
Abstract
Rapeseed (Brassica napus L.) is an important oil-producing crop for the world. Its adaptation, yield and quality have been considerably improved in recent decades, but the genomic basis underlying successful breeding selection remains unclear. Hence, we conducted a comprehensive genomic assessment of rapeseed in the breeding process based on the whole-genome resequencing of 418 diverse rapeseed accessions. We unraveled the genomic basis for the selection of adaptation and agronomic traits. Genome-wide association studies identified 628 associated loci-related causative candidate genes for 56 agronomically important traits, including plant architecture and yield traits. Furthermore, we uncovered nonsynonymous mutations in plausible candidate genes for agronomic traits with significant differences in allele frequency distributions across the improvement process, including the ribosome recycling factor (BnRRF) gene for seed weight. This study provides insights into the genomic basis for improving rapeseed varieties and a valuable genomic resource for genome-assisted rapeseed breeding.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
The raw resequencing data of 418 rapeseed accessions are available in the NCBI Sequence Read Archive under accessions PRJNA416679 and Genome Sequence Archive (GSA) database in BIG Data Center (http://bigd.big.ac.cn/gsa) under accession number CRA005979. The high-coverage resequencing data of 135 rapeseed accessions and Hi-C data were deposited in GSA under accession number CRA005980. The public RNA-seq data used was downloaded from NCBI and the accession numbers are summarized in Supplementary Table 22. The relevant data of genotype and phenotype are available at the website http://brassicanapusdata.cn/DOWNLOAD. All the data and research materials in this study are available upon reasonable request.
Code availability
All the analysis tools used in this study have been published before as described in the Methods and Reporting Summary.
References
Chalhoub, B. et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 345, 950–953 (2014).
Prakash, S., Wu, X.-M. & Bhat, S. R. in Plant Breeding Reviews Vol. 35 (ed. Janick, J.) Ch. 2 (John Wiley & Sons, 2011).
Lu, K. et al. Whole-genome resequencing reveals Brassica napus origin and genetic loci involved in its improvement. Nat. Commun. 10, 1154 (2019).
Bus, A., Korber, N., Snowdon, R. J. & Stich, B. Patterns of molecular variation in a species-wide germplasm set of Brassica napus. Theor. Appl. Genet. 123, 1413–1423 (2011).
Hu, D. D. et al. Exploring the gene pool of Brassica napus by genomic-based approaches. Plant Biotechnol. J. 19, 1693–1712 (2021).
Wei, D. Y. et al. A genome-wide survey with different rapeseed ecotypes uncovers footprints of domestication and breeding. J. Exp. Bot. 68, 4791–4801 (2017).
Wu, D. Z. et al. Whole-genome resequencing of a worldwide collection of rapeseed accessions reveals the genetic basis of ecotype divergence. Mol. Plant 12, 30–43 (2019).
Song, J. M. et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants 6, 34–35 (2020).
An, H. et al. Transcriptome and organellar sequencing highlights the complex origin and diversification of allotetraploid Brassica napus. Nat. Commun. 10, 2878 (2019).
Cheng, F. et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 48, 1218–1224 (2016).
Clark, R. M. et al. Common sequence polymorphisms shaping genetic diversity in Arabidopsis thaliana. Science 317, 338–342 (2007).
Zhou, Z. K. et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 33, 408–414 (2015).
Yano, K. et al. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 48, 927–934 (2016).
Yang, J. H. et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 48, 1225–1232 (2016).
Qian, L. W. et al. Sub-genomic selection patterns as a signature of breeding in the allopolyploid Brassica napus. BMC Genomics 15, 1170 (2014).
Qian, L. W. et al. Deletion of a stay-green gene associates with adaptive selection in Brassica napus. Mol. Plant 9, 1559–1569 (2016).
Shi, J. Q. et al. Linkage and regional association analysis reveal two new tightly-linked major-QTLs for pod number and seed number per pod in rapeseed (Brassica napus L.). Sci. Rep. 5, 14481 (2015).
Zhao, W. et al. Genome-wide identification of QTL for seed yield and yield-related traits and construction of a high-density consensus map for QTL comparison in Brassica napus. Front. Plant Sci. 7, 17 (2016).
Lu, K. et al. Genome-wide association and transcriptome analyses reveal candidate genes underlying yield-determining traits in Brassica napus. Front. Plant Sci. 8, 206 (2017).
Nour-Eldin, H. H. et al. Reduction of antinutritional glucosinolates in Brassica oilseeds by mutation of genes encoding transporters. Nat. Biotechnol. 35, 377–382 (2017).
Wang, B. et al. Dissection of the genetic architecture of three seed-quality traits and consequences for breeding in Brassica napus. Plant Biotechnol. J. 16, 1336–1348 (2018).
Tan, Z. D. et al. Genome- and transcriptome-wide association studies reveal the genetic basis and the breeding history of seed glucosinolate content in Brassica napus. Plant Biotechnol. J. 20, 211–225 (2022).
Wang, Y. H. & Li, J. Y. Molecular basis of plant architecture. Annu. Rev. Plant Biol. 59, 253–279 (2008).
Hu, J. H. et al. Auxin-related genes associated with leaf petiole angle at the seedling stage are involved in adaptation to low temperature in Brassica napus. Environ. Exp. Bot. 182, 104308 (2021).
Chen, Y. N. et al. Over-expression of OsPIN2 leads to increased tiller numbers, angle and shorter plant height through suppression of OsLAZY1. Plant Biotechnol. J. 10, 139–149 (2012).
Liu, J. et al. Natural variation in ARF18 gene simultaneously affects seed weight and silique length in polyploid rapeseed. Proc. Natl Acad. Sci. USA 112, E5123–E5132 (2015).
Shi, J. Q. et al. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus. Genetics 182, 851–861 (2009).
Fang, L. et al. Genomic analyses in cotton identify signatures of selection and loci associated with fiber quality and yield traits. Nat. Genet. 49, 1089–1098 (2017).
Luo, Z. et al. Incorporating pleiotropic quantitative trait loci in dissection of complex traits: seed yield in rapeseed as an example. Theor. Appl. Genet. 130, 1569–1586 (2017).
Radoev, M. et al. Genetic analysis of heterosis for yield and yield components in rapeseed (Brassica napus L.) by quantitative trait locus mapping. Genetics 179, 1547–1548 (2008).
Wang, L. et al. The Arabidopsis chloroplast ribosome recycling factor is essential for embryogenesis and chloroplast biogenesis. Plant Mol. Biol. 74, 47–59 (2010).
King, S. P. et al. Carbohydrate content and enzyme metabolism in developing canola siliques. Plant Physiol. 114, 153–160 (1997).
Bennett, E. J. et al. The role of the pod in seed development: strategies for manipulating yield. New Phytol. 190, 838–853 (2011).
Kondra, Z. & Stefansson, B. Inheritance of the major glucosinolates of rapeseed (Brassica napus) meal. Can. J. Plant. Sci. 50, 643–647 (1970).
Wu, G. et al. Zero erucic acid trait of rapeseed (Brassica napus L.) results from a deletion of four base pairs in the fatty acid elongase 1 gene. Theor. Appl. Genet. 116, 491–499 (2008).
Wang, N. et al. A functional genomics resource for Brassica napus: development of an EMS mutagenized population and discovery of FAE1 point mutations by TILLING. New Phytol. 180, 751–765 (2008).
Jiang, C. C. et al. Quantitative trait loci that control the oil content variation of rapeseed (Brassica napus L.). Theor. Appl. Genet. 127, 957–968 (2014).
Tang, S. et al. Genome- and transcriptome-wide association studies provide insights into the genetic basis of natural variation of seed oil content in Brassica napus. Mol. Plant 14, 470–487 (2021).
Fu, W. F. et al. Acyl-CoA N-acyltransferase influences fertility by regulating lipid metabolism and jasmonic acid biogenesis in cotton. Sci. Rep. 5, 11790 (2015).
Li, L. X., Long, Y., Li, H. & Wu, X. M. Comparative transcriptome analysis reveals key pathways and hub genes in rapeseed during the early stage of Plasmodiophora brassicae infection. Front. Genet. 10, 1275 (2020).
Hurgobin, B. et al. Homoeologous exchange is a major cause of gene presence/absence variation in the amphidiploid Brassica napus. Plant Biotechnol. J. 16, 1265–1274 (2018).
Jin, J. et al. Genetic control of rice plant architecture under domestication. Nat. Genet. 40, 1365–1369 (2008).
Li, F. et al. A genome-wide association study of plant height and primary branch number in rapeseed (Brassica napus). Plant Sci. 242, 169–177 (2016).
Chen, L. et al. A 2.833-kb insertion in BnFLC.A2 and its homeologous exchange with BnFLC.C2 during breeding selection generated early-flowering rapeseed. Mol. Plant 11, 222–225 (2018).
Zheng, M. et al. Knockout of two BnaMAX1 homologs by CRISPR/Cas9- targeted mutagenesis improves plant architecture and increase yield in rapeseed (Brassica napus L.). Plant Biotechnol. J. 18, 644–654 (2020).
Chen, B. Y. et al. Evaluation yield agronomic traits and their genetic variation in 488 global collections of Brassica napus L. Genet. Resour. Crop Evol. 61, 979–999 (2014).
Murray, M. & Thomspin, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 8, 4321–4326 (1980).
Li, L. X. et al. A genome-wide association study reveals new loci for resistance to clubroot disease in Brassica napus. Front. Plant Sci. 7, 1483 (2016).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Li, H. et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Mckenna, A. et al. The genome analysis toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303 (2010).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 38, e164 (2010).
Vilella, A. J. et al. EnsemblCompara genetrees: complete, duplication-aware phylogenetic trees in vertebrates. Genome Res. 19, 327–335 (2009).
Barrett, J. C., Fry, B., Maller, J. & Daly, M. J. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21, 263–265 (2005).
Pickrell, J. K. & Pitchard, J. K. Inference of population splits and mixtures from genone-wide allele frequency data. PLoS Genet. 8, e1002967 (2012).
Patterson, N. et al. Ancient admixture in human history. Genetics 192, 1065–1093 (2012).
Chen, H., Patterson, N. & Reich, D. Population differentiation as a test for selective sweeps. Genome Res. 20, 393–402 (2010).
Alachiotis, N., Stamatakis, A. & Pavlidis, P. OmegaPlus: a scalable tool for rapid detection of selective sweeps in whole-genome datasets. Bioinformatics 28, 2274–2275 (2012).
Zhou, X. & Matthew, S. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 44, 821–824 (2012).
Thompson, J. D., Higgins, D. G. & Gibson, T. J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680 (1994).
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729 (2013).
Fang, C. et al. Genome-wide association studies dissect the genetic networks underlying agronomical traits in soybean. Genome Biol. 18, 161 (2017).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575 (2007).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Xie, T. et al. Biased gene retention during diploidization in Brassica linked to 3D genome organization. Nat. Plants 5, 822–832 (2019).
Servant, N. et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biol. 16, 259 (2015).
Chu, P. et al. An RNA-seq transcriptome analysis of floral buds of an interspecific Brassica hybrid between B. carinata and B. napus. Plant Reprod. 27, 225–237 (2014).
Yong, H. Y. et al. Comparative transcriptome analysis of leaves and roots in response to sudden increase inn salinity in Brassica napus by RNA-seq. BioMed. Res. Int. 27, 225–237 (2014).
Miao, L. Y. et al. Transcriptome analysis of stem and globally comparison with other tissues in Brassica napus. Front. Plant Sci. 7, 1403 (2016).
Wan, H. et al. Time-series analyses of transcriptomes and proteomes reveal molecular networks underlying oil accumulation in canola. Front. Plant Sci. 7, 2007 (2017).
Sun, F. M. et al. The high-quality genome of Brassica napus cultivar ‘ZS11’ reveals the introgression history in semi-winter morphotype. Plant J. 92, 452–468 (2017).
Ye, J. et al. An integrated analysis of QTL mapping and RNA sequencing provides further insights and promising candidates for pod number variation in rapeseed (Brassica napus L.). BMC Plant Biol. 18, 71 (2017).
Trapnell, C. et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 7, 562–578 (2012).
Trapnell, C. et al. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28, 511–515 (2010).
Dai, C. et al. An efficient Agrobacterium-mediated transformation method using hypocotyl as explants for Brassica napus. Mol. Breed. 40, 96 (2020).
Acknowledgements
We thank G. Cai (Oil Crops Research Institute, CAAS) for help with technical analysis. This work was supported by the National Key Research and Development Program of China (2016YFD0100202), Germplasm Resources Protection Project in China (2016NWB040) and Agricultural Science and Technology Innovation Project of Chinese Academy of Agricultural Sciences (CAAS-ASTIP- 2015-OCRI) to X.W. and National Natural Science Foundation of China to J.H. (31901426) and to G.Y. (32072106) and the Young Elite Scientists Sponsorship Program by China Association for Science and Technology (2019QNRC001) to T.X.
Author information
Authors and Affiliations
Contributions
X.W., S.T. and J.H. conceived and designed the research. X.W., B.C., K.X. and H.L. participated in germplasm collection. B.C., J.H., H.L., K.X., F.Z., G.G., L.L., H.G.L., Q.H., J.W., W.S. and Y.L. performed field experiments and phenotyping. J.H., J.Z., X.W., S.T., T.X. and G.J. performed sequencing, genomic variant and analyzed the data. T.X. and F.Z. performed Hi-C experiments and data analysis. G.Y., M.Z., F.Z. and J.H. performed plasmid construction and genetic transformation. F.Z., J.H. and X.Z. conducted gene expression analysis. J.H., J.Z. and T.X. wrote the manuscript. X.W., J.C.P, H.A., S.T. and J.B. revised the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Genetics thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Changes in agronomic traits during rapeseed breeding in Europe and China.
PH, plant height; BH, branch height; BA, branch angle; BN, primary branch number; DM, diameter of main stem; YP, yield per plant; SW, 1,000-seed weight; SP, seeds per pod; SN, silique number per plant; HI, harvest index; FT, flowering time; SOC, seed oil content; GSL, glucosinolate. Different letters above the boxes indicate significant differences (P < 0.05, two-tailed t-test). The center lines indicate the median, box limits represent the upper and lower quartiles, whiskers extend to 1.5× the interquartile range, and dots represent outliers. EU, Europe; CN, China. EU1950&70 s (n = 93); EU1980&90 s (n = 35); CN1950&70 s (n = 73); CN1980&90 s (n = 67); CN00, CN2000&10 s (n = 35) (See Supplementary Fig. 6).
Extended Data Fig. 2 GWAS identification of the candidate genes for flowering time.
a, Manhattan plots of SNP-GWAS for flowering time in Wuhan 2015. The red arrowheads indicate the FT genes (BnaA02g12130D and BnaC06g27090D). b,c, Boxplots for flowering time based on the haplotypes (Hap) of BnaA02g12130D (b) and BnaC06g27090D (c). Center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. Significant differences between the haplotypes were evaluated by two-tailed t-test and shown by P value or different letters (P < 0.01). d, Haplotype frequency changes of BnaA02g12130D and BnaC06g27090D in different ecotypes and breeding eras. e, Local Manhattan plot (top) and LD heatmap (bottom) surrounding the peak for flowering time on chromosome C06 (BnaC06g27090D). The red arrowheads indicate the SNP in candidate gene (BnaC06g27090D). The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). f, XP-CLR plot of BnaC06g27090D. The red horizontal dashed line represents the genome-wide cutoff with the highest being 1%. g, The spectrum of allele frequencies at the causal polymorphisms of BnaC06g27090D. EU, Europe; CN, China. EU50, EU1950&70 s; EU80, EU1980&90 s; CN50, CN1950&70 s; CN80, CN1980&90 s; CN00, CN2000&10 s.
Extended Data Fig. 3 GWAS identification of candidate genes for plant height and branch height.
a, b, Manhattan plots of SNP-GWAS and InDel-GWAS for plant height (a) and branch height (b) in Yangluo 2013. The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). c, Local Manhattan plot (top) and LD heatmap (bottom) surrounding the peak for plant height and branch height on chromosome C07 using SNPs (blue dots) and InDels (red triangles). The candidate region lies between red dashed lines. The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). d, XP-CLR plot of BnaC07g34270D. The red horizontal dashed line represents the genome-wide cutoff with the highest being 1%. e, Boxplot of plant height and branch height for the haplotypes (Hap) of BnaC07g34270D using both SNPs and InDels. Center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. Different letters above the boxes indicate significant differences (P < 0.01, two-tailed t-test) in a pairwise comparison. f, Haplotype frequency changes of BnaC07g34270D in different ecotypes and breeding eras.
Extended Data Fig. 4 GWAS for seed weight and identification of the candidate gene on chromosome A09 and C08.
a, Manhattan plots of SNP-GWAS (blue dots) and InDel-GWAS (red dots) for 1000-seed weight on chromosome A09_random. The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). b, Boxplots for silique length and 1000-seed weight based on the haplotypes of BnaA09g55580D (ARF18). Center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. The P values based on two-tailed t-test are shown. c, Phenotypes of silique length (left) and seed size (right) between 2AF456 (small seed) and 2AF410 (large seed). Scale bar, 1 cm and 0.3 cm. d, Boxplots for 1000-seed weight based on the haplotypes of BnaC08g31420D and frequency changes of BnaC08g31420D in different ecotypes and breeding eras. Center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. Different letters above the boxes indicate significant differences (P < 0.01, two-tailed t-test) in a pairwise comparison. e, XP-CLR plot of BnaC08g32730D (BnRRF). The red horizontal dashed line represents the genome-wide cutoff with the highest being 1%. f, Phenotypes of seeds in WT (Col -0) and Arabidopsis overexpression (OE) lines. Scale bar, 0.5 mm. g, Overexpression (OE) of BnRRF in Arabidopsis increases the seed weight with the expression levels. Error bars, data were presented as the mean ± SD, n = 6 and 3, respectively. Significant differences were evaluated by two-tailed Student’s t-test (** P < 0.01).
Extended Data Fig. 5 GWAS for yield and identification of the candidate genes associated with 1,000-seed weight (SW) and silique length (SL).
a, b, Manhattan plots of SNP-GWAS and InDel-GWAS for SW (a) and SL(b) in Wuhan 2016. The red arrowheads indicate the previous reported genes (ARF18 and CYP78A9). The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). c,d, Boxplot for SW and SL based on the haplotypes of BnaA09g40330D (c) and BnaC08g32200D (d) using SNPs. e, qRT–PCR analysis of the expression of BnaA09g40330D in large seed and small seed accessions. Error bars, data were presented as the mean ± SD, n = 3 biological replicates. f, Gene structure and DNA polymorphism of BnaA09g40330D and BnaC08g32200D. Two nonsynonymous SNPs were found to be associated with the GWAS signals for SW and SL on chromosomes A09 (A09DFG and A09DCG) and C08 (C08LAL and C08LVL), respectively. g, Boxplot for SW and SL based on the haplotypes of the recombination of two alleles on A09 and C08. In c, d and g, center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. The P values based on two-tailed t-test are shown. h, qRT–PCR analysis of the expression of BnaC08g32200D in large seed and small seed accessions. Error bars, data were presented as the mean ± SD, n = 3 biological replicates.
Extended Data Fig. 6 GWAS identification of the candidate genes for glucosinolate (GSL) content.
a, Manhattan plots of InDel-GWAS for GSL content. The red arrowheads indicate the previous reported genes (GTR2 and HAG1). The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). b, Selective sweeps of BnaA02g33270D based on π-ratio and FST between oil- and vegetable-use rapeseeds. c, Local Manhattan plot (top) and LD heatmap (bottom) surrounding the peak for GSLs on chromosome C09 based on SNPs (blue dots) and InDels (red triangles). The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). d, Boxplots for GSLs based on the haplotypes (Hap) of BnaC09g05240D using both SNPs and InDels. e, Haplotype frequency changes of BnaC09g05240D in different ecotypes and breeding eras. f, XP-CLR plot of BnaC09g04850D. The red horizontal dashed line represents the genome-wide cutoff with the highest being 1%. g, Boxplots for GSL based on the haplotypes (Hap) of BnaC09g04850D using both SNPs and InDels. In d and g, center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. Different letters above the boxes indicate significant differences (P < 0.01, two-tailed t-test) in a pairwise comparison. h, Haplotype frequency changes of BnaC09g04850D in different ecotypes and breeding eras.
Extended Data Fig. 7 GWAS identification of candidate genes for erucic acid (C22:1) content.
a, Local Manhattan plot (top) and LD heatmap (bottom) surrounding the peak for C22:1 on chromosome A08 using SNPs. The red dashed line indicates the previous reported genes (FAE1). The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction). b, XP-CLR plot of BnaA08g11130D (FAE1) and BnaA08g11140D (KCS17). The red horizontal dashed line represents the genome-wide cutoff with the highest being 1%. c, Selective sweeps of BnaA08g11130D and BnaA08g11140D based on π-ratio and FST betweenoil- and vegetable-use rapeseeds. d, Exon–intron structure and DNA polymorphism of BnaA08g11140D. e, Boxplots for C22:1 based on the haplotypes (Hap) of BnaA08g11020D and BnaA08g11140D. f, qRT–PCR analysis of the expression of BnaA08g11140D in large seed and small seed accessions. In e and f, center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. Significant differences between the haplotypes or pairwise comparison were evaluated by two-tailed t-test and shown by P values or different letters (P < 0.01). g, Haplotype frequency changes of BnaA08g11140D in different ecotypes and breeding eras.
Extended Data Fig. 8 GWAS analysis and identification of the candidate genes for clubroot resistance.
a, Manhattan plots for disease index of clubroot on chromosome C04 using SNPs. Arrowhead indicates the position of strong associated peak with the SNP (C04:32815573). The horizontal dashed line represents the significance threshold (P < 1 × 10−6, Bonferroni correction).b, Heatmap showing the differentially expressed genes during four stages after infecting clubroot in resistant and sensitive rapeseeds (Li et al., 2020). c, Phenotypic features clubroot tolerant (2AF430 and 2AF451) and sensitive (2AF459 and 2AF478) accessions. Scale bar, 5 cm. d, Exon–intron structure and DNA polymorphism of BnaC04g30930D. e, Boxplots for disease index of clubroot based on the haplotypes (Hap) of BnaC04g30930D. Center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range, and dots represent outliers. Different letters above the boxes indicate significant differences (P < 0.01, two-tailed t-test) in a pairwise comparison. f, Local Manhattan plot (top) and LD heatmap (bottom) surrounding the peak for disease index of clubroot on chromosome C04 (BnaC04g30930D). g, Expression levels of the clubroot resistance candidate gene BnaC04g30930D in different rapeseed accessions by qRT–PCR. Error bars, data were presented as the mean ± SD, n = 3 biological replicates. Different letters above the boxes indicate significant differences (P < 0.05, two-tailed t-test).
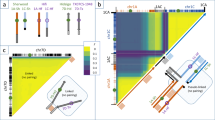
Extended Data Fig. 9 Association networks across different traits in rapeseed and the chromatin interactions (Hi-C) for plant architecture (PA) and yield (PY) traits.
a, Network for 17 traits based on the link powers between loci. The nodes represent traits and their corresponding loci (see Supplementary Table 19). The edges between the loci from different traits are linked by LD. Only the edges with an average LD ≥ 0.4 are shown. BH, branch height; BA, branch angle; BN, primary branch number; DM, diameter of main stem; FT, flowering time; PH, plant height; SI, Silique number of main inflorenscence; SL, silique length; SN, silique number per plant; SP, seeds per pod; SS, Stem strength; SW, 1,000-seed weight; YP, yield per plant. The octagon represents traits and circle indicates different loci. Blue, flowering time; skyblue, PA; purple, PY; yellow, fatty acid components. b, High chromatin interactions (top 5%) between the loci of PA and PY. c, Collinearity analysis of the causal genes for PA and PY with the high chromatin interactions.
Extended Data Fig. 10 Stepwise selection for agronomic traits during rapeseed breeding.
a, Allelic distributions of genes for plant growth and circadian rhythm. b, Allelic distributions of genes involved in plant architecture and yield. c, The spectrum of allele frequencies at the causal polymorphisms in genes involved in seed quality. EU50, EU1950&70 s; EU80, EU1980&90 s; CN50, CN1950&70 s; CN80, CN1980&90 s; CN00, CN2000&10 s. PH, plant height; BH, branch height; YP, yield per plant; SP, seeds per pod. GSL, glucosinolate; SOC, seed oil content.
Supplementary information
Supplementary Information
Supplementary Figs. 1–44, Tables 2–4, 8, 13–15, 18, 22 and 24 and captions for Tables 1, 5–7, 9–12, 16, 17, 19–21 and 23.
Supplementary Tables
Supplementary Tables: 1, Summary of 418 accessions for genome resequencing in this study; 5, Selective sweeps among the three ecotypes of semiwinter versus winter and spring versus winter; 6, Selective sweeps among different breeding eras in Europe and China; 7, Summary of QTL and genes for PA, FT and PY that overlap with selective sweeps; 9, Different environments for the 56 traits of rapeseed phenotyping in this study; 10, Summary of quantitative traits phenotyped in six environments; 11, Genome-wide association signals of agronomics traits by SNP-GWAS; 12, Genome-wide association signals of agronomics traits by InDel-GWAS; 16, Summary of the InDels from exons overlapped with GWAS loci identified in this study; 17, Summary of the haplotypes of candidate genes identified by SNP-GWAS for different agronomic traits; 19, Link loci across 17 traits based on the pairwise LD value in rapeseed; 20, Characterization of large interchromosomal translocations between 418 B. napus and ‘Darmor-bzh' reference genomes; 21, The common loci between PA and PY-related traits identified in rapeseed; 23, Summary of rapeseed accessions used for qRT–PCR of different traits in this study
Rights and permissions
About this article

Cite this article
Hu, J., Chen, B., Zhao, J. et al. Genomic selection and genetic architecture of agronomic traits during modern rapeseed breeding. Nat Genet 54, 694–704 (2022). https://doi.org/10.1038/s41588-022-01055-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-022-01055-6
This article is cited by
-
Genome-wide characterization of SDR gene family and its potential role in seed dormancy of Brassica napus L.
BMC Plant Biology (2024)
-
Effects of marker density on genomic prediction for yield traits in sweet corn
Euphytica (2024)
-
High-throughput phenotyping of individual plant height in an oilseed rape population based on Mask-RCNN and UAV images
Precision Agriculture (2024)
-
Genetic dissection of ten photosynthesis-related traits based on InDel- and SNP-GWAS in soybean
Theoretical and Applied Genetics (2024)
-
Downregulation of the expression of subgenomic chromosome A7 genes promotes plant height in resynthesized allopolyploid Brassica napus
Theoretical and Applied Genetics (2024)
