
Abstract
H3K4me1 methyltransferases MLL3 (KMT2C) and MLL4 (KMT2D) are critical for enhancer activation, cell differentiation and development. However, roles of MLL3/4 enzymatic activities and MLL3/4-mediated enhancer H3K4me1 in these processes remain unclear. Here we report that constitutive elimination of both MLL3 and MLL4 enzymatic activities prevents initiation of gastrulation and leads to early embryonic lethality in mice. However, selective elimination of MLL3/4 enzymatic activities in embryonic, but not extraembryonic, lineages leaves gastrulation largely intact. Consistent with this, embryonic stem cells (ESCs) lacking MLL3/4 enzymatic activities can differentiate toward the three embryonic germ layers but show aberrant differentiation to extraembryonic endoderm (ExEn) and trophectoderm. The failure in ExEn differentiation can be attributed to markedly reduced enhancer-binding of the lineage-determining transcription factor GATA6. Furthermore, we show that MLL3/4-catalyzed H3K4me1 is largely dispensable for enhancer activation during ESC differentiation. Together, our findings suggest a lineage-selective, but enhancer activation-independent, role of MLL3/4 methyltransferase activities in early embryonic development and ESC differentiation.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
All datasets described in the paper have been deposited in NCBI Gene Expression Omnibus under accession number GSE154475. The mm9 was used as the mouse genome reference. Source data are provided with this paper.
Code availability
No custom code package or newly developed algorithm was generated in this study.
References
Creyghton, M. P. et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl Acad. Sci. USA 107, 21931–21936 (2010).
Jin, Q. et al. Distinct roles of GCN5/PCAF-mediated H3K9ac and CBP/p300-mediated H3K18/27ac in nuclear receptor transactivation. EMBO J. 30, 249–262 (2011).
Lee, J. E. et al. H3K4 mono- and di-methyltransferase MLL4 is required for enhancer activation during cell differentiation. eLife 2, e01503 (2013).
Wang, C. et al. Enhancer priming by H3K4 methyltransferase MLL4 controls cell fate transition. Proc. Natl Acad. Sci. USA 113, 11871–11876 (2016).
Lai, B. et al. MLL3/MLL4 are required for CBP/p300 binding on enhancers and super-enhancer formation in brown adipogenesis. Nucleic Acids Res. 45, 6388–6403 (2017).
Froimchuk, E., Jang, Y. & Ge, K. Histone H3 lysine 4 methyltransferase KMT2D. Gene 627, 337–342 (2017).
Dorighi, K. M. et al. Mll3 and Mll4 facilitate enhancer RNA synthesis and transcription from promoters independently of H3K4 monomethylation. Mol. Cell 66, 568–576 e4 (2017).
Jang, Y. et al. H3.3K4M destabilizes enhancer H3K4 methyltransferases MLL3/MLL4 and impairs adipose tissue development. Nucleic Acids Res. 47, 607–620 (2019).
Arnold, S. J. & Robertson, E. J. Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nat. Rev. Mol. Cell Biol. 10, 91–103 (2009).
Tam, P. P. & Loebel, D. A. Gene function in mouse embryogenesis: get set for gastrulation. Nat. Rev. Genet. 8, 368–381 (2007).
Stern, C. D. & Downs, K. M. The hypoblast (visceral endoderm): an evo-devo perspective. Development 139, 1059–1069 (2012).
Keller, G. Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev. 19, 1129–1155 (2005).
Leahy, A., Xiong, J. W., Kuhnert, F. & Stuhlmann, H. Use of developmental marker genes to define temporal and spatial patterns of differentiation during embryoid body formation. J. Exp. Zool. 284, 67–81 (1999).
Bibel, M., Richter, J., Lacroix, E. & Barde, Y. A. Generation of a defined and uniform population of CNS progenitors and neurons from mouse embryonic stem cells. Nat. Protoc. 2, 1034–1043 (2007).
Ngondo, R. P., Cohen-Tannoudji, M. & Ciaudo, C. Fast in vitro procedure to identify extraembryonic differentiation defect of mouse embryonic stem cells. STAR Protoc. 1, 100127 (2020).
Ashokkumar, D. et al. MLL4 is required after implantation whereas MLL3 becomes essential during late gestation. Development 147, dev186999 (2020).
Stower, M. J. & Srinivas, S. Heading forwards: anterior visceral endoderm migration in patterning the mouse embryo. Phil. Trans. R. Soc. B 369, 20130546 (2014).
Arnold, S. J., Sugnaseelan, J., Groszer, M., Srinivas, S. & Robertson, E. J. Generation and analysis of a mouse line harboring GFP in the Eomes/Tbr2 locus. Genesis 47, 775–781 (2009).
Yan, J. et al. Histone H3 lysine 4 monomethylation modulates long-range chromatin interactions at enhancers. Cell Res. 28, 387 (2018).
Weinberger, L., Ayyash, M., Novershtern, N. & Hanna, J. H. Dynamic stem cell states: naive to primed pluripotency in rodents and humans. Nat. Rev. Mol. Cell Biol. 17, 155–169 (2016).
Pijuan-Sala, B. et al. A single-cell molecular map of mouse gastrulation and early organogenesis. Nature 566, 490–495 (2019).
Coucouvanis, E. & Martin, G. R. BMP signaling plays a role in visceral endoderm differentiation and cavitation in the early mouse embryo. Development 126, 535–546 (1999).
Fuchs, C. et al. Self-organization phenomena in embryonic stem cell-derived embryoid bodies: axis formation and breaking of symmetry during cardiomyogenesis. Cells Tissues Organs 195, 377–391 (2012).
Brown, K. et al. eXtraembryonic ENdoderm (XEN) stem cells produce factors that activate heart formation. PLoS ONE 5, e13446 (2010).
Chazaud, C., Yamanaka, Y., Pawson, T. & Rossant, J. Early lineage segregation between epiblast and primitive endoderm in mouse blastocysts through the Grb2-MAPK pathway. Dev. Cell 10, 615–624 (2006).
Niwa, H. et al. Interaction between Oct3/4 and Cdx2 determines trophectoderm differentiation. Cell 123, 917–929 (2005).
Pearton, D. J. et al. Elf5 counteracts precocious trophoblast differentiation by maintaining Sox2 and 3 and inhibiting Hand1 expression. Dev. Biol. 392, 344–357 (2014).
Wamaitha, S. E. et al. Gata6 potently initiates reprograming of pluripotent and differentiated cells to extraembryonic endoderm stem cells. Genes Dev. 29, 1239–1255 (2015).
Pataskar, A. et al. NeuroD1 reprograms chromatin and transcription factor landscapes to induce the neuronal program. EMBO J. 35, 24–45 (2016).
Hayashi, S., Lewis, P., Pevny, L. & McMahon, A. P. Efficient gene modulation in mouse epiblast using a Sox2Cre transgenic mouse strain. Mech. Dev. 119, S97–S101 (2002).
Xu, P. F. et al. Construction of a mammalian embryo model from stem cells organized by a morphogen signalling centre. Nat. Commun. 12, 3277 (2021).
Hnisz, D. et al. Super-enhancers in the control of cell identity and disease. Cell 155, 934–947 (2013).
Bergsland, M. et al. Sequentially acting Sox transcription factors in neural lineage development. Genes Dev. 25, 2453–2464 (2011).
Donnison, M. et al. Loss of the extraembryonic ectoderm in Elf5 mutants leads to defects in embryonic patterning. Development 132, 2299–2308 (2005).
Rickels, R. et al. Histone H3K4 monomethylation catalyzed by Trr and mammalian COMPASS-like proteins at enhancers is dispensable for development and viability. Nat. Genet. 49, 1647–1653 (2017).
Yan, J. et al. Transcription factor binding in human cells occurs in dense clusters formed around cohesin anchor sites. Cell 154, 801–813 (2013).
Ooi, S. K. et al. DNMT3L connects unmethylated lysine 4 of histone H3 to de novo methylation of DNA. Nature 448, 714–717 (2007).
Bleckwehl, T. et al. Enhancer-associated H3K4 methylation safeguards in vitro germline competence. Nat. Commun. 12, 5771 (2021).
Wang, H. et al. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 153, 910–918 (2013).
Chakraborty, S. et al. Enhancer-promoter interactions can bypass CTCF-mediated boundaries and contribute to phenotypic robustness. Nat. Genet. 55, 280–290 (2023).
Koehler, K. R. & Hashino, E. 3D mouse embryonic stem cell culture for generating inner ear organoids. Nat. Protoc. 9, 1229–1244 (2014).
Ran, F. A. et al. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 8, 2281–2308 (2013).
Cao, N. et al. In vitro differentiation of rat embryonic stem cells into functional cardiomyocytes. Cell Res. 21, 1316–1331 (2011).
Ferguson, R. & Subramanian, V. Embryoid body arrays: parallel cryosectioning of spheroid/embryoid body samples for medium through-put analysis. Stem Cell Res. 28, 125–130 (2018).
Shechter, D., Dormann, H. L., Allis, C. D. & Hake, S. B. Extraction, purification and analysis of histones. Nat. Protoc. 2, 1445–1457 (2007).
Xie, G. et al. UTX promotes hormonally responsive breast carcinogenesis through feed-forward transcription regulation with estrogen receptor. Oncogene 36, 5497–5511 (2017).
Cho, Y. W. et al. PTIP associates with MLL3- and MLL4-containing histone H3 lysine 4 methyltransferase complex. J. Biol. Chem. 282, 20395–20406 (2007).
Hong, S. et al. Identification of JmjC domain-containing UTX and JMJD3 as histone H3 lysine 27 demethylases. Proc. Natl Acad. Sci. USA 104, 18439–18444 (2007).
Corces, M. R. et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nat. Methods 14, 959–962 (2017).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Zang, C. et al. A clustering approach for identification of enriched domains from histone modification ChIP-seq data. Bioinformatics 25, 1952–1958 (2009).
Zhang, Y. et al. Model-based analysis of ChIP-seq (MACS). Genome Biol. 9, R137 (2008).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, 2016).
Kolde, R. Pheatmap: pretty heatmaps. R J. 1, 726 (2012).
Huang da, W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57 (2009).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Wang, S. et al. Target analysis by integration of transcriptome and ChIP-seq data with BETA. Nat. Protoc. 8, 2502–2515 (2013).
He, H. H. et al. Nucleosome dynamics define transcriptional enhancers. Nat. Genet. 42, 343–347 (2010).
Whyte, W. A. et al. Master transcription factors and mediator establish super-enhancers at key cell identity genes. Cell 153, 307–319 (2013).
Robinson, J. T. et al. Integrative genomics viewer. Nat. Biotechnol. 29, 24–26 (2011).
Acknowledgements
We thank D. Wan and H. Sun for assistance in EB immunofluorescence, R. Kopyto for assistance in genotyping, X. Zhu for assistance in TE differentiation, A.-K. Hadjantonakis and A. Xiao for suggestions on mouse experiments, H. Yan for suggestions on ESC culture and all members of the Ge laboratory for discussions and support. We also thank NHLBI DNA Sequencing and Genomics Core and UCSD IGM Genomics Center for next-generation sequencing, and NIH HPC group for high-performance computing systems. This work was supported by the Intramural Research Program of NIDDK, NIH to K.G (1ZIADK075003-15).
Author information
Authors and Affiliations
Contributions
G.X. and K.G. conceived the project. G.X., J.-E.L., W.P. and C.L. performed the methodology. G.X., A.D.S., Y.-K.P., Y.J., S.C., J.J.T. and K.M. carried out the experiments. J.-E.L., G.X. and W.P. performed bioinformatic analyses. G.X. and K.G. wrote the paper with inputs from all authors. K.G., P.P.R. and T.S.M. provided supervision and acquired funding.
Corresponding author
Ethics declarations
Competing interests
All authors declare no competing interests.
Peer review
Peer review information
Nature Genetics thanks Andrea Kranz and Yi Zhang for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Loss of MLL3/4 enzymatic activities results in embryonic lethality.
a, Alignment of SET domain sequences in different histone lysine methyltransferases in mice. The highly conserved tyrosine (Y) residues, Y4792 in MLL3 and Y5477 in MLL4, are mutated to alanine residues (A) in mice and/or ESCs in this study. b, Schematic of generating enzyme-dead MLL3 and MLL4 knock-in (KI) mice by Cas9 nickase. sgRNA, single guide RNA; nCas9, Cas9 nickase; ssDNA, single-strand DNA. c, Representative images of embryos with indicated genotypes and H&E-stained lung tissues at E18.5. Scale bar, 5 mm (embryo) or 250 μm (histology). d, Genotypes of progeny from Mll3KI/+;Mll4KI/+ intercrosses at E14.5. Numbers of dead pups are indicated in parentheses. e, Representative images of NANOG, GATA6 and CDX2 IF staining of E3.5 blastocysts. Scale bar, 100 μm. f, Schematic of early embryonic development in mice.
Extended Data Fig. 2 Generation and characterization of ESCs lacking MLL3/4 enzymatic activities.
a, Schematic of generating ESCs harboring enzyme-dead mutant (Y5477A) MLL4. Knock-in was done in Mll3−/−;Mll4f/f ESCs. b, Schematic representation of Mll4 flox, knockout (KO) and knock-in (KI) alleles. In flox and KI alleles, two loxP sites were inserted in the intron before exon 16 and the intron after exon 19; in the KI allele, the Y5477A mutation is located in exon 52. Locations of PCR genotyping primers P1, P2, P3, WF, MF and R are indicated by arrows. The MF primer is specific to the KI allele. c, PCR genotyping using primer pairs indicated in b. Sizes of PCR products are indicated on the left. d-h, Chromatograms of genomic PCR sequencing from two independent KI ES cell lines. Trace files display sequences around the Y5477A mutation (d) or sequences around potential exonic off-target regions of Dusp4 (e), Setd1b (f), Ccdc80 (g) and Fignl1 (h). In the reference sequences, sgRNA-defined target regions and PAM sites are highlighted in yellow and cyan, respectively. i, RNA-seq data of genes expressed in f/f or KI ESCs were presented as scatter plots. Pearson correlation coefficient (R) and the corresponding two-sided P value are shown. The number of genes in each group is indicated in parentheses.
Extended Data Fig. 3 ESCs lacking MLL3/4 enzymatic activities can differentiate towards the three germ layers, but show defects in cardiomyogenesis.
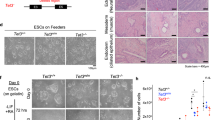
a, b, Immunoblotting in f/f and KI cells at day 0 (D0), D4 and D10 of EB differentiation. Whole cell lysates (a) or histone extracts (b) were analyzed using indicated antibodies. Histone modification levels were quantified and normalized to H3. c, RT-qPCR analysis of primitive streak markers (T, Mixl1) at indicated time points. Data are presented as means ± s.d., n = 3 independent experiments. d,e, RNA-seq data of genes expressed in f/f or KI D4 EBs (d) and f/f or KI D10 EBs (e) were presented as scatter plots. Pearson correlation coefficient (R) and the corresponding two-sided P value are shown. The number of genes in each group is indicated in parentheses. f, Gene set enrichment analysis (GSEA) of 3,845 genes induced from ESC to D10 EB on developmental terms. Nom p-value was determined by a 1000-fold permutation test; FDR q-value was adjusted for multiple hypotheses testing. NES, normalized enrichment score. Statistically significant data are highlighted in bold. g, The percentage of beating EBs at indicated time points. Data are presented as means ± s.d.. The result is from 3 independent experiments with 24 EBs per group. h, Representative microscopic images of attached EBs at D12. Scale bar, 250 μm. i, Beating frequencies of EBs at D12 are presented as dot plots. 24 EBs were measured per group. Horizontal lines represent mean values. Statistical significance was determined by the two-tailed unpaired t-test. j, RT-qPCR analysis of the cardiac progenitor marker (Tbx5) and cardiac myofilament genes (Myh6, Myl2, Ttn) at indicated time points. Data are presented as means ± s.d., n = 3 independent experiments. k, Teratoma assay of f/f and KI ESCs. Representative histological sections of structures belonging to endoderm, mesoderm and ectoderm are shown. Scale bar, 100 μm.
Extended Data Fig. 4 MLL3/4 enzymatic activities are generally dispensable for neural differentiation.
a, Schematic of neural differentiation. PBS, phosphate-buffered saline; PDL, poly-D-lysine; RA, Retinoic acid. b, Representative microscopic images of D8 aggregates and D9 NPCs. Scale bar, 250 μm (upper) or 25 μm (lower). White arrows indicate neural rosettes. c, Expression levels of 4,446 genes induced from ESC to neuron were presented as scatter plots. Pearson correlation coefficient (R) and the corresponding two-sided P value are shown. The number of genes in each group is indicated in parentheses. d, GO analysis of three gene groups in c. Statistical significance was determined by DAVID modified one-sided Fisher’s exact test. e,f, Expression levels of NPC (e) and neuron (f) markers. Data from RNA-seq are presented as dot plots (f/f, n = 2; KI, n = 2). Horizontal lines represent mean values. e, Data of KO ESCs and NPCs from RNA-seq are included.
Extended Data Fig. 5 Loss of MLL3/4 enzymatic activities impairs ExEn gene induction during EB differentiation.
a, GSEA of expression profiles in D4 EBs using cell-type markers defined in E6.5 embryos22. Nom p-value was determined by a 1000-fold permutation test; FDR q-value was adjusted for multiple hypotheses testing. NES, normalized enrichment score. Statistically significant data are highlighted in bold. b, Expression fold changes of markers of visceral endoderm, parietal endoderm and primitive streak, as well as of genes encoding components of BMP, Wnt and Nodal signaling pathways, between KI and f/f D4 EBs. c, Extraembryonic endoderm (ExEn) markers were analyzed by RT-qPCR in ESCs and D4 EBs. Data are presented as means ± s.d., n = 3 independent experiments. d, IF staining of GATA4 in D4 EBs. Scale bar, 100 μm. e, D2 EBs in suspension were plated onto gelatinized surfaces. Representative microscopic images of attached EBs at D4 are shown (left). ExEn-like cells migrating away from the EB periphery are indicated with brackets. Scale bar, 250 μm. Areas of attached EBs at D4 are presented as dot plots (right). Horizontal lines represent mean values. 6 EBs were measured per group. Statistical significance was determined by the two-tailed unpaired t-test. f, During EB differentiation, KI cells were cultivated with ExEn cell-conditioned medium or control medium. The percentage of beating EBs at indicated time points are shown. The result is from 3 independent experiments with 24 EBs per group. Data are presented as means ± s.d.. Statistical significance was determined by the two-tailed unpaired t-test. *, 2.6E-2; **, 3.5E-3; ***,6.7E-4.
Extended Data Fig. 6 ESCs lacking MLL3/4 enzymatic activities show aberrant differentiation to extraembryonic lineages.
a, ExEn markers were analyzed by RT-qPCR before and after 5-day ExEn differentiation. Data are presented as means ± s.d., n = 3 independent experiments. b, ExEn differentiation was extended to 9 days. ExEn markers were analyzed by time-course RT-qPCR at indicated time points. Data are presented as means ± s.d., n = 3 independent experiments. c, ExEn differentiation was extended to 8 days. ExEn markers were analyzed by time-course immunoblotting at indicated time points. Whole cell lysates were analyzed using indicated antibodies. BRG1 is shown as loading control. d, Motif analysis and Venn diagram of GATA6+ AEs in f/f and KI cells after ExEn differentiation. Statistical significance was determined using the SeqPos motif tool with default parameters. e, Whole cell lysates prepared from cells after ExEn differentiation were immunoprecipitated with MLL4 or UTX antibodies. Immunoprecipitated complexes were analyzed by immunoblotting using antibodies against MLL4, UTX and GATA6. f, GO analysis of three gene groups in Fig. 4g. Statistical significance was determined by DAVID modified one-sided Fisher’s exact test. g, ChIP-seq profiles of GATA6, MLL4, H3K4me1 and H3K27ac as well as RNA-seq profiles in f/f and KI cells are displayed on Gata4 and Sox17 loci. GATA6+ MLL4+ de novo AEs are highlighted in shades. h, Immunoblotting of T7-tagged CDX2 and EOMES. f/f and KI ESCs were infected with Doxycycline (Dox)-inducible lentiviral vector expressing T7-tagged CDX2 or EOMES and treated with 1 μg/ml Dox for 1 day. Whole cell lysates were analyzed. BRG1 is shown as loading control. i, After CDX2-stimulated TE differentiation, ratios of giant cell area to total area are presented as dot plots. Horizontal lines represent mean values. At least 6 different fields of view were observed per group for measurement. Statistical significance was determined by the two-tailed unpaired t-test. Data from 2 independent experiments (Exp #1 and Exp #2) are shown.
Extended Data Fig. 7 Lineage-selective roles of MLL3/4 enzymatic activities in early embryonic development.
a, Schematic representation of Mll3 flox and deletion, as well as Mll4SET flox, deletion and KI alleles10. Locations of PCR genotyping primers are indicated by arrows. The MF primer is specific to the KI allele. b, PCR genotyping of E9.5 yolk sac-removed embryos using primers showed in Extended Data Fig. 7a as well as primers detecting Sox2-Cre allele. Sizes of PCR products are indicated on the left. c, Whole cell lysates prepared from E9.5 yolk sac-removed embryos were analyzed with immunoblotting using indicated antibodies. d, Sox2-Ctr and Sox2-dKI embryos were collected at E9.5 for RNA-seq. Levels of expressed genes were presented as scatter plots. Pearson correlation coefficient (R) and the corresponding two-sided P value are shown. The number of genes in each group is indicated in parentheses. e, GSEA of 11,467 expressed genes in E9.5 embryos on developmental terms. Nom p-value was determined by a 1000-fold permutation test; FDR q-value was adjusted for multiple hypotheses testing. NES, normalized enrichment score. Statistically significant data are highlighted in bold. f, Heat maps of gene expression level in E9.5 embryos for markers of germ layer derivatives. Data from 3 embryos were presented for each genotype.
Extended Data Fig. 8 Loss of MLL3/4 enzymatic activities results in redistribution of MLL4 genomic binding.
a, D4 EBs were collected for ChIP-seq of MLL4. 34,772 MLL4+ regions were identified in f/f or KI ESCs. MLL4+ distal regions or promoters were split based on the MLL4 binding intensities in KI EBs compared to f/f EBs. Average profiles (left) and heat maps (right) of 34,772 MLL4+ regions in f/f and KI EBs are shown. b, MLL4+ AEs in ESCs and D4 EBs are depicted by the Venn diagram. 10,767 de novo MLL4+ AEs were divided into three groups based on the changes of MLL4 binding intensities from f/f to KI EBs: increased (Group I), unchanged (Group II) and decreased (Group III). c, Motif analysis of the three groups of de novo MLL4+ AEs. GATA family transcription factors were highlighted in bold. Statistical significance was determined using the SeqPos motif tool with default parameters. d, Heat maps of GATA6 and MLL4 genomic bindings as well as H3K4me1 and H3K27ac enrichments on de novo GATA6+ MLL4+ AEs in f/f and KI D4 EBs. e, Expression fold changes (log2) of genes associated predominantly with each group of de novo MLL4+ AEs. Sample size and RNA-seq data are presented in box plots. Center lines represent median values; the bottom and top of the boxes represent lower and upper quartiles; whiskers were calculated using the Tukey method. Statistical significance was determined by the two-sided Wilcoxon signed-rank test.
Extended Data Fig. 9 MLL3/4 proteins, but not MLL3/4-catalyzed H3K4me1, are required for activation of Group II AEs during early EB differentiation.
a, b, Heat maps (a) and average profiles (b) of MLL4 genomic bindings, H3K4me1 and H3K27ac enrichments in f/f and KO cells on Group II de novo MLL4+ AEs identified in Extended Data Fig. 8b. ChIP-seq data in KO cells were obtained from GSE505344 and reanalyzed. c, ChIP-seq profiles of the same datasets as in Fig. 7c are displayed on Gsc, Mixl1 and Evx1 loci. Group II de novo MLL4+ AEs are highlighted in shades.
Extended Data Fig. 10 MLL3/4-catalyzed H3K4me1 is dispensable for activation of super-enhancers during neural differentiation.
a, GO analysis of genes associated NPC SEs. Statistical significance was determined by DAVID modified one-sided Fisher’s exact test. b, ChIP-seq profiles of MLL4, H3K4me1 and H3K27ac, and RNA-seq profiles in f/f and KI cells are displayed on Nes, Ptn and Pax7 loci. NPC SEs are indicated by purple bars and de novo MLL4+ enhancers in SEs are highlighted in shades.
Supplementary information
Supplementary Information
Supplementary Discussion.
Supplementary Tables 1–7
Table 1: Detailed information of mating experiments. Table 2: Differentially expressed genes in ESCs. Table 3: Differentially expressed genes during EB differentiation. Table 4: Differentially expressed genes during neural differentiation. Table 5: Differentially expressed genes in embryos. Table 6: Information of antibodies. Table 7: Sequences of oligonucleotides.
Source data
Source Data Fig. 2
Unprocessed immunoblots.
Source Data Fig. 4
Unprocessed immunoblots.
Source Data Fig. 5
Unprocessed immunoblots.
Source Data Fig. 6
Unprocessed immunoblots.
Source Data Extended Data Fig. 2
Unprocessed PCR genotyping images.
Source Data Extended Data Fig. 3
Unprocessed immunoblots.
Source Data Extended Data Fig. 6
Unprocessed immunoblots.
Source Data Extended Data Fig. 7
Unprocessed immunoblots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Xie, G., Lee, JE., Senft, A.D. et al. MLL3/MLL4 methyltransferase activities control early embryonic development and embryonic stem cell differentiation in a lineage-selective manner. Nat Genet 55, 693–705 (2023). https://doi.org/10.1038/s41588-023-01356-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-023-01356-4
