
Abstract
N6-methyladenosine (6mA) is a DNA modification that has recently been found to play regulatory roles during mammalian early embryo development and mitochondrial transcription. We found that a dioxygenase CcTet from the fungus Coprinopsis cinerea is also a dsDNA 6mA demethylase. It oxidizes 6mA to the intermediate N6-hydroxymethyladenosine (6hmA) with robust activity of 6mA-containing duplex DNA (dsDNA) as well as isolated genomics DNA. Structural characterization revealed that CcTet utilizes three flexible loop regions and two key residues—D337 and G331—in the active pocket to preferentially recognize substrates on dsDNA. A CcTet D337F mutant protein retained the catalytic activity on 6mA but lost activity on 5-methylcytosine. Our findings uncovered a 6mA demethylase that works on dsDNA, suggesting potential 6mA demethylation in fungi and elucidating 6mA recognition and the catalytic mechanism of CcTet. The CcTet D337F mutant protein also provides a chemical biology tool for future functional manipulation of DNA 6mA in vivo.

This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
The coordinates of the crystal structure have been deposited with the Protein Data Bank under the accession nos. 7VPN and 7W5P. All structures cited in this publication are available under accession nos. 4NM6, 5CG8, 5CG9, 5ZMD, 7CY8, 4JHT, 6IMC, 6KSF, 3BUC, 3BKZ, 2IUW, 4NRO, 4QKD, 3THT and 3LFM. Source data are provided with this paper.
References
Luo, G.-Z., Blanco, M. A., Greer, E. L., He, C. & Shi, Y. DNA N6-methyladenine: a new epigenetic mark in eukaryotes? Nat. Rev. Mol. Cell Biol. 16, 705–710 (2015).
Wion, D. & Casadesús, J. N6-methyl-adenine: an epigenetic signal for DNA–protein interactions. Nat. Rev. Microbiol. 4, 183–192 (2006).
Fu, Y. et al. N6-methyldeoxyadenosine marks active transcription start sites in Chlamydomonas. Cell 161, 879–892 (2015).
Beh, L. Y. et al. Identification of a DNA N6-adenine methyltransferase complex and its impact on chromatin organization. Cell 177, 1781–1796.e25 (2019).
Luo, G.Z. et al. N6-methyldeoxyadenosine directs nucleosome positioning in Tetrahymena DNA. Genome Biol. 19, 200 (2018).
Greer, E. L. et al. DNA methylation on N6-adenine in C. elegans. Cell 161, 868–878 (2015).
Mondo, S. J. et al. Widespread adenine N6-methylation of active genes in fungi. Nat. Genet. 49, 964–968 (2017).
Zhang, G. et al. N6-methyladenine DNA modification in Drosophila. Cell 161, 893–906 (2015).
Zhou, C. et al. Identification and analysis of adenine N6-methylation sites in the rice genome. Nat. Plants 4, 554–563 (2018).
Liang, Z. et al. DNA N6-adenine methylation in Arabidopsis thaliana. Dev. Cell 45, 406–416.e3 (2018).
Luo, G.-Z. & He, C. DNA N6-methyladenine in metazoans: functional epigenetic mark or bystander? Nat. Struct. Mol. Biol. 24, 503–506 (2017).
Liu, J. et al. Abundant DNA 6mA methylation during early embryogenesis of zebrafish and pig. Nat. Commun. 7, 13052 (2016).
Wu, T. P. et al. DNA methylation on N6-adenine in mammalian embryonic stem cells. Nature 532, 329–333 (2016).
Yao, B. et al. DNA N6-methyladenine is dynamically regulated in the mouse brain following environmental stress. Nat. Commun. 8, 1122 (2017).
Xiao, C.-L. et al. N6-methyladenine DNA modification in the human genome. Mol. Cell 71, 306–318.e7 (2018).
O’Brown, Z. K. et al. Sources of artifact in measurements of 6mA and 4mC abundance in eukaryotic genomic DNA. BMC Genomics 20, 445 (2019).
Douvlataniotis, K., Bensberg, M., Lentini, A., Gylemo, B. & Nestor, C. E. No evidence for DNA N6-methyladenine in mammals. Sci. Adv. 6, eaay3335 (2020).
Kong, Y. et al. Critical assessment of DNA adenine methylation in eukaryotes using quantitative deconvolution. Science 375, 515–522 (2022).
Hao, Z. et al. N6-deoxyadenosine methylation in mammalian mitochondrial DNA. Mol. Cell 78, 382–395.e8 (2020).
Li, Z. et al. N6-methyladenine in DNA antagonizes SATB1 in early development. Nature 583, 625–630 (2020).
Jia, G. et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 7, 885–887 (2011).
Zheng, G. et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 49, 18–29 (2013).
Yao, B. et al. Active N6-methyladenine demethylation by DMAD regulates gene expression by coordinating with polycomb protein in neurons. Mol. Cell 71, 848–857.e6 (2018).
Zhang, M. et al. Mammalian ALKBH1 serves as an N6-mA demethylase of unpairing DNA. Cell Res. 30, 197–210 (2020).
Tian, L.-F. et al. Structural basis of nucleic acid recognition and 6mA demethylation by human ALKBH1. Cell Res. 30, 272–275 (2020).
Kweon, S.-M. et al. An adversarial DNA N6-methyladenine-sensor network preserves polycomb silencing. Mol. Cell 74, 1138–1147.e6 (2019).
Yi, C., Yang, C.-G. & He, C. A non-heme iron-mediated chemical demethylation in DNA and RNA. Acc. Chem. Res. 42, 519–529 (2009).
Fu, Y. et al. FTO-mediated formation of N6-hydroxymethyladenosine and N6-formyladenosine in mammalian RNA. Nat. Commun. 4, 1798 (2013).
Toh, J. D. W. et al. Distinct RNA N-demethylation pathways catalyzed by nonheme iron ALKBH5 and FTO enzymes enable regulation of formaldehyde release rates. Proc. Natl Acad. Sci. USA 117, 25284–25292 (2020).
Qin, F. X., Jiang, C.-Y., Jiang, T. & Cheng, G. New targets for controlling Ebola virus disease. Natl Sci. Rev. 2, 266–267 (2015).
Tahiliani, M. et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324, 930–935 (2009).
Ito, S. et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 333, 1300–1303 (2011).
He, Y.-F. et al. Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science 333, 1303–1307 (2011).
Hu, L. et al. Crystal structure of TET2-DNA complex: insight into TET-mediated 5mC oxidation. Cell 155, 1545–1555 (2013).
Hu, L. et al. Structural insight into substrate preference for TET-mediated oxidation. Nature 527, 118–122 (2015).
Xue, J.-H. et al. A vitamin-C-derived DNA modification catalysed by an algal TET homologue. Nature 569, 581–585 (2019).
Li, W. et al. Molecular mechanism for vitamin C-derived C5-glyceryl-methylcytosine DNA modification catalyzed by algal TET homologue CMD1. Nat. Commun. 12, 744 (2021).
Zhang, L. et al. A TET homologue protein from Coprinopsis cinerea (CcTet) that biochemically converts 5-methylcytosine to 5-hydroxymethylcytosine, 5-formylcytosine, and 5-carboxylcytosine. J. Am. Chem. Soc. 136, 4801–4804 (2014).
Zhang, L. et al. Thymine DNA glycosylase specifically recognizes 5-carboxylcytosine-modified DNA. Nat. Chem. Biol. 8, 328–330 (2012).
Zhu, C. & Yi, C. Switching demethylation activities between AlkB family RNA/DNA demethylases through exchange of active-site residues. Angew. Chem. Int. Ed. Engl. 53, 3659–3662 (2014).
Zhang, X. et al. Structural insights into FTO’s catalytic mechanism for the demethylation of multiple RNA substrates. Proc. Natl Acad. Sci. USA 116, 2919–2924 (2019).
Vanyushin, B. F., Tkacheva, S. G. & Belozersky, A. N. Rare bases in animal DNA. Nature 225, 948–949 (1970).
Yu, M. et al. Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome. Cell 149, 1368–1380 (2012).
Minor, W., Cymborowski, M., Otwinowski, Z. & Chruszcz, M. HKL-3000: the integration of data reduction and structure solution—from diffraction images to an initial model in minutes. Acta Crystallogr. D Biol. Crystallogr. 62, 859–866 (2006).
Adams, P. D. et al. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 58, 1948–1954 (2002).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
Acknowledgements
This project was supported by grants from the Major Research plan of the National Natural Science Foundation of China (no. 91853118 to Liang Zhang), NSFC (nos. 22107067 to Lin Zhang; 22077081 and 21722802 to Liang Zhang), Science and Technology Commission of Shanghai Municipality (nos. 20S11900300 and 22S11900600 to Liang Zhang), Shuguang Program supported by the Shanghai Education Development Foundation and Shanghai Municipal Education Commission (no. 20SG16 to Liang Zhang), innovative research team of high-level local universities in Shanghai (no. SSMU-ZLCX20180702 to Liang Zhang) and the Key Program of NSFC (no. 22137006 to H.-W.L.). We thank Professor Zhonghua Liu from Nanjing Normal University for generously providing the genomic DNA of C. cinerea. We thank the staff from the BL19U1 and BL18U beamlines of the NFPS in Shanghai at the SSRF for their assistance during data collection. We thank the staff from the Core Facility of Basic Medical Sciences at Shanghai Jiao Tong University School of Medicine for their assistance during the LC–MS/MS data collection. C.H. is a Howard Hughes Medical Institute Investigator.
Author information
Authors and Affiliations
Contributions
Liang Zhang and Lin Zhang designed the experiments. Y.M. and J.Z. performed the protein purification and crystallization. Lin Zhang and J.H. performed the LC–MS/MS-based activity assays and determined the kinetics. Lin Zhang performed the biophysical experiments. Liang Zhang, C.H., Lin Zhang, H.-W.L. and H.-Z.C. wrote the paper. All authors discussed and commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
C.H. is a scientific founder and member of the scientific advisory board of Accent Therapeutics and Inferna Green. The other authors declare no competing interests.
Peer review
Peer review information
Nature Chemical Biology thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 In vitro enzymatic activity of human TET2 catalytic domain on 6 mA or 5mC containing 19 bp dsDNA.
N = 3 biologically independent experiments. Data represent mean values ± s.d.
Extended Data Fig. 2 LC-MS/MS analysis of 6 mA demethylation catalyzed by overexpression of CcTet wild type and CcTet D337F in E. coli. in vivo.
The unpaired two-sided student’s t-test values were also labeled. P = 0.002 (CcTet), 0.000083 (CcTet D337F), **P < 0.01, ****P < 0.0001. N = 6 biologically independent experiments. Data represent mean values ± s.d.
Extended Data Fig. 3 Multiple sequence alignment of TETs and ALKBs family.
The structural comparison was generated in the DALI server (http://ekhidna2.biocenter.helsinki.fi/dali/), and the sequence alignment was generated through ESPript online server (https://espript.ibcp.fr/ESPript/ESPript/). The secondary structures of CcTet were shown and labeled. The two key loops of CcTet were shaded in cyan, the substrate selection region was shaded in orange, and the long-disordered loop in hTET2 was shaded in grey. The residues that involved in α-KG/Ion/dsDNA binding were labeled with spheres. The finger residue V232 was marked with a red star, and two key residues G331 and D337 involved in substrate selection were marked with blue stars.
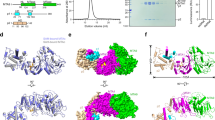
Extended Data Fig. 4 Superposition of CcTet (green) and CcTet-6mA-dsDNA (cyan) complex.
The black arrows indicate the rotation direction of the residue sidechains.
Extended Data Fig. 5 The interactions between DNA bases and CcTet residues.
CcTet and DNA chains were colored in cyan and wheat/palegreen. The two water molecules were shown in spheres and labeled as W1 and W2. The yellow dashes indicate H-bonds between two atoms.
Extended Data Fig. 6 Superposition of CcTet-6mA-dsDNA (cyan/wheat) with FTO-6mA-ssDNA (green/purple, pdb code: 5ZMD) complexes.
(a) Superposition of 6 mA in CcTet active pocket and 6 mA (purple) in FTO (green) active pocket. The wheat region indicates the key FTO residue that was reported to play key roles in m6A catalysis. (b) LC-MS/MS analysis of 6 mA demethylation by CcTet mutations. N = 3 biologically independent experiments. Data represent mean values ± s.d.
Extended Data Fig. 7 Superposition of CcTet-dsDNA with NgTet1-dsDNA or CMD1-dsDNA complex.
The yellow dashes indicate the H-bonds, and the grey dashes indicate the distance between two atoms, and the distances were labeled. (a) Superposition of CcTet-6mA-dsDNA (cyan/wheat) with NgTet1-5mC-dsDNA (yellow/orange, pdb code: 5CG9) complex. (b) Superposition of CcTet-6mA-dsDNA (cyan/wheat) with CMD1-5mC-dsDNA (slate/red, pdb code: 7CY8) complex.
Extended Data Fig. 8
Dot Blot analysis of concentration-dependent 6 mA (a) and 5mC (b) demethylation on Coprinopsis cinerea genomic DNA (mycelium stage) catalyzed by CcTet or D337F mutant. Upper panels show the representative antibody (anti-5mC or 6 mA) dot blot for the purified Coprinopsis cinerea genomic DNA; lower panels show the methylene blue staining to validate the equal loading amount of DNA.
Extended Data Fig. 9
Dot Blot analysis of concentration-dependent 6 mA (a) and 5mC (b) demethylation on green alga (Chlamydomonas reinhardtii) genomic DNA catalyzed by CcTet or D337F mutant. Upper panels show the representative antibody (anti-5mC or 6 mA) dot blot for the purified Chlamydomonas reinhardtii genomic DNA; lower panels show the methylene blue staining to validate the equal loading amount of DNA.
Supplementary information
Supplementary Information
Supplementary Tables 1 and 2 and Figs. 1–17.
Supplementary Data
Source data for the supplementary figures.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 8
Unprocessed western blots and gels.
Source Data Extended Data Fig. 9
Unprocessed western blots and gels.
Rights and permissions
About this article

Cite this article
Mu, Y., Zhang, L., Hu, J. et al. A fungal dioxygenase CcTet serves as a eukaryotic 6mA demethylase on duplex DNA. Nat Chem Biol 18, 733–741 (2022). https://doi.org/10.1038/s41589-022-01041-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41589-022-01041-3
This article is cited by
-
Time series-based hybrid ensemble learning model with multivariate multidimensional feature coding for DNA methylation prediction
BMC Genomics (2023)
-
Making a 6mA demethylase
Nature Chemical Biology (2022)
