
Abstract
Oxidative stress is a defining feature of most cancers, including those that stem from carcinogenic infections. Reactive oxygen species can drive tumor formation, yet the molecular oxidation events that contribute to tumorigenesis are largely unknown. Here we show that inactivation of a single, redox-sensitive cysteine in the host protease legumain, which is oxidized during infection with the gastric cancer-causing bacterium Helicobacter pylori, accelerates tumor growth. By using chemical proteomics to map cysteine reactivity in human gastric cells, we determined that H. pylori infection induces oxidation of legumain at Cys219. Legumain oxidation dysregulates intracellular legumain processing and decreases the activity of the enzyme in H. pylori-infected cells. We further show that the site-specific loss of Cys219 reactivity increases tumor growth and mortality in a xenograft model. Our findings establish a link between an infection-induced oxidation site and tumorigenesis while underscoring the importance of cysteine reactivity in tumor growth.

This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
Supplementary information is available for this paper. Mass-spectrometry proteomics data that support the findings of this study have been deposited in ProteomeXchange via the PRIDE partner repository with the dataset identifier PXD025841. Source data are provided with this paper.
References
Plummer, M. et al. Global burden of cancers attributable to infections in 2012: a synthetic analysis. Lancet Glob. Health 4, e609–e616 (2016).
Robinson, K., Letley, D. P. & Kaneko, K. The human stomach in health and disease: infection strategies by Helicobacter pylori. Curr. Top. Microbiol. Immunol. 400, 1–26 (2017).
Davies, G. R. et al. Helicobacter pylori stimulates antral mucosal reactive oxygen metabolite production in vivo. Gut 35, 179–185 (1994).
Katsurahara, M. et al. Reactive nitrogen species mediate DNA damage in Helicobacter pylori-infected gastric mucosa. Helicobacter 14, 552–558 (2009).
Butcher, L. D., den Hartog, G., Ernst, P. B. & Crowe, S. E. Oxidative stress resulting from Helicobacter pylori infection contributes to gastric carcinogenesis. Cell Mol. Gastroenterol. Hepatol. 3, 316–322 (2017).
Hardbower, D. M., de Sablet, T., Chaturvedi, R. & Wilson, K. T. Chronic inflammation and oxidative stress: the smoking gun for Helicobacter pylori-induced gastric cancer? Gut Microbes 4, 475–481 (2013).
Shirin, H. et al. Helicobacter pylori decreases gastric mucosal glutathione. Cancer Lett. 164, 127–133 (2001).
Liu, Y. et al. Nuclear lactate dehydrogenase A senses ROS to produce alpha-hydroxybutyrate for HPV-induced cervical tumor growth. Nat. Commun. 9, 4429 (2018).
Teoh, M. L., Turner, P. V. & Evans, D. H. Tumorigenic poxviruses up-regulate intracellular superoxide to inhibit apoptosis and promote cell proliferation. J. Virol. 79, 5799–5811 (2005).
Chaturvedi, R. et al. Increased Helicobacter pylori-associated gastric cancer risk in the Andean region of Colombia is mediated by spermine oxidase. Oncogene 34, 3429–3440 (2015).
Hussain, S. P., Hofseth, L. J. & Harris, C. C. Radical causes of cancer. Nat. Rev. Cancer 3, 276–285 (2003).
Anastasiou, D. et al. Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science 334, 1278–1283 (2011).
Bell, E. L. et al. The Qo site of the mitochondrial complex III is required for the transduction of hypoxic signaling via reactive oxygen species production. J. Cell Biol. 177, 1029–1036 (2007).
Horak, P. et al. Negative feedback control of HIF-1 through REDD1-regulated ROS suppresses tumorigenesis. Proc. Natl Acad. Sci. USA 107, 4675–4680 (2010).
Cao, J. et al. Prdx1 inhibits tumorigenesis via regulating PTEN/AKT activity. EMBO J. 28, 1505–1517 (2009).
Irani, K. et al. Mitogenic signaling mediated by oxidants in Ras-transformed fibroblasts. Science 275, 1649–1652 (1997).
Truong, T. H. et al. Molecular basis for redox activation of epidermal growth factor receptor kinase. Cell Chem. Biol. 23, 837–848 (2016).
De Marco, F. et al. Oxidative stress in HPV-driven viral carcinogenesis: redox proteomics analysis of HPV-16 dysplastic and neoplastic tissues. PLoS ONE 7, e34366 (2012).
Li, C. Q., Pignatelli, B. & Ohshima, H. Increased oxidative and nitrative stress in human stomach associated with cagA+ Helicobacter pylori infection and inflammation. Dig. Dis. Sci. 46, 836–844 (2001).
Gordon, E. M. & Hatzios, S. K. Chemical tools for decoding redox signaling at the host−microbe interface. PLoS Pathog. 16, e1009070 (2020).
Shi, Y. & Carroll, K. S. Activity-based sensing for site-specific proteomic analysis of cysteine oxidation. Acc. Chem. Res. 53, 20–31 (2020).
Weerapana, E. et al. Quantitative reactivity profiling predicts functional cysteines in proteomes. Nature 468, 790–795 (2010).
Stairs, D. B. et al. Deletion of p120-catenin results in a tumor microenvironment with inflammation and cancer that establishes it as a tumor suppressor gene. Cancer Cell 19, 470–483 (2011).
Meng, X. et al. RPL23 links oncogenic RAS signaling to p53-mediated tumor suppression. Cancer Res. 76, 5030–5039 (2016).
Zhou, S. et al. Peroxiredoxin 6 homodimerization and heterodimerization with glutathione S-transferase pi are required for its peroxidase but not phospholipase A2 activity. Free Radic. Biol. Med. 94, 145–156 (2016).
D’Costa, Z. C. et al. TBX2 represses CST6 resulting in uncontrolled legumain activity to sustain breast cancer proliferation: a novel cancer-selective target pathway with therapeutic opportunities. Oncotarget 5, 1609–1620 (2014).
Li, N. et al. Effects of legumain as a potential prognostic factor on gastric cancers. Med. Oncol. 30, 621 (2013).
Liu, C., Sun, C., Huang, H., Janda, K. & Edgington, T. Overexpression of legumain in tumors is significant for invasion/metastasis and a candidate enzymatic target for prodrug therapy. Cancer Res. 63, 2957–2964 (2003).
Shen, L. et al. Legumain-deficient macrophages promote senescence of tumor cells by sustaining JAK1/STAT1 activation. Cancer Lett. 472, 40–49 (2020).
Wang, H., Chen, B., Lin, Y., Zhou, Y. & Li, X. Legumain promotes gastric cancer progression through tumor-associated macrophages in vitro and in vivo. Int. J. Biol. Sci. 16, 172–180 (2020).
Liang, D. S. et al. Legumain protease-sheddable PEGylated, tuftsin-modified nanoparticles for selective targeting to tumor-associated macrophages. J. Drug Target 30, 82–93 (2022).
Manoury, B. et al. An asparaginyl endopeptidase processes a microbial antigen for class II MHC presentation. Nature 396, 695–699 (1998).
Lalmanach, G. et al. Regulation of the proteolytic activity of cysteine cathepsins by oxidants. Int. J. Mol. Sci. 21, 1944 (2020).
Zhou, Y. et al. Chemoproteomic strategy to quantitatively monitor transnitrosation uncovers functionally relevant S-nitrosation sites on Cathepsin D and HADH2. Cell Chem. Biol. 23, 727–737 (2016).
Dall, E. & Brandstetter, H. Activation of legumain involves proteolytic and conformational events, resulting in a context- and substrate-dependent activity profile. Acta Crystallogr. F 68, 24–31 (2012).
Xiao, H. et al. A quantitative tissue-specific landscape of protein redox regulation during aging. Cell 180, 968–983 e924 (2020).
Edgington, L. E. et al. Functional imaging of legumain in cancer using a new quenched activity-based probe. J. Am. Chem. Soc. 135, 174–182 (2013).
Dall, E. & Brandstetter, H. Mechanistic and structural studies on legumain explain its zymogenicity, distinct activation pathways, and regulation. Proc. Natl Acad. Sci. USA 110, 10940–10945 (2013).
Capurro, M. I. et al. VacA generates a protective intracellular reservoir for Helicobacter pylori that is eliminated by activation of the lysosomal calcium channel TRPML1. Nat. Microbiol. 4, 1411–1423 (2019).
Dall, E. & Brandstetter, H. Structure and function of legumain in health and disease. Biochimie 122, 126–150 (2016).
Lunde, N. N., Bosnjak, T., Solberg, R. & Johansen, H. T. Mammalian legumain − a lysosomal cysteine protease with extracellular functions? Biochimie 166, 77–83 (2019).
Lin, Y. et al. Functional role of asparaginyl endopeptidase ubiquitination by TRAF6 in tumor invasion and metastasis. J. Natl Cancer Inst. 106, dju012 (2014).
Lamb, A. et al. Helicobacter pylori CagA activates NF-kappaB by targeting TAK1 for TRAF6-mediated Lys 63 ubiquitination. EMBO Rep. 10, 1242–1249 (2009).
Chen, J. M., Fortunato, M. & Barrett, A. J. Activation of human prolegumain by cleavage at a C-terminal asparagine residue. Biochem. J. 352, 327–334 (2000).
Lee, K. E. et al. Helicobacter pylori and interleukin-8 in gastric cancer. World J. Gastroenterol. 19, 8192–8202 (2013).
Noto, J. M., Romero-Gallo, J., Piazuelo, M. B. & Peek, R. M. The Mongolian gerbil: a robust model of Helicobacter pylori-induced gastric inflammation and cancer. Methods Mol. Biol. 1422, 263–280 (2016).
Wu, W. et al. Targeting cell-impermeable prodrug activation to tumor microenvironment eradicates multiple drug-resistant neoplasms. Cancer Res. 66, 970–980 (2006).
Chen, J. M. et al. Cloning, isolation, and characterization of mammalian legumain, an asparaginyl endopeptidase. J. Biol. Chem. 272, 8090–8098 (1997).
Pettersen, E. F. et al. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Ran, F. A. et al. Genome engineering using the CRISPR−Cas9 system. Nat. Protoc. 8, 2281–2308 (2013).
Takacs, C. N. et al. Green fluorescent protein-tagged apolipoprotein E: a useful marker for the study of hepatic lipoprotein egress. Traffic 18, 192–204 (2017).
Lin, A. S. et al. Temporal control of the Helicobacter pylori Cag type IV secretion system in a Mongolian gerbil model of gastric carcinogenesis. mBio 11, e01296-20 (2020).
Hatzios, S. K. et al. Chemoproteomic profiling of host and pathogen enzymes active in cholera. Nat. Chem. Biol. 12, 268–274 (2016).
Eng, J. K., McCormack, A. L. & Yates, J. R. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J. Am. Soc. Mass Spectrom. 5, 976–989 (1994).
Tabb, D. L., McDonald, W. H. & Yates, J. R. III DTASelect and Contrast: tools for assembling and comparing protein identifications from shotgun proteomics. J. Proteome Res 1, 21–26 (2002).
Bak, D. W., Pizzagalli, M. D. & Weerapana, E. Identifying functional cysteine residues in the mitochondria. ACS Chem. Biol. 12, 947–957 (2017).
Perez-Riverol, Y. et al. The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucleic Acids Res. 47, D442–D450 (2019).
Abu-Remaileh, M. et al. Lysosomal metabolomics reveals V-ATPase- and mTOR-dependent regulation of amino acid efflux from lysosomes. Science 358, 807–813 (2017).
Kamitani, T., Kito, K., Nguyen, H. P. & Yeh, E. T. Characterization of NEDD8, a developmentally down-regulated ubiquitin-like protein. J. Biol. Chem. 272, 28557–28562 (1997).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408 (2001).
Acknowledgements
We thank V. Muthusamy (Center for Precision Cancer Modeling, Yale Cancer Center) and J. Nikolaus (West Campus Imaging Core, Yale University) for experimental assistance. We also thank R. Gaudet, J. MacMicking, R. Morris, R. Wasko, C. Takacs, S. Simon and M. Bogyo for reagents and/or technical support. We are grateful to the Hatzios lab, C. Crews and A. Goodman for comments on the manuscript, and to D. Monack and M. Waldor for helpful discussions. This work was supported by National Institutes of Health (NIH) Training Grant T32 GM067543 to Y.K. and E.M.G., and T32 AI007281 to J.H.B.S.; a Gruber Science Fellowship to E.M.G.; NSF Graduate Research Fellowship DGE 1147470 and a Stanford Graduate Fellowship to C.F.; NIH AI118932, CA116087 and Department of Veterans Affairs BX004447 to T.L.C.; Novo Nordisk Foundation Challenge Programme NNF19OC0056411 to M.R.A.; NIH R35GM134964 to E.W.; NIH R35GM137952, American Cancer Society Institutional Research Grant #IRG 17-172-57, a Pilot Grant from the Yale Cancer Center and a Conquer Cancer Now Award from the Concern Foundation to S.K.H.; and NIH Research Grant CA-16359 from the National Cancer Institute.
Author information
Authors and Affiliations
Contributions
Y.K. and S.K.H. developed the concept for the study. Y.K. and D.W.B. carried out formal analysis of the results. Y.K., D.W.B. and E.M.G. undertook the investigation. C.F., J.H.B.S., T.L.C., M.R.A. and E.W. were responsible for the study resources. Y.K. and D.W.B. curated the data. Y.K. and S.K.H. wrote the original draft of the manuscript. All authors reviewed and edited the manuscript. Y.K., D.W.B., E.M.G. and S.K.H. visualized the data. S.K.H. supervised the project. E.W. and S.K.H. undertook administration of the project. T.L.C., M.R.A., E.W. and S.K.H. acquired funding for the work.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Chemical Biology thanks Nina Salama, Jing Yang and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 H. pylori infection increases ROS accumulation, decreases GSH levels, and enhances protein cysteine sulfenylation in AGS cells.
(a) Intracellular ROS in AGS cells incubated with H. pylori G27MA (MOI 50, 18 h), 5 mM hydrogen peroxide for 1 h, or media alone were quantified using the fluorogenic ROS indicator CM-H2DCFDA (left). Fluorescence measurements were normalized by the total number of live cells per condition (right). (b) Intracellular GSH levels in H. pylori-infected (H. pylori G27MA, MOI 50, 18 h) and uninfected AGS cells (left), normalized by the total number of live cells per condition (right). Data represent three independent experiments. Each circle represents an independent experiment. Bars represent means ± SD. *P = 0.03; **P = 0.007; ***P = 0.005; ****P < 0.0001; n.s., not significant. Two-way analysis of variance (ANOVA) with Šídák’s multiple comparisons test was used for (a); two-tailed unpaired t-test was used for (b). (c) Western blot analysis of biotinylated proteins in H. pylori-infected (H. pylori G27MA, MOI 25, 18 h) and uninfected AGS cells. Proteins labeled with the sulfenic acid-specific probe DYn-2 were conjugated with diazo biotin azide and enriched on streptavidin beads prior to Western blot analysis. Western blot analysis was performed twice with consistent results.
Extended Data Fig. 2 Supplementary data for Fig. 1b,c.
(a) L/H ratios of peptides identified in IA-enriched samples from H. pylori-infected (H. pylori G27MA, MOI 50, 18 h) and uninfected AGS cells. (b) L/H ratios of peptides identified in unenriched lysates from H. pylori-infected (H. pylori G27MA, MOI 50, 18 h) and uninfected AGS cells. (c) Representative light (red) and heavy (blue) extracted ion chromatographs (EICs) (top) and isotopic envelopes (bottom) of cysteine-containing peptides from IA-enriched samples with protein abundance-corrected L/H ≤ 0.5. The L/H ratio of each peptide is shown above the corresponding EIC. The average L/H ratios (n = 3) of peptides are shown in (a) and (b). Representative L/H ratios of peptides are shown in (c).
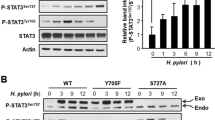
Extended Data Fig. 3 Legumain consistently exhibits reduced cysteine reactivity in H. pylori-infected cells under various infection conditions.
(a) Western blot analysis of legumain in H. pylori-infected (H. pylori G27MA, MOI 50, 18 h) and uninfected KATO III cells before (input) and after IA enrichment. (b) Western blot analysis of legumain in uninfected AGS cells and AGS cells infected with the H. pylori strains G27 or PMSS1 (MOI 50, 18 h) before (input) and after IA enrichment. (c) Western blot analysis of legumain in uninfected AGS cells and AGS cells infected with H. pylori G27MA for 18 h at MOI 25 or 50 before (input) and after IA enrichment. (d) Quantification of AGS cell viability following infection with H. pylori G27MA (MOI 25, 18 h) or incubation with media alone for 18 h. Data represent three independent experiments. Bars represent means ± SD. Each circle represents an independent experiment. n.s., not significant, by two-tailed unpaired t-test. Western blot analyses were performed two (c) or three (a, b) times with consistent results. Band intensities of mature legumain and ACAT1 were normalized by the corresponding band intensities in the uninfected sample.
Extended Data Fig. 4 Biochemical validation of legumain-deficient KO cells generated via CRISPR/Cas9 genome editing.
Western blot analysis of legumain and actin in WT AGS and KO cells. Western blot analysis was performed three times with consistent results.
Extended Data Fig. 5 Supplementary data for Fig. 2b.
Western blot analysis of (a) HA (top), protein markers of various subcellular compartments (S6K1, cytosol; VDAC1, mitochondria; CALR, endoplasmic reticulum), and (b) cathepsins B (left) and D (right) before (input) and after (anti-HA IP) lysosome enrichment of H. pylori-infected (H. pylori G27MA, MOI 25, 18 h) and uninfected AGS cells. Western blot analyses were performed three times with consistent results.
Extended Data Fig. 6 Supplementary data for Fig. 2h.
Western blot analysis of legumain, HA-tagged ubiquitin, and Myc-tagged TRAF6 before (left, input) and after (right, IP) anti-FLAG IP of lysates from H. pylori-infected and uninfected HEK293T cells transiently transfected with FLAG-tagged LGMN, HA-tagged UBB, and/or Myc-tagged TRAF6. Cells were infected with H. pylori G27MA at MOI 25 for 15 h followed by a 3-h incubation in serum-deficient medium. Western blot analyses were performed three times with consistent results.
Extended Data Fig. 7 Intracellular legumainC219A is not processed to the mature enzyme.
Western blot analysis of legumain in KO cells transfected with WT LGMN or LGMNC219A. Western blot analysis was performed three times with consistent results.
Extended Data Fig. 8 KO cells expressing WT legumain or legumainC219S exhibit similar NF-κB activity and IL-8 expression during H. pylori infection.
(a) Luminescence of H. pylori-infected (H. pylori G27MA, MOI 25, 18 h) and uninfected KO cells transduced with WT LGMN or LGMNC219S and transiently transfected with the NF-κB-inducible luciferase reporter pNiFty2-Luc. (b) RT-qPCR analysis of IL-8 expression normalized to GAPDH expression in H. pylori-infected (H. pylori G27MA, MOI 25, 18 h) and uninfected KO cells transduced with WT LGMN or LGMNC219S. Data represent three independent experiments. Each circle represents an independent experiment. Bars represent means ± SD. n.s., not significant. Two-way analysis of variance (ANOVA) with Tukey’s multiple comparisons test was used in (a); two-tailed unpaired t-test was used in (b).
Extended Data Fig. 9 Legumain is expressed in the stomach tissue of H. pylori-infected gerbils.
Representative confocal micrographs with immunofluorescent detection of legumain (LGMN; green) in gastric tissue sections from H. pylori-infected gerbils. Sections with labeled legumain (top) or stained with secondary antibody alone (bottom) are shown. Tissue sections were counterstained with DAPI (blue) and wheat germ agglutinin (WGA; red) to detect nuclei and cellular glycoconjugates, respectively. Sections from six gerbils, using at least two different fields of view per section, were analyzed with consistent results. Scale bar, 40 µm.
Extended Data Fig. 10 Supplementary data for Fig. 4.
(a) Western blot analysis of legumain in two independently derived clonal lines of KO cells transduced with WT LGMN (WT#1 and WT#2) or LGMNC219S (C219S#1 and C219S#2). Western blot analysis was performed three times with consistent results. (b) Cell viability of WT AGS cells and WT#1, WT#2, C219S#1, and C219S#2 cells quantified by alamarBlue fluorescence. Data represent three independent experiments. Error bars represent means ± SD. (c) Average tumor volume of Rag2−/− IL2RG−/− mice subcutaneously implanted with KO cells transduced with WT LGMN or LGMNC219S. These data represent a second independent experiment performed using n = 12 mice and a single clonal cell line per condition (WT#1 and C219S#1). Error bars represent means ± SEM. ****P < 0.0001 by two-tailed unpaired t-test. (d) Survival curve of mice in (c). Tick marks represent censored events. ****P < 0.0001 by Mantel-Cox test.
Supplementary information
Supplementary Information
Supplementary Tables 1−3.
Supplementary Data 1
Cysteine peptides quantified by reactivity profiling and total protein abundance quantified by SILAC mass-spectrometry analysis in H. pylori-infected versus uninfected human gastric cells.
Source data
Source Data Fig. 1
Full-length western blots/gels.
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Full-length western blots/gels.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Full-length western blots/gels.
Source Data Fig. 4
Full-length western blots/gels.
Source Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 1
Full-length western blots/gels.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Full-length western blots/gels.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Full-length western blots/gels.
Source Data Extended Data Fig. 5
Full-length western blots/gels.
Source Data Extended Data Fig. 6
Full-length western blots/gels.
Source Data Extended Data Fig. 7
Full-length western blots/gels.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 10
Full-length western blots/gels.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
About this article

Cite this article
Kovalyova, Y., Bak, D.W., Gordon, E.M. et al. An infection-induced oxidation site regulates legumain processing and tumor growth. Nat Chem Biol 18, 698–705 (2022). https://doi.org/10.1038/s41589-022-00992-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41589-022-00992-x
This article is cited by
-
Integrative analysis of TBI data reveals Lgmn as a key player in immune cell-mediated ferroptosis
BMC Genomics (2023)
-
Oxidize me to the space
Nature Chemical Biology (2022)
