
Abstract
Steroid hormone receptors are ligand-binding transcription factors essential for mammalian physiology. The androgen receptor (AR) binds androgens mediating gene expression for sexual, somatic and behavioural functions, and is involved in various conditions including androgen insensitivity syndrome and prostate cancer1. Here we identified functional mutations in the formin and actin nucleator DAAM2 in patients with androgen insensitivity syndrome. DAAM2 was enriched in the nucleus, where its localization correlated with that of the AR to form actin-dependent transcriptional droplets in response to dihydrotestosterone. DAAM2 AR droplets ranged from 0.02 to 0.06 µm3 in size and associated with active RNA polymerase II. DAAM2 polymerized actin directly at the AR to promote droplet coalescence in a highly dynamic manner, and nuclear actin polymerization is required for prostate-specific antigen expression in cancer cells. Our data uncover signal-regulated nuclear actin assembly at a steroid hormone receptor necessary for transcription.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
All data are available in the main text or supplementary materials. Source data are provided with this paper.
References
Matsumoto, T. et al. The androgen receptor in health and disease. Annu. Rev. Physiol. 75, 201–224 (2013).
Hornig, N. C. & Holterhus, P. M. Molecular basis of androgen insensitivity syndromes. Mol. Cell. Endocrinol. 523, 111146 (2021).
Hornig, N. C. et al. Identification of an AR mutation-negative class of androgen insensitivity by determining twenty-four hours endogenous AR activity. J. Clin. Endocrinol. Metab. 101, 4468–4477 (2016).
Faix, J. & Grosse, R. Staying in shape with formins. Dev. Cell 10, 693–706 (2006).
Goode, B. L. & Eck, M. J. Mechanism and function of formins in the control of actin assembly. Annu. Rev. Biochem. 76, 593–627 (2007).
Schneider, R. et al. DAAM2 variants cause nephrotic syndrome via actin dysregulation. Am. J. Hum. Genet. 107, 1113–1128 (2020).
Nakaya, M. A. et al. Identification and comparative expression analyses of Daam genes in mouse and Xenopus. Gene Expr. Patterns 5, 97–105 (2004).
Calvel, P. et al. A case of Wiedemann-Steiner syndrome associated with a 46,XY disorder of sexual development and gonadal dysgenesis. Sex. Dev. 9, 289–295 (2015).
Baarlink, C., Wang, H. & Grosse, R. Nuclear actin network assembly by formins regulates the SRF coactivator MAL. Science 340, 864–867 (2013).
Wang, Y. et al. GPCR-induced calcium transients trigger nuclear actin assembly for chromatin dynamics. Nat. Commun. 10, 5271 (2019).
Belin, B. J., Lee, T. & Mullins, R. D. DNA damage induces nuclear actin filament assembly by formin-2 and spire-1/2 that promotes efficient DNA repair. eLife 4, e07735 (2015).
Liu, C., Zhu, R. & Mao, Y. Nuclear actin polymerized by mDia2 confines centromere movement during CENP-A loading. iScience 9, 314–327 (2018).
Aymard, F. et al. Genome-wide mapping of long-range contacts unveils clustering of DNA double-strand breaks at damaged active genes. Nat. Struct. Mol. Biol. 24, 353–361 (2017).
Betzig, E. Excitation strategies for optical lattice microscopy. Opt. Express 13, 3021–3036 (2005).
Heintzmann, R. Saturated patterned excitation microscopy with two-dimensional excitation patterns. Micron 34, 283–291 (2003).
Gustafsson, M. G. Surpassing the lateral resolution limit by a factor of two using structured illumination microscopy. J. Microsc. 198, 82–87 (2000).
Rippe, K. & Papantonis, A. Functional organization of RNA polymerase II in nuclear subcompartments. Curr. Opin. Cell Biol. 74, 88–96 (2022).
Boehning, M. et al. RNA polymerase II clustering through carboxy-terminal domain phase separation. Nat. Struct. Mol. Biol. 25, 833–840 (2018).
Costes, S. V. et al. Automatic and quantitative measurement of protein-protein colocalization in live cells. Biophys. J. 86, 3993–4003 (2004).
Baarlink, C. et al. A transient pool of nuclear F-actin at mitotic exit controls chromatin organization. Nat. Cell Biol. 19, 1389–1399 (2017).
Plessner, M., Melak, M., Chinchilla, P., Baarlink, C. & Grosse, R. Nuclear F-actin formation and reorganization upon cell spreading. J. Biol. Chem. 290, 11209–11216 (2015).
Caridi, C. P. et al. Nuclear F-actin and myosins drive relocalization of heterochromatic breaks. Nature 559, 54–60 (2018).
Schrank, B. R. et al. Nuclear ARP2/3 drives DNA break clustering for homology-directed repair. Nature 559, 61–66 (2018).
Cho, W. K. et al. Mediator and RNA polymerase II clusters associate in transcription-dependent condensates. Science 361, 412–415 (2018).
Funk, J. et al. Profilin and formin constitute a pacemaker system for robust actin filament growth. eLife 8, e50963 (2019).
Waizumi, T. et al. Polymerization/depolymerization of actin cooperates with the morphology and stability of cell-sized droplets generated in a polymer solution under a depletion effect. J. Chem. Phys. 155, 075101 (2021).
Plessner, M., Knerr, J. & Grosse, R. Centrosomal actin assembly is required for proper mitotic spindle formation and chromosome congression. iScience 15, 274–281 (2019).
Tsopoulidis, N. et al. T cell receptor–triggered nuclear actin network formation drives CD4+ T cell effector functions. Sci. Immunol. 4, eaav1987 (2019).
Li, F. & Higgs, H. N. The mouse Formin mDia1 is a potent actin nucleation factor regulated by autoinhibition. Curr. Biol. 13, 1335–1340 (2003).
Grotsch, H. et al. RWDD1 interacts with the ligand binding domain of the androgen receptor and acts as a coactivator of androgen-dependent transactivation. Mol. Cell. Endocrinol. 358, 53–62 (2012).
Kircher, M. et al. A general framework for estimating the relative pathogenicity of human genetic variants Nat. Genet. 46, 310–315 (2014).
Eggers, S. et al. Disorders of sex development: insights from targeted gene sequencing of a large international patient cohort. Genome Biol. 17, 243 (2016).
Audi, L. et al. GENETICS IN ENDOCRINOLOGY: approaches to molecular genetic diagnosis in the management of differences/disorders of sex development (DSD): position paper of EU COST Action BM 1303 ‘DSDnet’. Eur. J. Endocrinol. 179, R197–R206 (2018).
Ullah, M. Z., Aono, M. & Seddiqui, M. H. Estimating a ranked list of human hereditary diseases for clinical phenotypes by using weighted bipartite network. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2013, 3475–3478 (2013).
Acknowledgements
We thank B. Karwelies and P. Davarnia for technical support; laboratory members of the Grosse group for discussions; O. Wunderlich for technical assistance; J. Altmueller for help with the sequencing facility; and H. U. Schweikert for providing GSFs. The study has been funded by the German Research Council (Deutsche Forschungsgemeinschaft) to N.C.H. (nos. HO 6028/2-1 and HO 6028/3-1) and R.G. (no. GR2111/13-1) and under Germany’s Excellence Strategy (EXC-2189, project ID: 390939984).
Author information
Authors and Affiliations
Contributions
J.K. performed most experiments. N.C.H. performed experiments for Fig. 1a–c and Extended Data Fig. 1 and IF staining for Figs. 2 and 3. R.W. and H.G. performed experiments for Fig. 1d. C.S. performed experiments for Extended Data Fig. 2b,c. H.W. and P.G. performed experiments for Fig. 1e–i. R.G. and N.C.H. acquired funding and wrote the original draft of the manuscript. R.G., C.S., J.K., N.C.H., R.W., A.C., M.S. and P.-M.H. contributed to writing and review editing. R.G., C.S. and J.K. performed data analysis for Figs. 2–4.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Dyche Mullins, Lothar Schermelleh and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Variant validation and protein expression of AR and DAAM2 in male control GSF and both mutant strains.
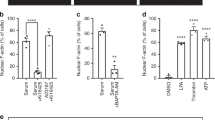
a Sanger Validation of the variants found in DAAM2 mutant strains GSF-104 (NM_001201427.1 c.1840_1841insC) and GSF-163 (NM_001201427.1 c.495C>G). b DAAM2 knock down efficiency in male control GSF-22 at the mRNA and c protein expression level. d DAAM2 mRNA expression levels in male control GSF and both mutant strains. e Western blots showing whole cell extract expressions of AR and DAAM2 in two different scrotal (GSF-22 and GSF-26) and two different foreskin controls (GSF-122 and GSF-128) together with the DAAM2 mutants (GSF-104 and GSF-163). Ponceau staining served as loading control. f Quantification of DAAM2 protein levels normalized to ponceau. g Quantification of AR protein levels normalized to ponceau. The experiments were performed in three independent biological replicates. Means +/− SD are shown. P-values were calculated using a two-sided t-test. * p = 0.022, ** p = 0.0034, **** p < 0001. Values of p < 0001 are denoted by four stars, values of p < 001 are denoted by three stars, values of p < 005 are denoted by two stars, and values of p < 05 are denoted by one star. h Cytoplasmic (CE) and nuclear (NE) expression of AR, DAAM2 in male control GSF (GSF-26 and GSF-122) and both mutant strains. TAF15 and TUB served as compartment specific loading controls.
Extended Data Fig. 2 Microscopy validation and quantification analysis.
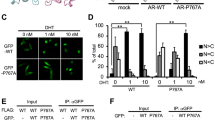
a 3D-volume rendering of DAAM2 IF signal for homozygous wt and heterozygous mutant cells, showing DAAM2 distribution throughout the cell. b Polystyrene beads (Bangs Laboratories, IN, USA) with 200 nm diameter labelled with Dragon Green (Exc: 480/Em: 520) or Flash Red (Exc: 660/Em: 690) were premixed (2:1 ratio) and subsequently mixed with viscous Mowiol resin for embedding. Z-stacks of beads were acquired with 0.091 µm z-distance in lattice-SIM mode similar to the acquisition of droplets in nuclei which are shown at the top of the figure. Droplets and beads were analyzed for colocalizing volume (CoLo). AR-/DAAM2 droplet pairs are closer than the Rayleigh criterion. In contrast, no beads were observed that missed the Rayleigh criterion. An additional comparison of lattice-SIM illumination and laser-epi illumination is shown in the bottom panel. c For comparing the association of beads and droplets a colocalization analysis was performed similar to Extended Data Fig. 2d. 15 bead pairs with differential fluorescence and maximal proximity in the lateral- or z-plane were selected for analysis and compared to representative 15 droplet pairs in the same size of DHT treated cells from Fig. 2f, j; Extended Data Fig. 2d. Droplets have an increased colocalizing volume indicating closer association. d Quantification of colocalizing volume between DAAM2 and AR of detected colocalization events in Fig. 2j. e Quantification showing percentage of DAAM2 droplets colocalizing with AR droplets in comparison to total nuclear DAAM2 droplets. f Immunofluorescence of endogenous AR (magenta) and RNA Polymerase-II (Ser2P, cyan) in control scrotal cells, treated with EtOH or DHT as well as addition of Hexanediol or Swinholide to DHT-treated cells. Area of zooms are indicated above (upper panel); lower panels displays magnified areas. Colocalization between AR and RNA Pol-II Ser2P is indicated by white asterisks. Scale bar overview: 3 µm; zoom: 0.3 µm. All images are shown as maximum intensity projections.
Extended Data Fig. 3 DAAM2 is required for nuclear actin assembly at the AR.
a ELYRA7 SIM-burst mode timelapse video of NIH3T3 cell, stably expressing the nuclear actin chromobody-mCherry transfected with AR-GFP (magenta) displaying nuclear actin tails at the AR after DHT-induced signaling for 16 h; areas of zoom are indicated. b Images of NIH3T3 cells stably expressing nAC-mcherry (green) and transfected with AR-GFP (magenta). Cells were treated with si-ctrl or siDAAM2 for 72 h and imaged after DHT addition for 16 h showing inhibition of actin-positive AR droplet formation; area of zooms are indicated; zoom 2 is indicated in Fig. 4d. c Western blot showing DAAM2 knockdown efficiency after 72 h. d SIM-burst mode timelapse video of LNCaP cells transfected with AR-GFP (magenta), DAAM2-mScarlet (orange) and nAC-SNAP (green) and stimulated with DHT for 16 h, showing DAAM2 mediated nuclear actin assembly at bigger AR droplet indicated by white asterisks. e Western blot of PSA reporter gene lysates in LNCaP cells showing expression of transfected NLS-BFP-R62D-myc and NLS-BFP-myc using anti myc antibody. Histone H3 serves as loading control. f Western blot of PSA reporter gene lysates in LNCaP cells showing expression of transfected NLS-Arpin, NLS-d.n.Arp2 and Exportin-6 using anti mCherry, anti RFP antibodies. GAPDH serves as loading control. g,h Representative Western blot displaying knockdown efficiency of DAAM2 and DIAPH3 in LNCaP cells 48 h after siRNA transfection using anti DAAM2 and anti DIAPH3 antibodies. Tubulin serves as loading control. i Representative Western blot showing test-expression of si-RNA resistant wt and mutant DAAM2-mScarlet constructs used for PSA gene reporter assay, detected with anti RFP antibody.
Supplementary information
Supplementary Fig. 1
Uncropped and labelled scans of SDS–PAGE gels and immunoblots for all figures (.tiff). Cropped regions used in figures are indicated by boxes.
Supplementary Video 1
DHT signalling drives colocalization of nuclear actin at the AR. Overview of transfected NIH3T3 cell showing colocalizing AR (magenta) and nuclear actin (green) 16 h after DHT signalling. Time is shown as m:ss.ms.
Supplementary Video 2
DHT signalling drives dynamic nuclear actin assembly at the AR. Magnification of Supplementary Video 1 showing dynamic nuclear actin (green) polymerization at the AR (magenta) 16 h after DHT signalling. Time is shown as m:ss.ms.
Supplementary Video 3
DHT-dependent coalescence of the AR is associated with dynamic nuclear actin assembly. Magnification of Supplementary Video 1 showing nuclear actin-positive (green) AR droplets (magenta) undergoing coalescence over time. Time is shown as m:ss.ms.
Supplementary Video 4
DHT signalling drives the formation of nuclear actin-positive AR droplets. Overview of transfected NIH3T3 cell showing nuclear actin (green)-positive AR droplets (magenta) 16 h after DHT signalling. Time is shown as m:ss.ms.
Supplementary Video 5
5 DHT signalling drives the formation of nuclear actin-positive AR droplets. Magnification of transfected NIH3T3 cell showing nuclear actin (green)-positive AR droplets (magenta) 16 h after DHT signalling. Time is shown as m:ss.ms.
Supplementary Video 6
DHT signalling drives droplet formation consisting of the AR, DAAM2 and nuclear actin. Overview of triple-transfected LNCaP cell showing colocalizing AR (magenta), nuclear actin (green) and DAAM2 (orange) 16 h after DHT signalling. Time is shown as m:ss.ms.
Supplementary Video 7
DHT signalling drives dynamic nuclear actin polymerization by DAAM2 at the AR. Magnification from overview Supplementaty Video 6. Triple-transfected LNCaP cell showing dynamic actin (green) polymerization by DAAM2 (orange) at the AR (magenta) 16 h after DHT signalling. Video shows first triple merge and subsequent respective double channels for the same time interval, indicated by labelling in the video. Time is shown as m:ss.ms.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Knerr, J., Werner, R., Schwan, C. et al. Formin-mediated nuclear actin at androgen receptors promotes transcription. Nature 617, 616–622 (2023). https://doi.org/10.1038/s41586-023-05981-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-023-05981-1
This article is cited by
-
MolPhase, an advanced prediction algorithm for protein phase separation
The EMBO Journal (2024)
-
DNA replication and replication stress response in the context of nuclear architecture
Chromosoma (2024)
-
Nuclear actin polymerization rapidly mediates replication fork remodeling upon stress by limiting PrimPol activity
Nature Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
