
Abstract
Using online surveys, we collected data regarding COVID-19-related loss of smell or taste from 69,841 individuals. We performed a multi-ancestry genome-wide association study and identified a genome-wide significant locus in the vicinity of the UGT2A1 and UGT2A2 genes. Both genes are expressed in the olfactory epithelium and play a role in metabolizing odorants. These findings provide a genetic link to the biological mechanisms underlying COVID-19-related loss of smell or taste.
Similar content being viewed by others

Main
Loss of sense of smell (anosmia) or taste (ageusia) are distinctive symptoms of COVID-19 and are among the earliest and most often reported indicators of the acute phase of SARS-CoV-2 infection. It is notable from other viral symptoms in its sudden onset and the absence of mucosal blockage1. While a large fraction of COVID-19 patients report loss of smell or taste, the underlying mechanism is unclear2. In this study, we conducted a genome-wide association study (GWAS) of COVID-19-related loss of smell or taste, having collected self-reported data from over 1 million 23andMe research participants as described previously3. By asking study participants to report the symptoms they encountered during their COVID-19 experience, we identified SARS-CoV-2 test-positive individuals who reported a loss of smell or taste and contrasted them with test-positive individuals who did not report a loss of smell or taste.
Of the individuals who self-reported having received a SARS-CoV-2 positive test, 68% reported loss of smell or taste as a symptom (47,298 out of a total of 69,841 individuals). Female respondents were more likely than male respondents to report this symptom (72% versus 61%; chi-squared test, P = 5.7 × 10−178) and those with this symptom were typically younger than those without this symptom (mean age of 41 years for those with loss of smell or taste versus 45 years for those without; P = 2.34 × 10−199, Welch’s t-test). Among genetically determined ancestral groups, rates of loss of smell or taste ranged between 63% and 70% (Table 1). As expected, compared to other symptoms surveyed, loss of smell or taste was much more common among those with a SARS-CoV-2 positive test compared to those who self-reported other cold or flu-like symptoms but who tested negative for SARS-CoV-2 (Extended Data Fig. 1). In a logistic regression model predicting loss of smell or taste as a function of age, sex and genetic ancestry, individuals of East Asian or African American ancestry were significantly less likely to report loss of smell or taste (odds ratio (OR) = 0.8 and 0.88, respectively) relative to individuals of European ancestry (Supplementary Table 1).
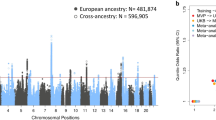
For unrelated individuals with complete data, we conducted GWAS within each ancestry group separately (total sample size = 56,373) before performing a multi-ancestry meta-analysis using a fixed effects model. Each input GWAS was adjusted for inflation via genomic control (λ = 1.029, 1.037, 1.024, 1.042 and 1.071 within the European, Latino, African American, East Asian and South Asian ancestry GWAS, respectively), as was the subsequent meta-analysis (λ = 1.001). Within the multi-ancestry meta-analysis, we identified a single associated locus at chr4q13.3 (Fig. 1). No other locus achieved genome-wide significance in the multi-ancestry meta-analysis or in any of the input populations. The index SNP at this locus was rs7688383 (C/T, with T being the risk allele, P = 1.4 × 10−14, OR = 1.11). While most of the support for this genetic association within the multi-ancestry analysis comes from the European population (for which we have the largest sample size), the estimated effect sizes are consistent across populations (Supplementary Table 2). The credible set from the multi-ancestry analysis contained 28 variants covering a 44.6-kilobase (kb) region (chr4:69.57–69.62 megabases (Mb); Supplementary Table 3).

a, Manhattan plot. SNPs achieving genome-wide significance are highlighted in red. The nearest gene to the index SNP is indicated above the relevant association peak. b, Regional plot around the UGT2A1/UGT2A2 locus. The colors indicate the strength of linkage disequilibrium (r2) relative to the index SNP (rs7688383). Imputed variants are indicated with ‘+’ symbols; coding variants are indicated with ‘x’ symbols. Where imputed variants were not available, directly genotyped variants are indicated by ‘o’ symbols; coding variants are indicated by diamond symbols.
We performed a phenome-wide association study on the index SNP across approximately 1,300 phenotypes defined in the 23andMe database. We identified four additional associated phenotypes with P < 1 × 10−6, of which two are related to the ability to smell, one is related to ice cream taste preference and one is related to tobacco use (Supplementary Table 4). We detected no other associations with COVID-19 symptoms, susceptibility or severity.
There are four genes within 150 kb of the association (UGT2A1, UGT2A2, UGT2B4, SULT1B1), with the index variant itself being within an intron of the overlapping UGT2A1 and UGT2A2 genes. While the GWAS index variant appears to be physically proximal to expression quantitative trait loci (eQTLs) for UGT2A1, evidence for colocalization between the GWAS and eQTLs for any of the nearby genes is generally weak (Supplementary Note and Extended Data Figs. 2 and 3). Nonetheless, of the four genes in the vicinity, UGT2A1 and UGT2A2 are not only the most proximal but also the most biologically plausible causal gene candidates. UGT2A1 and UGT2A2 are part of a family of uridine diphosphate glycosyltransferases, enzymes that metabolize lipophilic substrates through conjugation with glucoronic acid. During olfaction, animal studies show that these enzymes, which are expressed in the olfactory epithelium, are involved in the elimination of the odorants that enter the nasal cavity and bind to olfactory receptors. For example, glucuronidation of odorants fails to stimulate the olfactory bulb, which prevents the odor from being detected by the brain, functionally demonstrating the effect of the enzyme produced by these genes on the odorant4. This results in the clearance of the odorant to facilitate the transient experience of olfaction, once the stimuli are no longer present in the environment5. UGT2A2 is a splice variant of UGT2A1, with identical C-terminal residues but different N termini6. Conversely, UGT2B4 and SULT1B1 appear less plausible from a biological perspective, with neither having a clear link to olfactory or gustatory function.
While mechanistic explanations have been proposed7 for COVID-19-related loss of smell, experimental studies suggest that loss of smell is related to damage to the cilia and olfactory epithelium but not infection of the olfactory neurons. For example, in an experiment where hamsters were nasally infected with SARS-CoV-2, the olfactory epithelium and cilia became very damaged, which can completely inhibit the ability to smell, but no infection was observed in the olfactory neurons8. Recent evidence suggests that SARS-CoV-2 enters and accumulates in olfactory support cells, specifically, sustentacular cells, which unlike olfactory neurons abundantly express the viral entry proteins angiotensin-converting enzyme 2 (ACE2) and transmembrane protease serine 2 (TMPRSS2; refs. 9,10). These support cells are metabolically and functionally associated with olfactory neurons and with odorant signal transduction (processing odorants by endocytosing the odorant-binding protein complex, detoxifying, maintaining the cilia of mature olfactory receptor neurons and maintaining epithelial integrity). It has been proposed that olfactory sensation is impaired when these essential functions are disrupted, causing ciliary impairment7. How UGT2A1 and UGT2A2 are involved in this process is unclear but given their localization and essential function in metabolizing and detoxifying such compounds, these genes may play a role in the physiology of infected cells and the resulting functional impairment that contributes to loss of ability to smell. Notably, the variant identified in this study also appears to be associated with general ability to smell, which may suggest that those with heightened smell or taste sensitivity may be more prone to notice a loss of these senses resulting from a SARS-CoV-2 infection.
Our study has several limitations. First, while our study was large in scale, it was biased toward individuals of European ancestry and lacked a replication cohort. Second, we relied on self-reported case and symptom status; replication within a cohort with clinical ascertainment could be beneficial. Third, given that loss of smell or taste were combined in a single survey question, we cannot further disentangle these two symptoms. Loss of smell without loss of taste may be distinct from loss of both or loss of taste without loss of smell. Given this, it is unclear if our findings relate more strongly to one symptom or the other.
Methods
Overview of study recruitment and data collection
Participants in this study were recruited from the customer base of 23andMe, a personal genetics company. Participants provided informed consent and participated in the research online, under a protocol approved by the external Association for the Accreditation of Human Research Protection Programs-accredited institutional review board, Ethical and Independent Review Services. Participants were included in the analysis based on consent status as checked at the time data analyses were initiated.
Full details of the data collection paradigm for this study have been described previously3. In brief, primary recruitment was carried out by email to approximately 6.7 million 23andMe research participants over 18 years of age and living in the USA or UK. Additionally, pre-existing customers were invited to participate in the study through promotional materials on the 23andMe website, the 23andMe mobile application and via social media. Study participation consisted solely of web-based surveys, including an initial baseline survey and three follow-up surveys fielded each month after completion of the baseline survey. The surveys collected information regarding individuals’ experiences with COVID-19 and included questions regarding recently experienced symptoms with or without a SARS-CoV-2 positive test. Enrollment continued after the initial recruitment efforts until a data freeze was taken for this study in March 2021, when 1.3 million participants had completed the baseline survey.
Phenotype definition for GWAS
Using the information derived from the surveys, we defined a phenotype to contrast SARS-CoV-2 positive individuals that experienced COVID-19-related loss of smell or taste from those who did not. Specifically, participants were asked to respond to the question ‘Have you been tested for COVID-19 (coronavirus)?’, with possible responses ‘Yes, it was positive/Yes, it was negative/No/My results are pending/I’m not sure’. Of those who responded ‘Yes, it was positive’, we further considered the question ‘During your illness, did you experience any of the following symptoms?’, to which participants could select as many as needed from the following list of responses: ‘Muscle or body aches/Fatigue/Dry cough/Sore throat/Coughing up of sputum or phlegm (productive cough)/Loss of smell or taste/Chills/Difficulty breathing or shortness of breath/Pressure or tightness in upper chest/Diarrhea/Nausea or vomiting/Sneezing/Loss of appetite/Runny nose/Headache/Intensely red or watery eyes’. We defined cases as SARS-CoV-2 test-positive individuals who also reported ‘Loss of smell or taste’, and controls as SARS-CoV-2 test-positive individuals who did not report ‘Loss of smell or taste’. While some participants reported a COVID-19 diagnosis absent a confirmed positive test for SARS-CoV-2, we did not include such individuals within this analysis.
Descriptive statistics
Sample sizes and proportions were calculated by age, sex and ancestry. Differences in loss of smell or taste by sex were statistically evaluated with a chi-squared statistics and mean differences in age were evaluated with a t-test. A logistic regression model was constructed to evaluate loss of smell or taste as a function of ancestry, age (categorical) and sex. All analyses were conducted in R v.3.6.3.
Genotyping and SNP imputation
DNA extraction and genotyping were performed on saliva samples by Clinical Laboratory Improvement Amendments-certified and College of American Pathologists-accredited clinical laboratories of Laboratory Corporation of America. Samples were genotyped on one of five genotyping platforms. The V1 and V2 platforms were variants of the Illumina HumanHap550 BeadChip and contained a total of about 560,000 SNPs, including about 25,000 custom SNPs selected by 23andMe. The V3 platform was based on the Illumina OmniExpress BeadChip and contained a total of about 950,000 SNPs and custom content to improve the overlap with our V2 array. The V4 platform was a fully custom array of about 950,000 SNPs and included a lower redundancy subset of V2 and V3 SNPs with additional coverage of lower-frequency coding variation. The V5 platform was based on the Illumina Global Screening Array, consisting of approximately 654,000 preselected SNPs and approximately 50,000 custom content variants. Samples that failed to reach 98.5% call rate were reanalyzed. Individuals whose analyses failed repeatedly were recontacted by the 23andMe customer service to provide additional samples as done for all 23andMe customers.
Participant genotype data were imputed using the Haplotype Reference Consortium (HRC) panel11, augmented by the phase 3 1000 Genomes Project panel12 for variants not present in the HRC. We phased and imputed data for each genotyping platform separately. For the non-pseudoautosomal region of the X chromosome, males and females were phased together in segments, treating males as already phased; the pseudoautosomal regions were phased separately. We then imputed males and females together, treating males as homozygous pseudo-diploids for the non-pseudoautosomal region.
GWAS
Genotyped participants were included in the GWAS analyses on the basis of ancestry as determined by a genetic ancestry classification algorithm13. We selected a set of unrelated individuals so that no 2 individuals shared more than 700 cM of DNA identical by descent (IBD). If a case and a control were identified as having at least 700 cM of DNA IBD, we preferentially discarded the control from the sample. This filtering paradigm resulted in approximately 1.76% of the sample being excluded.
We tested for association using logistic regression, assuming additive allelic effects. For tests using imputed data, we used the imputed dosages rather than best-guess genotypes. We included covariates for age, age squared, sex, a sex:age interaction, the top ten principal components to account for residual population structure and dummy variables to account for the genotyping platform. The association test P value was computed using a likelihood ratio test, which in our experience is better behaved than a Wald test on the regression coefficient. Results for the X chromosome were computed similarly, with men coded as if they were homozygous diploid for the observed allele.
We combined the GWAS summary statistics from both genotyped and imputed data. When choosing between imputed and genotyped GWAS results, we favored the imputed result, unless the imputed variant was unavailable or failed quality control. For imputed variants, we removed variants with low imputation quality (r2 < 0.5 averaged across batches or a minimum r2 < 0.3) or with evidence of batch effects (analysis of variance (ANOVA) F-test across batches, P < 10−50). For genotyped variants, we removed variants only present on our V1 or V2 arrays (due to small sample size) that failed a Mendelian transmission test in trios (P < 10−20), failed a Hardy–Weinberg test in individuals of European ancestry (P < 10−20), failed a batch effect test (ANOVA P < 10−50) or had a call rate <90%.
We repeated the GWAS analysis separately in each population cohort for which we had sufficient data (European, Latino, African American, East Asian and South Asian ancestry); the resulting summary statistics were adjusted for inflation using genomic control when the inflation factor was estimated to be greater than 1. We then performed multi-ancestry meta-analysis using a fixed effects model (inverse variance method14), restricting to variants of at least 1% minor allele frequency in the pooled sample and minor allele count > 30 within each subpopulation. Both the input GWAS and resulting meta-analysis were adjusted for inflation using genomic control where necessary.
We identified regions with genome-wide significant associations. We defined the region boundaries by identifying all SNPs with P < 10−5 within the vicinity of a genome-wide significance association and then grouping these regions into intervals so that no 2 regions were separated by less than 250 kb. We considered the SNP with the smallest P value within each interval to be the index SNP. Within each region, we calculated a credible set using the method of Maller et al.15.
Reporting Summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The full set of de-identified summary statistics can be made available to qualified investigators who enter into an agreement with 23andMe that protects participant confidentiality. Interested investigators should visit https://research.23andme.com/covid19-dataset-access/.
References
Parma, V. et al. More than smell—COVID-19 is associated with severe impairment of smell, taste, and chemesthesis. Chem. Senses 45, 609–622 (2020).
Mutiawati, E. et al. Anosmia and dysgeusia in SARS-CoV-2 infection: incidence and effects on COVID-19 severity and mortality, and the possible pathobiology mechanisms—a systematic review and meta-analysis. F1000Res. 10, 40 (2021).
Shelton, J. F. et al. Trans-ancestry analysis reveals genetic and nongenetic associations with COVID-19 susceptibility and severity. Nat. Genet. 53, 801–808 (2021).
Neiers, F., Jarriault, D., Menetrier, F., Briand, L. & Heydel, J.-M. The odorant metabolizing enzyme UGT2A1: immunolocalization and impact of the modulation of its activity on the olfactory response. PLoS ONE 16, e0249029 (2021).
Lazard, D. et al. Odorant signal termination by olfactory UDP glucuronosyl transferase. Nature 349, 790–793 (1991).
Mackenzie, P. I. et al. Nomenclature update for the mammalian UDP glycosyltransferase (UGT) gene superfamily. Pharmacogenet. Genomics 15, 677–685 (2005).
Butowt, R. & von Bartheld, C. S. Anosmia in COVID-19: underlying mechanisms and assessment of an olfactory route to brain infection. Neuroscientist 27, 582–603 (2021).
Bryche, B. et al. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav. Immun. 89, 579–586 (2020).
Brann, D. H. et al. Non-neuronal expression of SARS-CoV-2 entry genes in the olfactory system suggests mechanisms underlying COVID-19-associated anosmia. Sci. Adv. 6, eabc5801 (2020).
Bilinska, K., Jakubowska, P., Von Bartheld, C. S. & Butowt, R. Expression of the SARS-CoV-2 entry proteins, ACE2 and TMPRSS2, in cells of the olfactory epithelium: identification of cell types and trends with age. ACS Chem. Neurosci. 11, 1555–1562 (2020).
McCarthy, S. et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 48, 1279–1283 (2016).
Auton, A. et al. A global reference for human genetic variation. Nature 526, 68–74 (2015).
Durand, E. Y., Do, C. B., Mountain, J. L. & Macpherson, J. M. Ancestry Composition: a novel, efficient pipeline for ancestry deconvolution. Preprint at bioRxiv https://doi.org/10.1101/010512 (2014).
Willer, C. J., Li, Y. & Abecasis, G. R. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics 26, 2190–2191 (2010).
Maller, J. B. et al. Bayesian refinement of association signals for 14 loci in 3 common diseases. Nat. Genet. 44, 1294–1301 (2012).
Acknowledgements
We thank the 23andMe research participants and employees who made this study possible. The 23andMe Research Team is: S. Aslibekyan, A. Auton, E. Babalola, R. K. Bell, J. Bielenberg, K. Bryc, E. Bullis, D. Coker, G. Cuellar Partida, D. Dhamija, S. Das, S. L. Elson, T. Filshtein, K. Fletez-Brant, P. Fontanillas, W. Freyman, P. M. Gandhi, K. Heilbron, B. Hicks, D. A. Hinds, E. M. Jewett, Y. Jiang, K. Kukar, K.-H. Lin, M. Lowe, J. McCreight, M. H. McIntyre, S. J. Micheletti, M. E. Moreno, J. L. Mountain, P. Nandakumar, E. S. Noblin, J. O’Connell, A. A. Petrakovitz, G. D. Poznik, M. Schumacher, A. J. Shastri, J. F. Shelton, J. Shi, S. Shringarpure, V. Tran, J. Y. Tung, X. Wang, W. Wang, C. H. Weldon, P. Wilton, A. Hernandez, C. Wong and C. Toukam Tchakouté.
Author information
Authors and Affiliations
Consortia
Contributions
J.F.S., A.J.S., S.A. and A.A. designed this study. The 23andMe COVID-19 Team developed the recruitment and participant engagement strategy and acquired and processed the data. J.F.S., K.F.-B. and A.A. analyzed the data. J.F.S., A.J.S., K.F.-B. and A.A. interpreted the data. J.F.S., A.J.S. and A.A. wrote the manuscript. All authors participated in the preparation of the manuscript by reading and commenting on the drafts before submission.
Corresponding author
Ethics declarations
Competing interests
J.F.S., A.J.S., K.F.-B., S.A. and A.A. are current employees of 23andMe and hold stock or stock options in 23andMe.
Peer review information
Nature Genetics thanks Patrick Sulem and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Self-reported symptoms experienced during SARS-CoV-2 infection with a positive test (n = 69,841) as compared to individuals self-reporting cold or flu-like illness but with a negative SARS-CoV-2 test (n = 314,441).
Loss of smell or taste was reported by 68% of individuals with a positive test for SARS-CoV-2 infection.
Extended Data Fig. 2 Conditional association LocusZoom plots for rs768838.
Lack of evidence for conditional associations. Left, LocusZoom plot of primary association in the European population prior to conditional analysis. Right, LocusZoom plot of the same region having included rs7688383 in the regression model.
Extended Data Fig. 3 Examples of eQTL associations for UTG2A1, UGT2B4, and SULT1B1.
eQTL association plots for UGT2A1 (left), UGT2B4 (middle), and SULT1B1 (right). No eQTL associations were observed for UGT2A2. For each gene, the three tissues with the strongest eQTL associations are shown. Colors represent the linkage disequilibrium with the GWAS index SNP (rs7688383).
Supplementary information
Supplementary Information
Supplementary Note and Tables 1–4.
Rights and permissions
About this article

Cite this article
Shelton, J.F., Shastri, A.J., Fletez-Brant, K. et al. The UGT2A1/UGT2A2 locus is associated with COVID-19-related loss of smell or taste. Nat Genet 54, 121–124 (2022). https://doi.org/10.1038/s41588-021-00986-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-021-00986-w
This article is cited by
-
TAS2R38 haplotypes, COVID-19 infection, and symptomatology: a cross-sectional analysis of data from the Canadian Longitudinal Study on Aging
Scientific Reports (2024)
-
Next-generation sequencing of host genetics risk factors associated with COVID-19 severity and long-COVID in Colombian population
Scientific Reports (2024)
-
Emergence and influence of sequence bias in evolutionarily malleable, mammalian tandem arrays
BMC Biology (2023)
-
Mechanism and treatment of olfactory dysfunction caused by coronavirus disease 2019
Journal of Translational Medicine (2023)
-
Olfactory immune response to SARS-CoV-2
Cellular & Molecular Immunology (2023)
