
Abstract
Characterizing the stem cells responsible for lung repair and regeneration is important for the treatment of pulmonary diseases. Recently, a unique cell population located at the bronchioalveolar-duct junctions has been proposed to comprise endogenous stem cells for lung regeneration. However, the role of bronchioalveolar stem cells (BASCs) in vivo remains debated, and the contribution of such cells to lung regeneration is not known. Here we generated a genetic lineage-tracing system that uses dual recombinases (Cre and Dre) to specifically track BASCs in vivo. Fate-mapping and clonal analysis showed that BASCs became activated and responded distinctly to different lung injuries, and differentiated into multiple cell lineages including club cells, ciliated cells, and alveolar type 1 and type 2 cells for lung regeneration. This study provides in vivo genetic evidence that BASCs are bona fide lung epithelial stem cells with deployment of multipotency and self-renewal during lung repair and regeneration.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others
Data availability
The generated sequencing data have been deposited in the GEO database under accession code GSE118891.
Change history
07 March 2019
In the version of this article initially published, the following grant numbers and recipients were missing from the Acknowledgements: XDB19000000 to H.J. and B.Z.; 81430066 and 31621003 to H.J.; 2017YFA0505500 to H.J.; and 15XD1504000 to H.J. The errors have been corrected in the HTML and PDF versions of the article.
References
Kuo, C. S. & Krasnow, M. A. Formation of a neurosensory organ by epithelial cell slithering. Cell 163, 394–405 (2015).
Morrisey, E. E. & Hogan, B. L. Preparing for the first breath: genetic and cellular mechanisms in lung development. Dev. Cell 18, 8–23 (2010).
Rock, J. R. & Hogan, B. L. Epithelial progenitor cells in lung development, maintenance, repair, and disease. Annu. Rev. Cell Dev. Biol. 27, 493–512 (2011).
Hogan, B. L. et al. Repair and regeneration of the respiratory system: complexity, plasticity, and mechanisms of lung stem cell function. Cell Stem Cell 15, 123–138 (2014).
Desai, T. J., Brownfield, D. G. & Krasnow, M. A. Alveolar progenitor and stem cells in lung development, renewal and cancer. Nature 507, 190–194 (2014).
Kumar, P. A. et al. Distal airway stem cells yield alveoli in vitro and during lung regeneration following H1N1 influenza infection. Cell 147, 525–538 (2011).
Chen, F. & Krasnow, M. A. Progenitor outgrowth from the niche in Drosophila trachea is guided by FGF from decaying branches. Science 343, 186–189 (2014).
Zuo, W. et al. p63+Krt5+ distal airway stem cells are essential for lung regeneration. Nature 517, 616–620 (2014).
Peng, T. et al. Hedgehog actively maintains adult lung quiescence and regulates repair and regeneration. Nature 526, 578–582 (2015).
Hong, K. U., Reynolds, S. D., Watkins, S., Fuchs, E. & Stripp, B. R. Basal cells are a multipotent progenitor capable of renewing the bronchial epithelium. Am. J. Pathol. 164, 577–588 (2004).
Hong, K. U., Reynolds, S. D., Watkins, S., Fuchs, E. & Stripp, B. R. In vivo differentiation potential of tracheal basal cells: evidence for multipotent and unipotent subpopulations. Am. J. Physiol. Lung Cell. Mol. Physiol. 286, L643–L649 (2004).
Rock, J. R. et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc. Natl Acad. Sci. USA 106, 12771–12775 (2009).
Rawlins, E. L. et al. The role of Scgb1a1+ Clara cells in the long-term maintenance and repair of lung airway, but not alveolar, epithelium. Cell Stem Cell 4, 525–534 (2009).
Hong, K. U., Reynolds, S. D., Giangreco, A., Hurley, C. M. & Stripp, B. R. Clara cell secretory protein-expressing cells of the airway neuroepithelial body microenvironment include a label-retaining subset and are critical for epithelial renewal after progenitor cell depletion. Am. J. Respir. Cell Mol. Biol. 24, 671–681 (2001).
Barkauskas, C. E. et al. Type 2 alveolar cells are stem cells in adult lung. J. Clin. Invest. 123, 3025–3036 (2013).
Zacharias, W. J. et al. Regeneration of the lung alveolus by an evolutionarily conserved epithelial progenitor. Nature 555, 251–255 (2018).
Nabhan, A., Brownfield, D. G., Harbury, P. B., Krasnow, M. A. & Desai, T. J. Single-cell Wnt signaling niches maintain stemness of alveolar type 2 cells. Science 359, 1118–1123 (2018).
Vaughan, A. E. et al. Lineage-negative progenitors mobilize to regenerate lung epithelium after major injury. Nature 517, 621–625 (2014).
Tata, P. R. et al. Dedifferentiation of committed epithelial cells into stem cells in vivo. Nature 503, 218–223 (2013).
Giangreco, A., Reynolds, S. D. & Stripp, B. R. Terminal bronchioles harbor a unique airway stem cell population that localizes to the bronchoalveolar duct junction. Am. J. Pathol. 161, 173–182 (2002).
Kim, C. F. et al. Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell 121, 823–835 (2005).
Nolen-Walston, R. D. et al. Cellular kinetics and modeling of bronchioalveolar stem cell response during lung regeneration. Am. J. Physiol. Lung Cell. Mol. Physiol. 294, L1158–L1165 (2008).
Dovey, J. S., Zacharek, S. J., Kim, C. F. & Lees, J. A. Bmi1 is critical for lung tumorigenesis and bronchioalveolar stem cell expansion. Proc. Natl Acad. Sci. USA 105, 11857–11862 (2008).
Zacharek, S. J. et al. Lung stem cell self-renewal relies on BMI1-dependent control of expression at imprinted loci. Cell Stem Cell 9, 272–281 (2011).
Lee, J. H. et al. Lung stem cell differentiation in mice directed by endothelial cells via a bmp4-nfatc1-thrombospondin-1 axis. Cell 156, 440–455 (2014).
He, L. et al. Enhancing the precision of genetic lineage tracing using dual recombinases. Nat. Med. 23, 1488–1498 (2017).
Zhang, H. et al. Genetic lineage tracing identifies endocardial origin of liver vasculature. Nat. Genet. 48, 537–543 (2016).
Madisen, L. et al. Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance. Neuron 85, 942–958 (2015).
Madisen, L. et al. A robust and high-throughput cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 13, 133–140 (2010).
Zhang, H. et al. Endocardium contributes to cardiac fat. Circ. Res. 118, 254–265 (2016).
Reinert, R. B. et al. Tamoxifen-induced Cre-loxP recombination is prolonged in pancreatic islets of adult mice. PLoS One 7, e33529 (2012).
Rock, J. R. et al. Multiple stromal populations contribute to pulmonary fibrosis without evidence for epithelial to mesenchymal transition. Proc. Natl Acad. Sci. USA 108, E1475–E1483 (2011).
Snippert, H. J. et al. Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell 143, 134–144 (2010).
Montoro, D. T. et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 12771, 319–324 (2018).
Plasschaert, L. W. et al. A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte. Nature 545, 377–381 (2018).
Honda, H. et al. Leucine-rich α-2 glycoprotein promotes lung fibrosis by modulating TGF-β signaling in fibroblasts. Physiol. Rep. 5, e13556 (2017).
Wang, X. et al. LRG1 promotes angiogenesis by modulating endothelial TGFβ signalling. Nature 499, 306–311 (2013).
Schultz, C. J., Torres, E., Londos, C. & Torday, J. S. Role of adipocyte differentiation-related protein in surfactant phospholipid synthesis by type II cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 283, L288–L296 (2002).
El Agha, E. et al. Two-way conversion between lipogenic and myogenic fibroblastic phenotypes marks the progression and resolution of lung fibrosis. Cell Stem Cell 20, 261–273.e3 (2016).
Zhang, X. et al. Comparative membrane proteomic analysis between lung adenocarcinoma and normal tissue by iTRAQ labeling mass spectrometry. Am. J. Transl. Res. 6, 267–280 (2014).
Zhang, X. D. et al. Identification of adipophilin as a potential diagnostic tumor marker for lung adenocarcinoma. Int. J. Clin. Exp. Med. 7, 1190–1196 (2014).
Xu, X. et al. Evidence for type II cells as cells of origin of K-Ras-induced distal lung adenocarcinoma. Proc. Natl Acad. Sci. USA 109, 4910–4915 (2012).
Noh, M. S. et al. Magnetic surface-enhanced Raman spectroscopic (M-SERS) dots for the identification of bronchioalveolar stem cells in normal and lung cancer mice. Biomaterials 30, 3915–3925 (2009).
Kotton, D. N. & Morrisey, E. E. Lung regeneration: mechanisms, applications and emerging stem cell populations. Nat. Med. 20, 822–832 (2014).
Pardo-Saganta, A. et al. Parent stem cells can serve as niches for their daughter cells. Nature 523, 597–601 (2015).
Jain, R. et al. Plasticity of Hopx+ type I alveolar cells to regenerate type II cells in the lung. Nat. Commun. 6, 6727 (2015).
Giangreco, A. et al. Stem cells are dispensable for lung homeostasis but restore airways after injury. Proc. Natl Acad. Sci. USA 106, 9286–9291 (2009).
Tian, X. et al. Subepicardial endothelial cells invade the embryonic ventricle wall to form coronary arteries. Cell Res. 23, 1075–1090 (2013).
Liu, Q. et al. Genetic targeting of sprouting angiogenesis using Apln-CreER. Nat. Commun. 6, 6020 (2015).
Degryse, A. L. et al. Repetitive intratracheal bleomycin models several features of idiopathic pulmonary fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 299, L442–L452 (2010).
He, L. et al. Preexisting endothelial cells mediate cardiac neovascularization after injury. J. Clin. Invest. 127, 2968–2981 (2017).
Liu, Q. et al. Genetic lineage tracing identifies in situ Kit-expressing cardiomyocytes. Cell Res. 26, 119–130 (2016).
Chen, J. et al. Spatial transcriptomic analysis of cryosectioned tissue samples with Geo-seq. Nat. Protoc. 12, 566–580 (2017).
Picelli, S. et al. Smart-seq2 for sensitive full-length transcriptome profiling in single cells. Nat. Methods 10, 1096–1098 (2013).
Pertea, M., Kim, D., Pertea, G. M., Leek, J. T. & Salzberg, S. L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 11, 1650–1667 (2016).
Anders, S., Pyl, P. T. & Huber, W. HTSeq: a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Kiselev, V. Y. et al. SC3: consensus clustering of single-cell RNA-seq data. Nat. Methods 14, 483–486 (2017).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Acknowledgements
This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (CAS, XDB19000000 to H.J. and B.Z.; XDA16020204 to B.Z.; XDA16020404 to G.P.; and XDA16020501 to N.J.), the National Science Foundation of China (31730112, 91639302, 31625019, 91849202 and 81761138040 to B.Z.; 31601168 to Q.L.; 31701292 and 81872241 to L.H.; and 31571503, 91749122 and 81872132 to X.T.; 81430066 and 31621003 to H.J.), the National Key Research and Development Program of China (2018YFA0107900 and 2016YFC1300600 to X.T.; 2018YFA0108100 and 2017YFC1001303 to L.H.; 2017YFA0505500 to H.J.), the Key Project of Frontier Sciences of CAS (QYZDB-SSW-SMC003), the Shanghai Science and Technology Commission (17ZR1449600 to B.Z., 17ZR1449800 to X.T., 15XD1504000 to H.J. and 15XD1504000 to B.Z.), the Shanghai Yangfan Project (16YF1413400 to L.H.), the China Postdoctoral Innovative Talent Support Program (BX20180338 to Y.L.), China Young Talents Lift Engineering (YESS20160050 to Q.L. and 2017QNRC001 to L.H.), the collaboration fund of Research Beyond Borders at Boehringer Ingelheim Pharma GmbH (B.Z.), Astrazeneca (B.Z.) and a Royal Society-Newton Advanced Fellowship (B.Z., NA170109) and the Program for Guangdong Introducing Innovative and Entrepreneurial Teams (2017ZT07S347 to B.Z.). We thank the Shanghai Model Organisms Center, Inc. (SMOC) and Nanjing Biomedical Research Institute of Nanjing University for mouse generation. We also acknowledge technical help from L. Qiu, W. Bian, T. Zhang and members of National Center for Protein Science Shanghai for assistance in flow cytometry and microscopy and Y. Xing for antibody sharing. We thank P. Nicklin, M. Franti and W. Zhang for valuable suggestions and comments on this study. We also thank the Genome Tagging Project (GTP) Center for support.
Author information
Authors and Affiliations
Contributions
Q.L., K.L. and B.Z. designed the study and wrote the manuscript. Q.L., K.L. and G.C. performed experiments and analyzed the data. W.G., G.C., G.P. and N.J. performed scRNA-seq and analyzed data. X.H., S.Y., Z.Q., Y.L., R.Y., W.P., L.Z., L.H., H.Z., W.Y., M.T., X.T., D.C., Y.N., S.H., T.R., Z.Q., H.H. and Y.A.Z. bred the mice, performed experiments or provided material, important suggestions and valuable comments. H.J. designed the study and provided valuable comments. B.Z. supervised the study and analyzed the data.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Scgb1a1-CreER targets mainly CC10+ club cells.
(a) Strategy for labeling of CC10+ cells by Cre-loxP system. (b) Whole-mount bright-field and fluorescence view of lung from adult Scgb1a1-CreER;R26-tdTomato mice 1 week post Tamoxifen induction. (c-f) Immunostaining for tdTomato and lineage markers CC10, CK5, SPC, aSMA or VE-cad on lung sections. White arrowheads in c indicate weak expression of CC10 in some tdTomato+ cells at BADJ regions. Boxed region are magnified on the right. Scale bars, yellow, 1 mm; white 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 2 Cell labeling by Scgb1a1-CreER;R26-tdTomato.
(a) Immunostaining for tdTomato and SPC on lung sections collected from Scgb1a1-CreER;R26-tdTomato mice treated with 0.2 mg/g tamoxifen. In addition to club cell labeling, Scgb1a1-CreER also labels a subset of AT2 cells (arrowheads). (b) Immunostaining for CC10, SPC and tdTomato on lung tissue sections from Scgb1a1-CreER;R26-tdTomato mice treated with 0.025 mg/g tamoxifen. A significant fewer tdTomato+ AT2 cells (arrowhead) were detected in 0.025 mg/g tamoxifen treated mice (b) compared with 0.2 mg/g tamoxifen treated mice (a). In 0.025 mg/g tamoxifen treated samples, tdTomato+ BASCs (arrows) could also be detected. Scale bars, 100 µm. Each figure is representative of 5 individual biological samples.
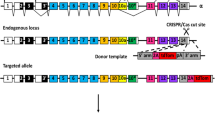
Supplementary Figure 3 Sftpc-DreER targets mainly SPC+ AT2 cells.
(a) Schematic figure showing knock-in strategy for Sftpc-DreER allele by homologous recombination. (b) Strategy for labeling of SPC+ cells by Dre-rox recombination. (c) Whole-mount fluorescence image showing tdTomato labeling of lung from Sftpc-DreER;R26-RSR-tdTomato mouse at 1 week after tamoxifen treatment. Dotted lines mark trachea. (d-h) Immunostaining for tdTomato and cell lineage markers SPC, CC10, CK5, T1a, aSMA and VE-cad on Sftpc-DreER;R26-RSR-tdTomato lung. Boxed regions are magnified on the right. XZ and YZ indicate signals from dotted lines on Z-stack image. Scale bars, yellow, 1 mm; white, 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 4 Sftpc-DreER labels most AT2 cells (CC10–SPC+) and BASCs (CC10+SPC+), and very few club cells (CC10+SPC–).
(a-d) Immunostaining for tdTomato, CC10 and SPC on lung tissue sections collected from Sftpc-DreER;R26-RSR-tdTomato mice at 1week after tamoxifen induction (0.2 mg/g). The labeled BASCs (arrowheads) were located at BADJs. Boxed regions in a are magnified in b,c,d. Scale bars, 100 µm. Each figure is representative of 5 individual biological samples.
Supplementary Figure 5 BASCs-Tracer does not label AT1 cells, ciliated cells or neuroendocrine cells.
(a-c) Immunostaining for tdTomato and T1a (AT1 cell marker), β-tubulin (ciliated cell marker) or CGRP (neuroendocrine cell marker) on BASCs-Tracer mouse lung sections. Tamoxifen was induced one week before analysis. Scale bars, 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 6 BASCs-Tracer does not label K5+ or P63+ cells.
(a-d) Immunostaining for tdTomato and K5 (a) or P63 (c) on lung tissue sections. Tamoxifen was induced at 1 week before tissue collection. K5 and P63 could be detected in the lung trachea (b) or stomach (d) tissue sections of mouse. Scale bars, 100 µm. Each image is a representative of 4 biologically independent sample.
Supplementary Figure 7 Scgb1a1-CreER or Sftpc-DreER rarely labels cells by R26-RSR-LSL-tdTomato reporter.
(a,b) Schematic figure showing crossing of Scgb1a1-CreER or Sftpc-DreER with R26-RSR-LSL-tdTomato reporter respectively. Tamoxifen was injected one week before tissue analysis. (c,d) Whole-mount fluorescence and bright-field view of lungs from Scgb1a1-CreER;R26-RSR-LSL-tdTomato mouse (c) or Sftpc-DreER;R26-RSR-LSL-tdTomato mouse (d). (e-l) Immunostaining for tdTomato and cell lineage markers T1a, SPC, CC10 and CK5. Boxed region are magnified on the right. Scale bars, yellow, 1 mm; white, 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 8 Negligible leakiness of BASCs-Tracer mouse without tamoxifen induction.
(a,b) Immunostaining for tdTomato, SPC and CC10 on lung sections of BASCs-Tracer mouse. Boxed region is magnified in the below images. Most fields in the lung sections are negative for tdTomato (a), with only very few fields that contain sparse tdTomato+ cells (b). The extremely rare tdTomato+ cell is located in BADJ region, and is expressing SPC and CC10 (arrowhead). (c) The sparse tdTomato+ cell (arrowhead) does not express AT1 cell marker T1a. Scale bars, 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 9 BASCs differentiate mainly into club cells after naphthalene-induced lung injury.
Immunostaining for CC10, tdTomato and SPC on lung tissue sections after naphthalene treatment. Yellow arrowheads indicate tdTomato+CC10+SPC+ BASCs at BADJ; white arrowheads indicate tdTomato+CC10+SPC– club cells. Scale bars, 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 10 BASCs contribute to club cells and ciliated cells after bronchiolar injury.
(a) Schematic figure showing timeline for tamoxifen (Tam), Naphthalene (Naph.) treatment, and lung tissue analysis. Mice were treated with naphthalene at 3 weeks after tamoxifen induction. (b) Diagram showing BSACs regenerate terminal bronchiole after Naph. injury. (c) Immunostaining for CC10, SPC and tdTomato on lung section shows a subset of tdTomato+ cells residing in BADJ continue to express CC10 and SPC (arrowheads), while the majority of tdTomato+ cells are CC10+SPC– and detected in the terminal bronchiole. (d) Immunostaining for tdTomato, CC10 and β-Tubulin or Acetylated-Tubulin on lung section shows tdTomato+ ciliated cells (arrowheads). (e) Immunostaining for CGRP, tdTomato and CC10 on tissue sections shows tdTomato+ cells (arrowheads) do not express CGRP. (f) Immunostaining for T1a and tdTomato on lung section shows tdTomato+ cells (arrowheads) do not express T1a. Scale bars, 100 µm. Each image is a representative of five individual samples.
Supplementary Figure 11 Expansion of BASC-derived AT1 and AT2 cells after bleomycin-induced lung injury.
(a) Schematic figure showing experimental design. EdU was injected at 24 hours before analysis. (b) Immunostaining for EdU on lung sections after bleomycin or Vehicle treatment. (c) Quantification of the percentage of EdU+ cells in bleomycin (Bleom.) or vehicle-treated lung tissues. P is calculated by two-tailed t-test; n = 5 biologically independent mice; data are showing by box and whiskers plot; box spans the interquartile range with band inside the box represents median and whiskers represent maximum and minimum values. (d-f) Immunostaining for tdTomato, EdU and SPC, CC10 or T1a on sections. Dotted line demarcates bronchioles. Boxed regions are magnified on the right. Arrowheads indicate EdU+tdTomato+ cells. YZ indicate signals from dotted lines on Z-stack images in f (Vehicle). Scale bars, 100 µm. (g) Immunostaining for CC10, tdTomato and SPC on lung tissue sections shows that the majority of tdTomato+ cells are SPC+CC10– close to the BADJs. Each image is a representative of 5 individual samples.
Supplementary Figure 12 BASCs do not contribute to fibroblasts, pericytes, smooth muscle cells or endothelial cells in bleomycin-treated lung.
(a) Sirius Red staining on bleomycin or vehicle treated lung sections. (b-g) Immunostaining for tdTomato and different cell lineage markers PDGFRa, PECAM, PDGFRb, T1a, aSMA and VE-cad. XZ and YZ indicates signals from dotted lines on Z-stack images. Boxed regions are magnified on the right. Scale bars, 100 µm. Each image is representative of 5 individual samples.
Supplementary Figure 13 BASCs contribute to AT1 and AT2 cells after alveolar injury.
(a) Schematic figure showing timeline for tamoxifen (Tam), bleomycin treatment, and lung tissue analysis. Bleomycin is treated at 3 weeks after tamoxifen induction. (b) Diagram showing BASCs regenerate alveoli after bleomycin injury. (c) Immunostaining for T1a and tdTomato on lung section shows a subset of tdTomato+ cells residing in BADJ region express AT1 (arrowheads). XZ and YZ indicate signals from dotted lines on Z-stack images. (d) Immunostaining for SPC and tdTomato on lung section shows tdTomato+SPC+ AT2 cells (arrowheads). (e) Immunostaining for CC10, SPC and tdTomato on tissue sections shows most tdTomato+ cells (arrowheads) differentiate into SPC+ AT2 cells and do not express CC10. Scale bars, 100 µm. Each image is a representative of five individual samples.
Supplementary Figure 14 Clonal analysis of BASCs after lung injuries.
(a) Immunostaining for GFP, YFP, RFP, Acetylated-tubulin on Sftpc-DreER;Scgb1a1-CreER;R26-Confetti2 mouse lung sections after naphthalene treatment. Arrowheads indicate YFP+Acetylated-tubulin+ cell or RFP+Acetylated-tubulin+ cells in naphthalene-induced lung. (b-e) Immunostaining for GFP, YFP, RFP, CC10, SPC or T1a on Sftpc-DreER;Scgb1a1-CreER;R26-Confetti2 mouse lung sections after naphthalene, vehicle or bleomycin treatment. Arrowheads indicates GFP/YFP+SPC+ cell in naphthalene-induced lung (b), GFP+SPC+ cell in vehicle-induced lung (c), RFP+CC10+ cell in bleomycin-induced lung (d) and RFP+T1a– cell in vehicle-treated lung (e). Scale bars, 100 µm. Each image is a representative of 4 individual samples.
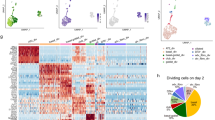
Supplementary Figure 15 Comparison of cell clusters from scRNA-seq.
(a) t-SNE of 480 scRNA-seq profiles (points), colored by expression of selected AT2, Club cell and Ciliated cell markers. (b) Pearson correlation coefficients (r) between 4 cell clusters. (c,d) Distribution of expression levels of Plin2 and Lrg1 in each cell cluster, Violin plots show the Gaussian kernel probability densities of the data. (e-h) t-SNE plot of 455 scRNA-seq profiles (points) including AT2, BASCs-1, BASCs-2 and Club cell clusters, showing genes enriched in BASCs-1 (e,f) and BASCs-2 (g,h) subpopulation.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–15
Rights and permissions
About this article

Cite this article
Liu, Q., Liu, K., Cui, G. et al. Lung regeneration by multipotent stem cells residing at the bronchioalveolar-duct junction. Nat Genet 51, 728–738 (2019). https://doi.org/10.1038/s41588-019-0346-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-019-0346-6
This article is cited by
-
The novel molecular mechanism of pulmonary fibrosis: insight into lipid metabolism from reanalysis of single-cell RNA-seq databases
Lipids in Health and Disease (2024)
-
Airway epithelial cell identity and plasticity are constrained by Sox2 during lung homeostasis, tissue regeneration, and in human disease
npj Regenerative Medicine (2024)
-
Single-cell division tracing and transcriptomics reveal cell types and differentiation paths in the regenerating lung
Nature Communications (2024)
-
Intraluminal chloride regulates lung branching morphogenesis: involvement of PIEZO1/PIEZO2
Respiratory Research (2023)
-
Lung development and regeneration: newly defined cell types and progenitor status
Cell Regeneration (2023)
