
Abstract
Reactive oxygen species (ROS) are generated by virus-infected cells; however, the physiological importance of ROS generated under these conditions is unclear. Here we found that the inflammation and cell death induced by exposure of mice or cells to sources of ROS were not altered in the absence of canonical ROS-sensing pathways or known cell-death pathways. ROS-induced cell-death signaling involved interactions among the cellular ROS sensor and antioxidant factor KEAP1, the phosphatase PGAM5 and the proapoptotic factor AIFM1. Pgam5 –/– mice showed exacerbated lung inflammation and proinflammatory cytokines in an ozone-exposure model. Similarly, challenge with influenza A virus led to increased infiltration of the virus, lymphocytic bronchiolitis and reduced survival of Pgam5 –/– mice. This pathway, which we have called ‘oxeiptosis’, was a ROS-sensitive, caspase independent, non-inflammatory cell-death pathway and was important for protection against inflammation induced by ROS or ROS-generating agents such as viral pathogens.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
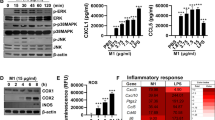
References
Sauer, H., Wartenberg, M. & Hescheler, J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell. Physiol. Biochem. 11, 173–186 (2001).
Nakamura, H., Nakamura, K. & Yodoi, J. Redox regulation of cellular activation. Annu. Rev. Immunol. 15, 351–369 (1997).
Sandalio, L. M., Rodríguez-Serrano, M., Romero-Puertas, M. C. & del Río, L. A. Role of peroxisomes as a source of reactive oxygen species (ROS) signaling molecules. Subcell. Biochem. 69, 231–255 (2013).
Rosca, M. G. et al. Oxidation of fatty acids is the source of increased mitochondrial reactive oxygen species production in kidney cortical tubules in early diabetes. Diabetes 61, 2074–2083 (2012).
Lü, J.-M., Lin, P. H., Yao, Q. & Chen, C. Chemical and molecular mechanisms of antioxidants: experimental approaches and model systems. J. Cell. Mol. Med. 14, 840–860 (2010).
Taguchi, K., Motohashi, H. & Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes to Cells 16, 123–140 (2011).
Song, J. J. et al. Role of glutaredoxin in metabolic oxidative stress. Glutaredoxin as a sensor of oxidative stress mediated by H2O2. J. Biol. Chem. 277, 46566–46575 (2002).
Zhang, D. D. & Hannink, M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol. Cell. Biol. 23, 8137–8151 (2003).
Bryan, H. K., Olayanju, A., Goldring, C. E. & Park, B. K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 85, 705–717 (2013).
Kaspar, J. W., Niture, S. K. & Jaiswal, A. K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 47, 1304–1309 (2009).
Circu, M. L. & Aw, T. Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 48, 749–762 (2010).
Yang, D. et al. Pro-inflammatory cytokines increase reactive oxygen species through mitochondria and NADPH oxidase in cultured RPE cells. Exp. Eye Res. 85, 462–472 (2007).
Hroudová, J., Singh, N. & Fišar, Z. Mitochondrial dysfunctions in neurodegenerative diseases: relevance to Alzheimer’s disease. BioMed. Res. Int. 2014, 175062 (2014).
Ano, Y. et al. Oxidative damage to neurons caused by the induction of microglial NADPH oxidase in encephalomyocarditis virus infection. Neurosci. Lett. 469, 39–43 (2010).
Li, Y., Boehning, D. F., Qian, T., Popov, V. L. & Weinman, S. A. Hepatitis C virus core protein increases mitochondrial ROS production by stimulation of Ca2+ uniporter activity. FASEB J. 21, 2474–2485 (2007).
Bhattacharya, A. et al. Superoxide dismutase 1 protects hepatocytes from type I interferon-driven oxidative damage. Immunity 43, 974–986 (2015).
Olagnier, D. et al. Cellular oxidative stress response controls the antiviral and apoptotic programs in dengue virus-infected dendritic cells. PLoS Pathog. 10, e1004566 (2014).
Bottero, V., Chakraborty, S. & Chandran, B. Reactive oxygen species are induced by Kaposi’s sarcoma-associated herpesvirus early during primary infection of endothelial cells to promote virus entry. J. Virol. 87, 1733–1749 (2013).
Edwards, M. R. et al. The Marburg virus VP24 protein interacts with Keap1 to activate the cytoprotective antioxidant response pathway. Cell Reports 6, 1017–1025 (2014).
Johnson, B. et al. Dimerization controls Marburg virus VP24-dependent modulation of host antioxidative stress responses. J. Mol. Biol. 428, 3483–3494 (2016).
Amri, F., Ghouili, I., Amri, M., Carrier, A. & Masmoudi-Kouki, O. Neuroglobin protects astroglial cells from hydrogen peroxide-induced oxidative stress and apoptotic cell death. J. Neurochem. 140, 151–169 (2017).
Kim, J.-Y. & Park, J.-H. ROS-dependent caspase-9 activation in hypoxic cell death. FEBS Lett. 549, 94–98 (2003).
Radogna, F. et al. Cell type-dependent ROS and mitophagy response leads to apoptosis or necroptosis in neuroblastoma. Oncogene 35, 3839–3853 (2016).
Ravindran, J., Gupta, N., Agrawal, M., Bala Bhaskar, A. S. & Lakshmana Rao, P. Modulation of ROS/MAPK signaling pathways by okadaic acid leads to cell death via, mitochondrial mediated caspase-dependent mechanism. Apoptosis 16, 145–161 (2011).
Shen, C. et al. Aldehyde dehydrogenase 2 deficiency negates chronic low-to-moderate alcohol consumption-induced cardioprotecion possibly via ROS-dependent apoptosis and RIP1/RIP3/MLKL-mediated necroptosis. Biochim. Biophys. Acta 1863, 1912–1918 (2017).
Wiegman, C. H. et al. Oxidative stress-induced mitochondrial dysfunction drives inflammation and airway smooth muscle remodeling in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 136, 769–780 (2015).
Triantaphyllopoulos, K. et al. A model of chronic inflammation and pulmonary emphysema after multiple ozone exposures in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 300, L691–L700 (2011).
Kosmider, B., Loader, J. E., Murphy, R. C. & Mason, R. J. Apoptosis induced by ozone and oxysterols in human alveolar epithelial cells. Free Radic. Biol. Med. 48, 1513–1524 (2010).
Wang, Z., Jiang, H., Chen, S., Du, F. & Wang, X. The mitochondrial phosphatase PGAM5 functions at the convergence point of multiple necrotic death pathways. Cell 148, 228–243 (2012).
Kobayashi, A. et al. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol. Cell. Biol. 24, 7130–7139 (2004).
Lo, S.-C. & Hannink, M. PGAM5, a Bcl-XL-interacting protein, is a novel substrate for the redox-regulated Keap1-dependent ubiquitin ligase complex. J. Biol. Chem. 281, 37893–37903 (2006).
Lo, S.-C. & Hannink, M. PGAM5 tethers a ternary complex containing Keap1 and Nrf2 to mitochondria. Exp. Cell Res. 314, 1789–1803 (2008).
Ceh-Pavia, E., Spiller, M. P. & Lu, H. Folding and biogenesis of mitochondrial small Tim proteins. Int. J. Mol. Sci. 14, 16685–16705 (2013).
Susin, S. A. et al. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature 397, 441–446 (1999).
Joza, N. et al. AIF: not just an apoptosis-inducing factor. Ann. NY Acad. Sci. 1171, 2–11 (2009).
Wilkins, J. M., McConnell, C., Tipton, P. A. & Hannink, M. A conserved motif mediates both multimer formation and allosteric activation of phosphoglycerate mutase 5. J. Biol. Chem. 289, 25137–25148 (2014).
Pichlmair, A. et al. Viral immune modulators perturb the human molecular network by common and unique strategies. Nature 487, 486–490 (2012).
Page, A. et al. Marburgvirus hijacks nrf2-dependent pathway by targeting nrf2-negative regulatorkeap1. Cell Rep. 6, 1026–1036 (2014).
Medvedev, R., Ploen, D. & Hildt, E. HCV and oxidative stress: implications for HCV life cycle and HCV-associated pathogenesis. Oxid. Med. Cell. Longev. 2016, 9012580 (2016).
Ren, J.-H. et al. Protective Role of Sirtuin3 (SIRT3) in oxidative stress mediated by hepatitis B virus X protein expression. PLoS One 11, e0150961 (2016).
Kajitani, N. et al. Mechanism of A23187-induced apoptosis in HL-60 cells: dependency on mitochondrial permeability transition but not on NADPH oxidase. Biosci. Biotechnol. Biochem. 71, 2701–2711 (2007).
Blaser, H., Dostert, C., Mak, T. W. & Brenner, D. TNF and ROS crosstalk in inflammation. Trends Cell Biol. 26, 249–261 (2016).
Chacinska, A. et al. Essential role of Mia40 in import and assembly of mitochondrial intermembrane space proteins. EMBO J. 23, 3735–3746 (2004).
Shindo, R., Kakehashi, H., Okumura, K., Kumagai, Y. & Nakano, H. Critical contribution of oxidative stress to TNFα-induced necroptosis downstream of RIPK1 activation. Biochem. Biophys. Res. Commun. 436, 212–216 (2013).
Dixon, S. J. et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 149, 1060–1072 (2012).
Moriwaki, K. et al. The mitochondrial phosphatase PGAM5 is dispensable for necroptosis but promotes inflammasome activation in macrophages. J. Immunol. 196, 407–415 (2016).
Szatrowski, T. P. & Nathan, C. F. Production of large amounts of hydrogen peroxide by human tumor cells. Cancer Res. 51, 794–798 (1991).
Marconett, C. N. et al. BZL101, a phytochemical extract from the Scutellaria barbata plant, disrupts proliferation of human breast and prostate cancer cells through distinct mechanisms dependent on the cancer cell phenotype. Cancer Biol. Ther. 10, 397–405 (2010).
Wang, R. et al. Hypermethylation of the Keap1 gene in human lung cancer cell lines and lung cancer tissues. Biochem. Biophys. Res. Commun. 373, 151–154 (2008).
Nogusa, S. et al. RIPK3 activates parallel pathways of MLKL-driven necroptosis and fadd-mediated apoptosis to protect against influenza A virus. Cell Host Microbe 20, 13–24 (2016).
Habjan, M. et al. Sequestration by IFIT1 impairs translation of 2’O-unmethylated capped RNA. PLoS Pathog. 9, e1003663 (2013).
Gabriel, G. et al. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. USA 102, 18590–18595 (2005).
Provencher, S. W. & Glöckner, J. Estimation of globular protein secondary structure from circular dichroism. Biochemistry 20, 33–37 (1981).
Cox, J. & Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 26, 1367–1372 (2008).
Cox, J. et al. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteomics 13, 2513–2526 (2014).
Crooks, G. E., Hon, G., Chandonia, J.-M. & Brenner, S. E. WebLogo: a sequence logo generator. Genome Res. 14, 1188–1190 (2004).
Besnard, A.-G. et al. Dual Role of IL-22 in allergic airway inflammation and its cross-talk with IL-17A. Am. J. Respir. Crit. Care Med. 183, 1153–1163 (2011).
Acknowledgements
We thank F. Weber (Veterinary University Giessen) for pI.18_3xFlag_NSs and pI.18_3xFlag_DMx_1xFlag; F. Meissner (Max-Planck Institute of Biochemistry, Munich) for Jurkat cells; R. Klein (Max-Planck Institute of Neurobiology, Munich) for SKN-BE2 cells; A. Bowie (Trinity College Dublin) for HEK293 cells; A. Musacchio (Max-Planck Institute of Cell Biology, Dresden) for HeLa FlpIn cells; G. Kochs (University of Freiburg) for antibody to FluAV NS1; A. Chaikuad and S. Knapp (University of Oxford) for amino-terminal His-tagged PGAM5 (pNIC28-Bsa4-PGAM5(Δ2–28); the innate immunity laboratory for critical discussions and suggestions; A. Mann for technical assistance; the MPI-B core facility for technical assistance with protein purification, analysis and imaging; M. Wilson and ECM Biosciences for raising the antibody to AIFM1 phosphorylated at Ser116; K. Mayr, I. Paron and G. Sowa for maintaining mass spectrometers; and S. Kaphengst and the MPI-B animal facility for breeding mice. Supported by the Max-Planck Free Floater program (A.P.), the German research foundation (PI 1084/2 and TRR179 to A.P. and SFB 1160, project 13) to P.S.), the European Research Council (starting grant ERC-StG iVIP, 311339 to A.P.), Infect-Era and the German Federal Ministry of Education and Research (ERASE) (A.P.), the US National Institutes of Health (R01 AI107056 and 1R41AI127009 (Moir) to D.W.L and G.K.A.), the Centre National de la Recherche Scientifique (B.R.) and the European Regional Development Fund (BIO-TARGET 2016-00110366 to Region Centre Val de Loire and la Région Centre (Respir_Ozone, 2014-00091905) (Valerie Quesniaux), to B.R.).
Author information
Authors and Affiliations
Contributions
C.H., Ch.M., Cl.M., D.A.H., F.L.P., D.S., J.W., M.B. and P.S. conducted the experiments; C.B. performed structural modelling; C.H., D.A.H. and P.H. analyzed data; D.W.L and G.K.A. contributed reagents; and C.H., D.W.L, G.K.A., F.P., P.S., B.R. and A.P. designed the experiments and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Ozone- and H2O2-exposure model: testing of cell-death pathways
(a-b) C57BL/6 mice (wt) or Nlrp3 –/– , Casp1/11 –/– and Asc –/– mice were exposed to 1 ppm of ozone for 1h and BAL analyzed for protein content and for myeloperoxidase (MPO) 24h later. Dots indicate individual mice. Graph shows average +/- SD, two-sided t-test, ns: non-significant. (c) Electron microscopy analysis of H2O2 treated MEFs. Representative micrographs show MEFs wt treated for 6h with 1 mM H2O2 with or without pre-treatment with Z-VAD. Left panel shows 5000x magnification, right panel shows 30000x magnifications. One representative experiment of three is shown. (d) Comparison of cell viability assays of HeLa cells after hydrogen peroxide (H2O2) treatment. Cells were treated for 21h with 0.5 mM H2O2. Cell titers were determined by CellTiter-Glo, MTT and resazurin-based cell viability assay. The plot shows the mean ± S.D. of six individual treatments. (e) LDH-assay to determine cell viability of HeLa cells after hydrogen peroxide (H2O2) treatment. Cells were treated 21h with 0.5 mM H2O2. Release of active LDH was determined by LDH-assay. The plot shows the mean ± S.D. of six individual treatments. One representative experiment of four is shown. ** p-value < 0.0001, two-sided t-test. (f) Viability of Jurkat cells after hydrogen peroxide (H2O2) treatment. Cells were left untreated or treated with 20 µM Z-VAD-FMK (Z-VAD) for 1h, followed by 21h treatment with 0.5 mM H2O2. Cell titers were determined by resazurin-based cell viability assay. The plot shows the mean ± S.D. of six individual treatments. One representative experiment of three is shown. * p-value < 0.001, two-sided t-test. (g) Immunoblot analysis of HeLa cells and MEFs treated with autophagy inducer Carbonyl cyanide m-chlorophenylhydrazone (CCCP) and H2O2 as comparison for expression levels of LC3-A II and Atg7. One experiment of three is shown. (h) HeLa cells were left untreated or treated with 1 µM Ferrostatin-1 (Fer-1) for 1h, followed by 21h treatment with 0.5 mM H2O2 or 20 µM ferroptosis-inducer Sorafenib. The plot shows the mean release of active LDH ± S.D. of six individual treatments. One representative experiment of three is shown. * p-value < 0.001, 2way ANOVA. (i) Cells were left untreated or treated with indicated concentrations of Ferrostatin-1 for 1h, followed by 21h treatment with 0.5 mM H2O2. Cell titers were determined by resazurin-based cell viability assay. The plot shows the mean ± S.D. of six individual treatments. One representative experiment of two is shown. (j) Expression of NRF2 regulated target genes after siRNA mediated knockdown of KEAP1, KEAP1 and NRF or siScr in HeLa cells 48h after siRNA treatment.
Supplementary Figure 2 KEAP1 interacts with PGAM5, which is required for H2O2-mediated cell death
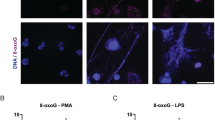
(a) Binding of endogenous KEAP1 to PGAM5 deletion mutants. Expression of SII-HA-tagged PGAM5 wild-type (wt) and deletion mutants lacking MLS and TM domain (Δ 2-29), lacking MLS, TM domain and phosphatase activity modulating region (PAMR) (Δ 2-69) and lacking MLS, TM domain, PAMR and KEAP1 binding site (Δ2-89) in HEK293T cells. Immunoblot analysis of input lysates and SII-AP. One of two experiments with similar results are shown. (b) Representative confocal images of HeLa cells left untreated or treated with 0.5 mM H2O2 for 12h and stained for DAPI (blue), the mitochondrial marker COX IV (green) and PGAM5 or KEAP1 (red), respectively. Overlays are shown in yellow. One representative experiment of six is shown. (c) Viability of HeLa cells treated with siRNA against PGAM5 and siScr after 21h H2O2 treatment. Cell viability was determined by resazurin-conversion assay. The plot shows the mean ± S.D. of four individual treatments. One representative experiment of two is shown. (d) as (c) but different siRNAs targeting PGAM5 were used. (e) as (c) but cells were pre-treated with 20 µM Z-VAD-FMK (Z-VAD) for 1h before 21h treatment with 0.5 mM H2O2. * p < 0.01, ** p < 0.001, ns: non-significant, 2way ANOVA with Bonferroni post t-test. (f) Characterization of Pgam5 knockout MEFs by genotyping (PCR) (bottom) and quantification of Pgam5 mRNA levels by RT-qPCR (top) normalized to hydroxymethylbilane synthase mRNA (Hmbs). Histogram shows mean fold change of analyses done in triplicates. N.D.: not detectable, one representative experiment of three is shown (g) MEFs of the indicated genotype were treated with the indicated concentrations of Sorafenib (Sora) for 20h and viability was tested by resazurin-conversion assay. The graph shows mean activity ± S.D. of six measurements. One representative experiment of two is shown.
Supplementary Figure 3 PGAM5 interacts with AIFM1, which is involved in H2O2.mediated cell death
(a) AP of Renilla tagged (-Ren) AIFM1 mutant proteins with SII-HA-PGAM5. HEK293T cells were co-transfected with SII-HA-tagged PGAM5 and Renilla-tagged AIFM1 mutants or Renilla-tagged-control (THYN1; ctrl). After 24h Renilla activity was measured in cell lysate and SII precipitates. Immunoblot shows expression of bait proteins in cell lysates. One representative experiment of five with similar results is shown. (b) HeLa cells were treated with siRNAs against AIFM1 and siScr exposed 0.5 mM H2O2 for 21h. Cell viability was determined by resazurin assay. The plot shows the mean ± S.D. of four individual treatments. One representative experiment of two is shown. (c) As (b) but siRNA against AIFM1 and PGAM5 was transfected in SKN-BE2 cells. (d) Viability of HeLa cells treated with indicated siRNA combinations against KEAP1, AIFM1 and PGAM5 or siScr after 21h H2O2 treatment. Cell titers were determined by resazurin-based viability assay. (c), (d) The plot shows the mean ± S.D. of six individual treatments. One representative experiment of two is shown. * p-value < 0.01, ** p-value < 0.001, ns: non-significant, 1way ANOVA with Bonferroni post t-test. (e) HeLa cells were treated for 5h with the indicated amounts of H2O2 followed by measuring mitochondrial respiration. Addition of 1.5 µM Oligomycin (OligoA), 1 µM Carbonyl cyanide m-chlorophenyl hydrazone (CCCP), 4 µM Antimycin A (AA) and 2 µM Rotenone is indicated. The graph shows average oxygen consumption rate (OCR) ± S.D. of three measurements.
Supplementary Figure 4 PGAM5–AIFM1 interaction and PGAM5’s phosphatase activity
(a) Sequence alignment of AIFM1 amino acids 109-122 of the indicated species. (b) Selectivity of AIFM1 pS116 antibody. HeLa cell lysates were treated for 15 min with or without calf intestine alkaline phosphatase (CIAP) and subjected to Immunoblot stained for AIFM1 pS116 and AIFM1. One representative experiment of three with similar results is shown. (c) Surface charge potential of PGAM5 dimer crystal structure (PDB 3MXO). (d) Zoom in on dimer interphase interface of PGAM5. One monomer is highlighted in black, the other in grey. Left panel shows the wild-type (wt) protein, right panel shows the modeled mutant PGAM5[F244D]. Side chains of phenylalanine (F) 244 and, after mutation, aspartic acid (D) 244 are highlighted in purple (carbon) and red (oxygen). (e) Binding of Ren-PGAM5, Ren-PGAM5[F244D] or Ren-ctrl (THYN1) to SII-HA-PGAM5 in HEK293T cells. Renilla assay of cell lysates or SII precipitates. (f) Phosphatase activity of PGAM5 and PGAM5 mutants carrying an N-terminal (N) or C-terminal (C) tag precipitated from cell lysates. The plot shows the mean ± S.D. of one AP with three independent measurements. One representative experiment of three is shown. (g) Circular dichroism (CD) spectroscopy of PGAM5Δ2-28 and PGAM5Δ2-28[F244D] to test comparability of secondary structures and overall integrity of both recombinant proteins. (h) Size exclusion chromatography analysis of PGAM5Δ2-28 wt and [F244D] mutant for dimer and monomer formation. (i) Phosphatase activity of 0.1 µg recombinant PGAM5Δ2-28 wt and [F244D] mutant. Where indicated phosphatase inhibitor 1 mM orthovanatade (Van) has been added to the reaction mix. The plot shows the mean ± S.D. of three independent measurements. One representative experiment of three is shown. * p-value < 0.001 2way ANOVA.
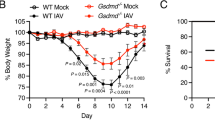
Supplementary Figure 5 Exposure of Pgam5 –/– mice to ozone
Pgam5 +/+ and Pgam5 –/– mice (n = 3-6 per condition) were treated 1 ppm ozone for 1h and sacrificed after 24h recovery time. (a) Determination of total proteins in BAL. (b) Total cells and neutrophils in BAL. (a), (b) * p-value < 0.001 by 1way Anova and Bonferroni post t-test. Graphs show individual mice (dots), mean +/- SEM. One representative experiment of two is shown.
Supplementary Figure 6 Targeting and activation of oxeiptosis during viral infection
(a) AP-LC-MS/MS experiments using SII-HA-tagged K3 of Karposi Sarcoma Herpes virus (KSHV) and SII-HA-THYN1 (ctrl) as baits. Volcano plots show the average degrees of enrichment (ratio of label-free quantitation (LFQ) protein intensities; x-axis) and p-value (two-tailed t-test; y-axis) for each identified protein. Significantly enriched proteins (FDR: 0.01, S0=1) are separated from background proteins by a hyperbolic curve (dotted line). Bait is marked in red, PGAM5 is highlighted in green. Four independent APs were performed for all baits. (b) As in (a) but Immunoblot for endogenous PGAM5 and HA tagged bait proteins was performed. (c) Ren-K3, Ren-K5 or Ren-ctrl were co-transfected with SII-HA-PGAM5 into HEK293T cells and used for Immunoblot analysis or Renilla activity measurement in input lysate or after SII precipitation. Graphs show mean +/- SD of triplicate measurements. (d) HEK293T cells were co-transfected with SII-HA-tagged PGAM5mutant construct and Renilla-tagged K3 or Renilla-tagged-ctrl (THYN1). After 24h Renilla activity was measured in lysate and SII precipitates. Immunoblot shows expression of bait proteins in cell lysates. (e) AP of Ren-AIFM1 using Flag tagged non-structural protein S (NSs) of LaCV or Flag-(ctrl) (ΔMx) as baits followed by Renilla assay. (f) AP of endogenous AIFM1 in HEK cells using transfected Flag-NSs or Flag-ctrl (ΔMx) as baits followed by Immunoblot analysis. (g) As in (a) but with non-structural protein 2 (NS2) of RSV or non-expressing controls, AIFM1 is highlighted in green. (h) AP of Ren-NS2 (RSV) and Ren-ctrl (THYN1) using SII-HA-AIFM1 as bait followed by Renilla assay. Graph shows mean +/− SD of triplicate (input) or quadruplicate (AP) measurements. (i) Interaction of SII-HA-AIFM1 and AIFM1 mutant proteins and SII-HA-ctrl (THYN1) with co-expressed Ren-NS2 in HEK293T cells. Renilla activity was tested in cell lysates and after SII precipitation. (j) Influence of FluAV infection on NRF2 regulated gene transcription in HeLa cells tested by RT-qPCR normalized to mRNA of TATA-binding protein (TBP). (k) Viability of HeLa cells treated with siRNAs against PGAM5 or control (siSCR) and infected with the indicated MOI of FluAV for 40h. Histograms show average resazurin activity of six measurements +/− SD. One representative experiment of three is shown, ** p-value < 0.01, 2way ANOVA with Bonferroni post t-test. (l) Flu NS1 and (m) IL-6 mRNA levels in lung tissue of Pgam5 +/– and Pgam5 –/– mice that were infected with FluAV for five days (n = 10) and uninfected wt mice (n = 2). * p-value < 0.05, two-sided t-test. (n) CXCL1 and CCL2 accumulation in BAL of Pgam5 +/– and Pgam5 –/– mice that were infected with FluAV for five days and uninfected wt mice. (d-g), (i) Histograms show mean +/-SD of triplicate measurements. * p-value < 0.05, *** p-value < 0.001, t-test. One representative experiment of two (i), three (d), (e), (f), (h) or four (b), (c) is shown.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1-6 and Supplementary Tables 1-4
Rights and permissions
About this article

Cite this article
Holze, C., Michaudel, C., Mackowiak, C. et al. Oxeiptosis, a ROS-induced caspase-independent apoptosis-like cell-death pathway. Nat Immunol 19, 130–140 (2018). https://doi.org/10.1038/s41590-017-0013-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-017-0013-y
This article is cited by
-
Pan-cancer evaluation of regulated cell death to predict overall survival and immune checkpoint inhibitor response
npj Precision Oncology (2024)
-
Functional genomic screens with death rate analyses reveal mechanisms of drug action
Nature Chemical Biology (2024)
-
The E3 ubiquitin ligase MARCH2 protects against myocardial ischemia-reperfusion injury through inhibiting pyroptosis via negative regulation of PGAM5/MAVS/NLRP3 axis
Cell Discovery (2024)
-
An apoptosis-inducing factor controls programmed cell death and laccase expression during fungal interactions
Applied Microbiology and Biotechnology (2024)
-
Exploring Therapeutic Approaches for Vitiligo: An Inclusive Review from Translational Modalities to Alternative Therapies
Current Dermatology Reports (2024)
