD8+ T cells were transduced with shRNAs againt each of the lncRNAs and co-cultured with MDA-MB-231 lysate-pulsed DCs for 6 days. Apoptosis of the CTLs induced by anti-CD3 was determined by Annexin V/PI staining and the bars correspond to anti-CD3-induced apoptotic rates (mean ± SEM, n = 3; ***P< 0.001). (d) The peptides of NKILA-interacting p65 (RELA), IκBα (NFKBIA), p50 (RELB), ANXA2, PKM and LDHA identified in mass spectrum were shown with the amino-acid sequence (n = 2). (e) In vitro interaction between NKILA and p65, p50, IκBα, PKM, LDHA or ANXA2, shown by RNA pulldown (n = 3). (f) NKILA expression dynamics were assayed by qRT-PCR in CD8+ T cells primed by DCs pulsed with MUC1-peptide, MDA-MB-231 lysates, NCI-H1703 lysates, MCF-7 lysates or PC-9 lysates for indicated days (mean ± SEM, n = 4, **P < 0.01, ***P < 0.001, compared with the CD8+ T cells co-cultured with naive DCs (control)). (g-i) Tumor-antigen activated CTLs were generated by co-culturing CD8+ T cells with DCs pulsed with MDA-MB-231 lysates for indicated days. (g) NKILA expression was examined by Northern blot (n = 3). (h) Confocal FISH images demonstrated NKILA expression in the cytoplasm of CTLs (n = 3). Scale bar, 10μm. (i) Estimated copy numbers of NKILA per T cell. Equivalent molecules per cell were calculated based on the assumption that total RNA per cell is 20 pg. Red dots indicate molecules per cell in the resting and d6 CTLs (n = 3). P-values, unpaired two-tailed Student’s t-test (a-c, f).
Supplementary Figure 4 NKILA lncRNA promotes AICD of tumor-specific T cells via suppression of NF-κB.
T cells were transduced with lentivirus carrying an NKILA expression cassette (NKILA) or an empty vector (vec), or NKILA-shRNA (shNK1, shNK2), GFP-shRNA (shGFP) or shRNA control vector (shvec), followed by PHA activation (a, f, n) or co-culturing with autologous DCs pulsed with MUC1-peptide (b, c), MDA-MB-231/NCI-H1703 lysates (d, e, j-m) or primary tumor lysates (g, h, i) for indicated days. (a) NKILA expression was evaluated by qRT-PCR in the indicated T cells (mean ± SEM, n = 4; NS, not significant, P>0.05; **P<0.01; ***P<0.001). (b) Apoptosis of MUC1-specific CTLs induced by MUC1-loaded T2 cells (gray) or anti-CD3 (red) was determined by Annexin V/PI staining and the bars correspond to specific-induced apoptotic rates (mean ± SEM, n = 3, ***P<0.001 compared with shvec-transduced CTLs with indicated treatment). (c) CD8+ T cells were incubated with deep red cell tracker-labeled T2/MUC1 for 12 h. T2 cell death was examined by PI uptake using flow cytometry and the bars correspond to the percentages of PI+ T2 cells (mean ± SEM, n = 3; **, P<0.01). (d) Apoptosis of tumor-antigen activated CTLs induced by relevant tumor cells was determined by Annexin V/PI staining and the bars correspond to tumor cell-induced apoptotic rates (mean ± SEM, n = 3, ***P<0.001 compared with the CTLs transduced with shvec). (e) CD8+ T cells were incubated with CFSE cell tracker-labeled tumor cells for 12 h. Tumor cell death was examined by PI uptake using flow cytometry and the bars correspond to the percentages of PI+ tumor cells (mean ± SEM, n = 3; **, P<0.01; ***, P<0.001 compared with the T cells transduced with shvec). (f) Apoptosis of the indicated T cells induced by anti-CD3 was determined by Annexin V/PI staining. The numbers indicate the percentages of Annexin V+ cells (mean ± SEM, n = 4). ***, P<0.001 compared with T cells transduced with NKILA without anti-CD3 treatment. (g) Apoptosis of the indicated breast tumor-antigen activated CTLs induced by autologous tumor cells was determined by Annexin V/PI staining. The numbers indicate the percentages of Annexin V+ cells (mean ± SEM, n = 4). ***, P<0.001 compared with CTLs transduced with NKILA without tumor re-stimulation. (h) CD8+ T cells were incubated with deep red cell tracker-labeled autologous tumor cells for 12 h. Tumor cell death was examined by PI uptake using flow cytometry. The numbers indicate the percentages of PI+ tumor cells (mean ± SEM, n = 4; *P<0.05 compared with the T cells transduced with vec). (i) Apoptosis of the primed CTLs induced by co-culturing with autologous primary breast tumor cells pre-incubated with anti-HLA class I blocking antibody or control IgG antibody was determined by Annexin V/PI staining and the bars correspond to the percentages of Annexin V+ cells (mean ± SEM, n = 3, ***, P<0.001). (j) Apoptosis of the indicated CTLs induced by FasL+ MDA-MB-231 cells with or without anti-FasL antibody treatment was determined by Annexin V/PI staining. The numbers indicate the percentages of Annexin V+ cells (mean ± SEM, n = 3; **, P<0.01; ***, P<0.001 compared with NKILA-transduced T cells). (k) Immunoblotting was performed for total and phosphorylated IKK and IκBα in the indicated CTLs (n = 3). (l) P65 nuclear translocation in CTLs was assessed by immunofluorescent staining (n = 3). Scale bar, 10μm. (m) NF-κB activities of the CTLs were examined by EMSA (n = 3). The EMSA whole unchopped blot was shown below. (n) The expression of BCL2, BCL2L1, IER3 and GADD45B was examined by qRT-PCR (mean ± SEM, n = 3; *P<0.05). P-values, unpaired two-tailed Student’s t-test (a-j, n).
Supplementary Figure 5 NKILA is an effective target for adoptive T-cell therapy (ACT) for breast cancer patient-derived xenografts (PDX).
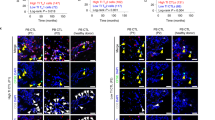
(a) Representative ER−PR−HER2−/ER−PR−HER2+ PDXs are shown in comparison with the original tumor samples. Tumor identification numbers and molecular subtyping are shown on the left. Tissue sections of the primary breast tumors from the patients and of the corresponding PDXs from the same individuals were stained in H&E and immunostained for ER, PR, HER2, Ki67 and FasL (n = 7 cases of PDX). Scale bars, 100 μm. (b) The transduction efficacy of CD8+ T cells was determined by flow cytometric staining of firefly luciferase. The numbers indicate the percentages of luciferase+ T cells (mean ± SEM, for n = 3, P<0.0001). (c, d) NKILA expression in the T cells before injection into PDX-bearing mice (c) and those isolated from the harvested PDX 4 weeks after injection (d) was evaluated by qRT-PCR (mean ± SEM, n = 4 cases of PDX per group; ***P<0.001). (e) PDX growth rates determined by tumor volume with indicated treatment are shown (mean ± SEM, n = 3 mice per group per PDX case; **P < 0.01, ***P < 0.001 compared to the PDX-bearing mice without ACT treatment). (f) The expression of BCL2, BCL2L1, IER3 and GADD45B in the CD8+ T cells isolated from the indicated PDXs. Bars correspond to mean ± SEM (n = 7 cases of PDX per group). **, P<0.01, ***, P<0.001. BCL2, P<0.0001; BCL2L1, P<0.0001; IER3, P=0.0065; GADD45B, P=0.0003. (g) Immunohistochemistry staining for MUC1 in the PDX and the primary tumor of the case SYMH005. Scale bars, 100 μm. (h) PDX-infiltrating CTLs (SYMH005) were incubated with deep red cell tracker-labeled T2, T2/MUC1 or T2/HER2 cells for 12 h and T2 cell death was assessed by PI uptake using flow cytometry. The numbers shown indicate the percentages of PI+ T2 cells (mean ± SEM, n = 3 mice; ***, P<0.001 compared with the percentages of unloaded T2 cell death). (i) Intracellular perforin levels and surface-marker CD107a expression in the isolated PDX-infiltrating CTLs (SYMH005) challenged by the T2, T2/MUC1 or T2/HER2 were determined by flow cytometry. Numbers indicate the percentages of gated cells stained for perforin or CD107a (mean ± SEM, n = 3 mice; **, P<0.01 compared with the untreated PDX-infiltrating CTLs (-). Perforin: P=0.0036; CD107a: P=0.0086). P-values, unpaired two-tailed Student’s t-test (b-f, h, i).
Supplementary Figure 6 Tumor-specific T cells from patients with breast and lung cancer have high amounts of NKILA and are sensitive to AICD.
(a) The expression of BCL2, BCL2L1, IER3 and GADD45B, normalized to GAPDH, in different T cell subsets isolated from peripheral blood (red) and tumor tissues (green) of breast cancer patients was evaluated by qRT-PCR (mean ± SEM, n = 24, ** P<0.01, *** P<0.001 compared with tumor-specific CTLs (CD8+MUC1-tetramer+ cells)). (b) IFN-γ releasing T cells from the peripheral blood of breast cancer patients were quantified by IFN-γ ELISPOT (n = 3) and isolated by IFN-γ capture assay (n = 48). (c, d) NKILA expression in IFNγ-releasing CD8+ (upper panel) and CD4+ (lower panel) T cells isolated from the tumor tissues (c) and peripheral blood (d) of breast (n = 48) and lung cancer patients (n = 32) was evaluated by qRT-PCR. ***, P<0.001 compared with non-IFNγ-releasing T cells. (e) Apoptosis of IFNγ-releasing CD8+ and CD4+ T cells isolated from the peripheral blood of breast (n = 5) and lung cancer patients (n = 5) induced by FasL was determined by Annexin V/PI staining and the bars correspond to FasL-induced apoptotic rates (mean ± SEM). ***, P<0.001. (f-h) The isolated tumor-infiltrating TH1 were transduced with shNK1, shNK2, or shvec (f) and the isolated tumor-infitrating TH2 (g) and Treg (h) were transduced with NKILA or vec, followed by co-culturing with autologous tumor cells with or without anti-FasL antibody. Apoptosis of T cells was determined by Annexin V/PI staining and the bars correspond to percentages of Annexin V+ cells (mean ± SEM, n = 3, *P<0.05, **P<0.01, ***P<0.001 compared with the untreated T cell subsets (-); #P<0.05, ##P<0.01, ###P<0.001 compared with the T cell subsets co-cultured with primary tumor cells). P-values, unpaired two-tailed Student’s t-test (a, e-h) and paired two-tailed Student’s t-test (c, d).
Supplementary Figure 7 IFN-γ/STAT1 signaling is activated after TCR stimulation and results in NKILA transcription.
(a) GSEA analysis for the mRNA expression profiles of unactivated (day 0) and activated (day 1) T cells primed by MDA-MB-231 lysate-pulsed DCs from three individual donors revealed the enrichment of STAT1 target genes in the activated CTLs (n = 3 independent experiments). (b) Western blotting was performed for total and phosphorylated JAK and STAT1 in T cells primed by MDA-MB-231 lysate-pulsed DCs (n = 3 independent experiments). (c) Flow cytometry analysis for STAT1 phosphorylation in CD4+ and CD8+ T cells primed by MDA-MB-231 lysate-pulsed DCs (mean ± SEM, n = 3 independent experiments, **, P<0.01; ***, P<0,001 compared with untreated T cells). (d, e) Naive CD4+ and CD8+ T cells were pre-treated with DMSO, JAK1/STAT1 inhibitors Fludarabine (FLU) or Ruxolitinib (RUX), followed by the incubation with anti-CD3 and anti-CD28 in the presence or absence of IFN-γ for 6 days (n=3 independent experiments). (d) NKILA expression of the indicated T cells was evaluated by qRT-PCR (mean ± SEM, n = 3 independent experiments; *, P<0.05; **, P<0.01; ***, P<0.001). (e) Apoptosis of the indicated T cells induced by anti-CD3 was determined by Annexin V/PI staining and the bars correspond to anti-CD3-induced apoptotic rates (mean ± SEM, n = 3 independent experiments; **, P<0.01; ***, P<0.001). (f) STAT1 activities of the T cells primed by MDA-MB-231 lysate-pulsed DCs were examined by EMSA (n = 3 independent experiments). P-values, unpaired two-tailed Student’s t-test (c-e).
Supplementary Figure 8 NKILA expression in activated T cells is finely tuned by calcium-influx-mediated histone acetylation.
CD8+ T cells were primed by the autologous DCs pulsed with MDA-MB-231 lysates and retrieved from the co-cultures on indicated days. (a) A typical CpG island is present at the promoter region of NKILA. Each vertical bar represents a single CpG site. The transcription start site (TSS) is indicated by a curved arrow. BGS analysis confirmed the methylation status of NKILA promoter in T cells primed for 0 day, 1 day and 6 days (n = 3, P>0.05). 8–10 clones were selected for sequencing in one test. Black, deep gray, light gray and white circles represent complete methylation (70.1~100.0%), partial methylation (30.1~70%), little methylation (0.1~30.0%) and unmethylation (0%) on each site, respectively. (b) NKILA expression in the primed CTLs pre-treated with inhibitors for DNA methyltransferase 5’-Aza or DAC was evaluated by qRT-PCR (mean ± SEM, n = 3; P>0.05 compared with DMSO). (c) ChIP analysis for the localization of H3K9me3 and H3K27me3 at NKILA promoter in the primed CTLs (DC group) (mean ± SEM, n = 3, P>0.05 compared with CTLs co-cultured with naive DCs (control group)). (d) NKILA expression in the primed CTLs pre-treated with pan HDAC inhibitors TSA and SAHA, selective Class I HDAC inhibitors MS-275 and FK228, selective Class II HDAC inhibitors TMP269 and Class III HDAC inhibitors droxinostat was evaluated by qRT-PCR (mean ± SEM, n = 3, **P < 0.01, ***P < 0.001 compared with DMSO-treated primed T cells). (e) ChIP analysis for the localization of H4Ac and STAT1 at NKILA promoter in the primed CTLs pre-treated with DMSO, TSA and MS-275 (mean ± SEM, n = 3; *P<0.05; ***P<0.001). (f) ChIP analysis for the localization of HDAC8 and HDAC4, 5 and 7 at NKILA promoter in the primed CTLs (DC group) (mean ± SEM, n = 3; P>0.05 compared with CTLs co-cultured with naive DCs (control group)). (g) Representative images of calmodulin (CaM) nuclear translocation and binding to HDAC1 in the CTLs primed for 1 day, assayed by CaM and HDAC1 double staining and immunofluorescent confocal microscopy (n = 3). Scale bar, 10 μm. (h) Interactions of HDACs (HDAC1, HDAC2, HDAC3) with nuclear-translocated CaM in the nucleus of CTLs upon co-culturing with DCs pulsed with MDA-MB-231 lysates for 1 day were determined by immunoprecipitation (n = 3). (i) The scheme of the mechanism underlying NKILA expression and contribution to the sensitivity to AICD of different T cell subsets. P-values, unpaired two-tailed Student’s t-test (a-f).
Supplementary information
Supplementary Figures
Supplementary Figures 1–8
Supplementary Text
Supplementary Tables 1–5
Supplementary Dataset 1
Uncropped gels
Rights and permissions
About this article

Cite this article
Huang, D., Chen, J., Yang, L. et al. NKILA lncRNA promotes tumor immune evasion by sensitizing T cells to activation-induced cell death. Nat Immunol 19, 1112–1125 (2018). https://doi.org/10.1038/s41590-018-0207-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-018-0207-y
This article is cited by
-
Identification of breast cancer subgroups and immune characterization based on glutamine metabolism-related genes
BMC Medical Genomics (2024)
-
Tumour circular RNAs elicit anti-tumour immunity by encoding cryptic peptides
Nature (2024)
-
Targeting and engineering long non-coding RNAs for cancer therapy
Nature Reviews Genetics (2024)
-
Mutant KRAS-activated circATXN7 fosters tumor immunoescape by sensitizing tumor-specific T cells to activation-induced cell death
Nature Communications (2024)
-
Long non-coding RNAs in cancer: multifaceted roles and potential targets for immunotherapy
Molecular and Cellular Biochemistry (2024)
