
Abstract
Metabolic rewiring underlies the effector functions of macrophages1,2,3, but the mechanisms involved remain incompletely defined. Here, using unbiased metabolomics and stable isotope-assisted tracing, we show that an inflammatory aspartate–argininosuccinate shunt is induced following lipopolysaccharide stimulation. The shunt, supported by increased argininosuccinate synthase (ASS1) expression, also leads to increased cytosolic fumarate levels and fumarate-mediated protein succination. Pharmacological inhibition and genetic ablation of the tricarboxylic acid cycle enzyme fumarate hydratase (FH) further increases intracellular fumarate levels. Mitochondrial respiration is also suppressed and mitochondrial membrane potential increased. RNA sequencing and proteomics analyses demonstrate that there are strong inflammatory effects resulting from FH inhibition. Notably, acute FH inhibition suppresses interleukin-10 expression, which leads to increased tumour necrosis factor secretion, an effect recapitulated by fumarate esters. Moreover, FH inhibition, but not fumarate esters, increases interferon-β production through mechanisms that are driven by mitochondrial RNA (mtRNA) release and activation of the RNA sensors TLR7, RIG-I and MDA5. This effect is recapitulated endogenously when FH is suppressed following prolonged lipopolysaccharide stimulation. Furthermore, cells from patients with systemic lupus erythematosus also exhibit FH suppression, which indicates a potential pathogenic role for this process in human disease. We therefore identify a protective role for FH in maintaining appropriate macrophage cytokine and interferon responses.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Data availability
Proteomics data from Fig. 1d were previously deposited11 to the ProteomeXchange Consortium through the PRIDE partner repository with the dataset identifier PXD029155. All other proteomics, RNA-seq data and metabolomics data have been deposited to Dryad (https://doi.org/10.5061/dryad.6wwpzgn28). All other data are available from the corresponding authors upon request. Source data are provided with this paper.
References
Mills, E. L. et al. Succinate dehydrogenase supports metabolic repurposing of mitochondria to drive inflammatory macrophages. Cell 167, 457–470.e13 (2016).
Mills, E. L. et al. Itaconate is an anti-inflammatory metabolite that activates Nrf2 via alkylation of KEAP1. Nature 556, 113–117 (2018).
Tannahill, G. M. et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 496, 238–242 (2013).
Lampropoulou, V. et al. Itaconate links inhibition of succinate dehydrogenase with macrophage metabolic remodeling and regulation of inflammation. Cell Metab. 24, 158–166 (2016).
Jha, A. K. et al. Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization. Immunity 42, 419–430 (2015).
Billingham, L. K. et al. Mitochondrial electron transport chain is necessary for NLRP3 inflammasome activation. Nat. Immunol. 23, 692–704 (2022).
Mills, E. L., Kelly, B. & O’Neill, L. A. J. Mitochondria are the powerhouses of immunity. Nat. Immunol. 18, 488–498 (2017).
Adam, J. et al. Renal cyst formation in Fh1-deficient mice is independent of the Hif/Phd pathway: roles for fumarate in KEAP1 succination and Nrf2 signaling. Cancer Cell 20, 524–537 (2011).
Kornberg, M. D. et al. Dimethyl fumarate targets GAPDH and aerobic glycolysis to modulate immunity. Science 360, 449–453 (2018).
Humphries, F. et al. Succination inactivates gasdermin D and blocks pyroptosis. Science 369, 1633–1637 (2020).
Williams, N. C. et al. Signaling metabolite L-2-hydroxyglutarate activates the transcription factor HIF-1α in lipopolysaccharide-activated macrophages. J. Biol. Chem. 298, 101501 (2021).
Cordes, T. et al. Immunoresponsive gene 1 and itaconate inhibit succinate dehydrogenase to modulate intracellular succinate levels. J. Biol. Chem. 291, 14274–14284 (2016).
Sass, E., Blachinsky, E., Karniely, S. & Pines, O. Mitochondrial and cytosolic isoforms of yeast fumarase are derivatives of a single translation product and have identical amino termini. J. Biol. Chem. 276, 46111–46117 (2001).
Adam, J. et al. A role for cytosolic fumarate hydratase in urea cycle metabolism and renal neoplasia. Cell Rep. 3, 1440–1448 (2013).
Takeuchi, T., Schumacker, P. T. & Kozmin, S. A. Identification of fumarate hydratase inhibitors with nutrient-dependent cytotoxicity. J. Am. Chem. Soc. 137, 564–567 (2015).
Ryan, D. G. et al. Disruption of the TCA cycle reveals an ATF4-dependent integration of redox and amino acid metabolism. eLife 10, e72593 (2021).
Hayashi, G. et al. Dimethyl fumarate mediates Nrf2-dependent mitochondrial biogenesis in mice and humans. Hum. Mol. Genet. 26, 2864–2873 (2017).
Sciacovelli, M. et al. Fumarate is an epigenetic modifier that elicits epithelial-to-mesenchymal transition. Nature 537, 544–547 (2016).
Wang, Y. P. et al. Malic enzyme 2 connects the Krebs cycle intermediate fumarate to mitochondrial biogenesis. Cell Metab. 33, 1027–1041 e1028 (2021).
Liao, S. T. et al. 4-Octyl itaconate inhibits aerobic glycolysis by targeting GAPDH to exert anti-inflammatory effects. Nat. Commun. 10, 5091 (2019).
Crooks, D. R. et al. Mitochondrial DNA alterations underlie an irreversible shift to aerobic glycolysis in fumarate hydratase-deficient renal cancer. Sci. Signal. 14, eabc4436 (2021).
Blatnik, M., Frizzell, N., Thorpe, S. R. & Baynes, J. W. Inactivation of glyceraldehyde-3-phosphate dehydrogenase by fumarate in diabetes: formation of S-(2-succinyl)cysteine, a novel chemical modification of protein and possible biomarker of mitochondrial stress. Diabetes 57, 41–49 (2008).
Tyrakis, P. A. et al. Fumarate hydratase loss causes combined respiratory chain defects. Cell Rep. 21, 1036–1047 (2017).
Ternette, N. et al. Inhibition of mitochondrial aconitase by succination in fumarate hydratase deficiency. Cell Rep. 3, 689–700 (2013).
Sullivan, L. B. et al. The proto-oncometabolite fumarate binds glutathione to amplify ROS-dependent signaling. Mol. Cell 51, 236–248 (2013).
Zheng, L. et al. Fumarate induces redox-dependent senescence by modifying glutathione metabolism. Nat. Commun. 6, 6001 (2015).
Bambouskova, M. et al. Electrophilic properties of itaconate and derivatives regulate the IκBζ–ATF3 inflammatory axis. Nature 556, 501–504 (2018).
Raimundo, N., Vanharanta, S., Aaltonen, L. A., Hovatta, I. & Suomalainen, A. Downregulation of SRF–FOS–JUNB pathway in fumarate hydratase deficiency and in uterine leiomyomas. Oncogene 28, 1261–1273 (2009).
Hu, X. et al. IFN-γ suppresses IL-10 production and synergizes with TLR2 by regulating GSK3 and CREB/AP-1 proteins. Immunity 24, 563–574 (2006).
Angel, P., Hattori, K., Smeal, T. & Karin, M. The jun proto-oncogene is positively autoregulated by its product, Jun/AP-1. Cell 55, 875–885 (1988).
Dickinson, S. E. et al. Inhibition of activator protein-1 by sulforaphane involves interaction with cysteine in the cFos DNA-binding domain: implications for chemoprevention of UVB-induced skin cancer. Cancer Res. 69, 7103–7110 (2009).
de Waal Malefyt, R., Abrams, J., Bennett, B., Figdor, C. G. & de Vries, J. E. Interleukin 10 (IL-10) inhibits cytokine synthesis by human monocytes: an autoregulatory role of IL-10 produced by monocytes. J. Exp. Med. 174, 1209–1220 (1991).
Luan, H. H. et al. GDF15 is an inflammation-induced central mediator of tissue tolerance. Cell 178, 1231–1244.e11 (2019).
Day, E. A. et al. Metformin-induced increases in GDF15 are important for suppressing appetite and promoting weight loss. Nat. Metab. 1, 1202–1208 (2019).
Coll, A. P. et al. GDF15 mediates the effects of metformin on body weight and energy balance. Nature 578, 444–448 (2020).
Wang, Y. et al. SLC25A39 is necessary for mitochondrial glutathione import in mammalian cells. Nature 599, 136–140 (2021).
Weng, J. H. et al. Colchicine acts selectively in the liver to induce hepatokines that inhibit myeloid cell activation. Nat. Metab. 3, 513–522 (2021).
Eisenstein, A. et al. Activation of the transcription factor NRF2 mediates the anti-inflammatory properties of a subset of over-the-counter and prescription NSAIDs. Immunity 55, 1082–1095.e5 (2022).
Asadullah, K. et al. Influence of monomethylfumarate on monocytic cytokine formation—explanation for adverse and therapeutic effects in psoriasis? Arch. Dermatol. Res. 289, 623–630 (1997).
Arts, R. J. et al. Glutaminolysis and fumarate accumulation integrate immunometabolic and epigenetic programs in trained immunity. Cell Metab. 24, 807–819 (2016).
Ryan, D. G. et al. Nrf2 activation reprograms macrophage intermediary metabolism and suppresses the type I interferon response. iScience 25, 103827 (2022).
Shanmugasundaram, K. et al. The oncometabolite fumarate promotes pseudohypoxia through noncanonical activation of NF-κB signaling. J. Biol. Chem. 289, 24691–24699 (2014).
West, A. P. et al. Mitochondrial DNA stress primes the antiviral innate immune response. Nature 520, 553–557 (2015).
Sliter, D. A. et al. Parkin and PINK1 mitigate STING-induced inflammation. Nature 561, 258–262 (2018).
McArthur, K. et al. BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science 359, eaao6047 (2018).
Dang, E. V., McDonald, J. G., Russell, D. W. & Cyster, J. G. Oxysterol restraint of cholesterol synthesis prevents AIM2 inflammasome activation. Cell 171, 1057–1071.e11 (2017).
Haag, S. M. et al. Targeting STING with covalent small-molecule inhibitors. Nature 559, 269–273 (2018).
Stunz, L. L. et al. Inhibitory oligonucleotides specifically block effects of stimulatory CpG oligonucleotides in B cells. Eur. J. Immunol. 32, 1212–1222 (2002).
Prantner, D. et al. 5,6-Dimethylxanthenone-4-acetic acid (DMXAA) activates stimulator of interferon gene (STING)-dependent innate immune pathways and is regulated by mitochondrial membrane potential. J. Biol. Chem. 287, 39776–39788 (2012).
Dhir, A. et al. Mitochondrial double-stranded RNA triggers antiviral signalling in humans. Nature 560, 238–242 (2018).
Tigano, M., Vargas, D. C., Tremblay-Belzile, S., Fu, Y. & Sfeir, A. Nuclear sensing of breaks in mitochondrial DNA enhances immune surveillance. Nature 591, 477–481 (2021).
Kariko, K., Buckstein, M., Ni, H. & Weissman, D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity 23, 165–175 (2005).
Rai, P. et al. IRGM1 links mitochondrial quality control to autoimmunity. Nat. Immunol. 22, 312–321 (2021).
Kruger, A. et al. Human TLR8 senses UR/URR motifs in bacterial and mitochondrial RNA. EMBO Rep. 16, 1656–1663 (2015).
Pichlmair, A. et al. RIG-I-mediated antiviral responses to single-stranded RNA bearing 5′-phosphates. Science 314, 997–1001 (2006).
Koshiba, T., Yasukawa, K., Yanagi, Y. & Kawabata, S. Mitochondrial membrane potential is required for MAVS-mediated antiviral signaling. Sci. Signal. 4, ra7 (2011).
Kim, S. et al. Mitochondrial double-stranded RNAs govern the stress response in chondrocytes to promote osteoarthritis development. Cell Rep. 40, 111178 (2022).
Rasa, S. M. M. et al. Inflammaging is driven by upregulation of innate immune receptors and systemic interferon signaling and is ameliorated by dietary restriction. Cell Rep. 39, 111017 (2022).
Buskiewicz, I. A. et al. Reactive oxygen species induce virus-independent MAVS oligomerization in systemic lupus erythematosus. Sci. Signal. 9, ra115 (2016).
Ruiz-Limon, P. et al. Atherosclerosis and cardiovascular disease in systemic lupus erythematosus: effects of in vivo statin treatment. Ann. Rheum. Dis. 74, 1450–1458 (2015).
Davis, P., Cunnington, P. & Hughes, G. R. Double-stranded RNA antibodies in systemic lupus erythematosus. Ann. Rheum. Dis. 34, 239–243 (1975).
Caielli, S. et al. Oxidized mitochondrial nucleoids released by neutrophils drive type I interferon production in human lupus. J. Exp. Med. 213, 697–713 (2016).
Sarkar, P. et al. Reduced expression of mitochondrial fumarate hydratase in progressive multiple sclerosis contributes to impaired in vitro mesenchymal stromal cell-mediated neuroprotection. Mult. Scler. 28, 1179–1188 (2022).
Zecchini, V. et al. Fumarate induces vesicular release of mtDNA to drive innate immunity. Nature https://doi.org/10.1038/s41586-023-05770-w (2023).
Li, Q. et al. RNA editing underlies genetic risk of common inflammatory diseases. Nature 608, 569–577 (2022).
Pang, Z. et al. MetaboAnalyst 5.0: narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 49, W388–W396 (2021).
Anders, S. & Huber, W. Differential expression analysis for sequence count data. Genome Biol. 11, R106 (2010).
Shah, A. D., Goode, R. J. A., Huang, C., Powell, D. R. & Schittenhelm, R. B. LFQ-Analyst: an easy-to-use interactive web platform to analyze and visualize label-free proteomics data preprocessed with MaxQuant. J. Proteome Res. 19, 204–211 (2020).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–W97 (2016).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Acknowledgements
We would like to thank members of the O’Neill Lab for discussions and staff at Novogene for assistance with RNA-seq; B. Moran and G. McManus for assistance with flow cytometry and confocal microscopy, respectively; A. Dhir for discussions; and A. Capps for assistance with anti-2SC immunoblotting. L.A.J.O. was funded by the European Research Council (Metabinnate 834370) and the Science Foundation Ireland (20/SPP/3685). V.Z. was funded by the WWCR (14-0319). A.V.K. was funded by the Wellcome Trust (Multiuser Equipment Grant, 208402/Z/17/Z). N.F. was funded by the NIH (R01NS1268). C. Johansson was funded by the Medical Research Council UK (MR/V000659/1). C. Jefferies was funded by the NIH (R01AI164504), the Office of the Assistant Secretary of Defense for Health Affairs through the Department of Defense Lupus Research Program (LRP), (W81XWH-18-1-0709) and Cedars-Sinai Precision Health RFP 2020. M.P.M. was funded by the Medical Research Council UK (MC_UU_00028/4) and the Wellcome Trust (Investigator award 220257/Z/20/Z). C.F. was funded by the Medical Research Council UK (MRC_MC_UU_12022/6) and the European Research Council (Consolidator ERC819920). Schematics in Fig. 1i, Extended Data Figs. 4e and 6j and Supplementary Fig. 1 were created using BioRender (https://biorender.com).
Author information
Authors and Affiliations
Contributions
A.H., C.G.P., D.G.R. and L.A.J.O. conceptualized the project. A.H., C.G.P. and D.G.R. were lead experimentalists, provided intellectual input, designed all experiments, analysed and visualized the data and co-wrote the paper with input from all authors. E.A.D. performed in vivo experiments. E.N.M., L.H., G.D.L.S., M.I., D.J.W., S.V. and C. Jefferies generated data from patients with SLE. J.E.T.-K. assisted with immunofluorescence experiments. C.F., M. Yang, A.S.H.C. and E.N. assisted with metabolomics. A.B.-C. and A.V.K. assisted with proteomics. A.F.M., M. Yin, T.A.J.R., A.M.C. and H.A.P. performed in vitro experiments. C.F. and V.Z. provided inducible Fh1+/fl and Fh1fl/fl mouse tissue. N.F. verified protein succination with 2SC antibody on provided macrophage lysates. C. Johansson provided Mavs–/– mouse tissue. M.P.M. and C.F. provided intellectual input and oversaw a portion of the research programme. L.A.J.O. obtained funding and oversaw the research programme.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Navdeep Chandel and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 LPS stimulation drives fumarate accumulation and protein succination.
a-c, Fumarate-mediated protein succination with LPS (n = 3) and 2SC abundance in NS and LPS-stimulated BMDMs (n = 5; LPS 4 h). d, Heatmap of metabolites linked to aspartate-argininosuccinate shunt in NS and LPS-stimulated BMDMs (n = 5; LPS 24 h) e, Metabolite abundance of aspartate-argininosuccinate shunt metabolites in LPS-stimulated BMDMs pre-treated with DMSO or AOAA (n = 3; LPS 4 h; aspartate (P = 0.0000005)). f, Asl expression with silencing of Asl following LPS stimulation (n = 3; LPS 24 h). g, Fumarate levels with silencing of Asl following LPS stimulation (n = 3; LPS 24 h). c,e-g, Data are mean ± s.e.m. a, 1 representative blot of 3 shown. n = biological replicates. P values calculated using two-tailed Student’s t-test for paired comparisons or one-way ANOVA for multiple comparisons.
Extended Data Fig. 2 LPS stimulation drives fumarate accumulation via glutamine anaplerosis and an aspartate-argininosuccinate shunt.
a, Schematic diagram indicating U-13C-glutamine tracing into distinct metabolic modules. b, U-13C-glutamine tracing into glutamate, α-KG and succinate in LPS-treated BMDMs (m+4 and m+5 labelling intensity and total isotopologue fraction distribution) (n = 3; LPS 4 h). c, U-13C-glutamine tracing into γ-glutamylcysteine, GSH and GSSG in LPS-treated BMDMs (m+5 labelling intensity and total isotopologue fraction distribution) (n = 3; LPS 4 h). d, U-13C-glutamine tracing into aspartate, argininosuccinate, fumarate and malate in LPS-treated BMDMs (m+4 labelling intensity and total isotopologue fraction distribution) (n = 3; LPS 4 h). Data are mean ± s.e.m. n = biological replicates. P values calculated using two-tailed Student’s t-test for paired comparisons.
Extended Data Fig. 3 LPS stimulation drives fumarate accumulation via glutamine anaplerosis and an aspartate-argininosuccinate shunt.
a, Schematic diagram indicating 15N2-glutamine tracing into distinct metabolic modules. b, 15N2-glutamine tracing into glutamate and asparagine in LPS-treated BMDMs (m+1 and m+2 labelling intensity and total isotopologue fraction distribution) (n = 3; LPS 4 h). c, 15N2-glutamine tracing into GSH and GSSG in LPS-treated BMDMs (m+1 and m+2 labelling intensity and total isotopologue fraction distribution) (n = 3; LPS 4 h). d, 15N2-glutamine tracing into aspartate, arginine and citrulline in LPS-treated BMDMs (m+1 labelling intensity and total isotopologue fraction distribution) (n = 3; LPS 4 h; aspartate (P = 0.000001)). Data are mean ± s.e.m. n = biological replicates. P values calculated using one-way ANOVA for multiple comparisons.
Extended Data Fig. 4 Increase in aspartate-argininosuccinate shunt metabolites in cytosol and Irg1–/– macrophages.
Heatmap (min-max) of metabolites linked to mitochondrial bioenergetics and redox signalling (a) and the aspartate-argininosuccinate shunt (b) in NS and BMDMs (n = 3; LPS 24 h). c, Metabolite abundance of TCA cycle and aspartate-argininosuccinate shunt metabolites in WT and Irg1–/– BMDMs (n = 3; LPS 24 h); itaconate (P = 0.00000000000002, succinate (P = 0.00000003), fumarate (P = 0.000018)). d, Nitrite levels in WT and Irg1–/– BMDMs (n = 3; LPS 24 h). e, Schematic of metabolic changes occurring during mid-phase TCA cycle rewiring in WT and Irg1–/– BMDMs. Data are mean ± s.e.m. n = biological replicates. P values calculated using two-tailed Student’s t-test for paired comparisons or one-way ANOVA for multiple comparisons. Schematic in panel e was created using BioRender (https://biorender.com).
Extended Data Fig. 5 FH deletion increases bioenergetic stress, fumarate, and mitochondrial membrane potential.
a, Bioenergetic ratios in BMDMs treated with DMSO or FHIN1 (n = 3). b, Fumarate and 2SC levels in BMDMs treated with DMSO or FHIN1 (n = 3). qPCR (n = 5) (c) and western blot (n = 2) (d) analysis of Fh1 expression in Fh1+/+ and Fh1–/– BMDMs (EtOH/TAM 72 h; LPS 4 h; Fh1+/+ NS vs Fh1+/+LPS (P = 0.00000002), Fh1+/+ NS vs Fh1–/– NS (P = 0.00000000000002), Fh1–/– NS vs Fh1–/– LPS (P = 0.0000000000014)). e, Bioenergetic ratios in Fh1+/+ and Fh1–/– BMDMs (n = 3; EtOH/TAM 48 h). f, Heatmap of top 50 significantly abundant metabolites in Fh1+/+ and Fh1–/– BMDMs (n = 3; LPS 4 h). g, Fumarate and 2SC levels in Fh1+/+ and Fh1–/– BMDMs (n = 3; EtOH/TAM 72 h). h, Glycolysis as measured by ECAR in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 8 (DMSO/FHIN1); (n = 6 (DMF); LPS 4 h). n = technical replicates from 1 experiment performed with 3 pooled biological replicates. Data are mean ± s.d. i, Glyceraldehyde 3- phosphate (G3P) and 2,3-phosphoglycerate (2/3-PG) levels and ratio in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h; G3P (P = 0.00004)). Immunofluorescence (j) and quantification (k) of Mitotracker red staining in BMDMs pre-treated with DMSO or FHIN1 (n = 8 (DMSO); n = 19 (FHIN1); LPS 4 h). n = technical replicates from representative experiment. Scale bar = 20 μm. Data are mean ± s.d. a-c,e,g,i Data are mean ± s.e.m. Representative blots or images of 2 (d) or 1 experiment(s) (j) shown. n = biological replicates unless stated otherwise. P values calculated using two-tailed Student’s t-test for paired comparisons or one-way ANOVA for multiple comparisons.
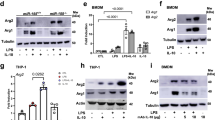
Extended Data Fig. 6 FH inhibition remodels inflammatory gene expression.
a, Il10 and Tnfa expression in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 5 (Il10); n = 6 (Tnfa); LPS 4 h; FHIN1/Il10 P = 0.000002, DMF/Il10 P = 0.0000004). b, Il1b expression and IL-6 release in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 6; 4 h LPS; DMF/Il1b (P = 0.000046), DMF/IL-6 (P = 0.00000002)). c, Enrichment map plot of shared significantly increased genes in BMDMs pre-treated with DMF or FHIN1 compared to DMSO control (n = 3; LPS 4 h). d, Western blot of total and phospho-AKT, JNK, ERK and p38 levels in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 2). e, Jun expression in RNA seq from BMDMs pre-treated with DMF or FHIN1 compared to DMSO control (n = 3; LPS 4 h). f, Fos expression in RNA seq from BMDMs pre-treated with DMF or FHIN1 compared to DMSO control (n = 3; LPS 4 h). g, Western blot of total and phospho-STAT3 levels in BMDMs pre-treated with anti-CD210 antibody (1 h) (n = 4; LPS 4 h). h, FH protein and gene expression levels in Fh1+/+ and Fh1+/– BMDMs (n = 2; EtOH/TAM 72 h). Data are mean. i, ELISA of IL-10 and TNF-α release in BMDMs pre-treated with DMSO or AOAA (n = 3; LPS 4 h; IL-10 (P = 0.000483)). j, Schematic depicting mild suppression of IL-10 expression during typical LPS signalling (left), and increased suppression of IL-10 following FH inhibition, leading to dysregulated TNF-α release (right). a,b,e,f,i Data are mean ± s.e.m. 1 representative blot of 2 (d, h) or 4 (g) shown. n = biological replicates. P values calculated using two-tailed Student’s t-test for paired comparisons or one-way ANOVA for multiple comparisons. Schematic in panel j was created using BioRender (https://biorender.com).
Extended Data Fig. 7 FH inhibition triggers the NRF2 and ATF4 stress response and promotes GDF15 release.
a, Heatmap of significantly differentially expressed RNA seq data in BMDMs pre-treated with FHIN1 compared to DMSO control (n = 3; LPS 4 h). Volcano plots of proteomics in BMDMs pre-treated with DMSO, FHIN1 (b) or DMF (c) (n = 5; LPS 4 h). d, ELISA of GDF15 in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). e, Nrf2 expression or ATF4 protein levels after silencing of Nrf2 or Atf4, respectively, in BMDMs pre-treated with DMSO or FHIN1 (n = 6; LPS 4 h). f, Gdf15 expression after silencing of Nrf2 or Atf4 respectively in BMDMs pre-treated with DMSO or FHIN1 (n = 3, LPS 4 h; FHIN1/Nrf2 RNAi (P = 0.000048)). d-f, Data are mean ± s.e.m. e, 1 representative blot of 6 shown. n = biological replicates unless stated otherwise. P values calculated using one-way ANOVA for multiple comparisons.
Extended Data Fig. 8 IFN-β release following FH inhibition is independent of cGAS-STING.
a, Heatmap (min-max) of significantly differentially expressed RNA seq data in BMDMs pre-treated with DMSO or DMF (n = 3; LPS 4 h). b, Phospho-STAT1, STAT1, phospho-JAK1 and JAK1 levels in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 3; LPS 4 h). c, Ifnb1 expression after silencing of Nrf2 in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 3, LPS 4 h). d, Nrf2 expression after silencing of Nrf2 in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 3, LPS 4 h; FHIN1 (P = 0.0000008), DMF (P = 0.0000012)). e, Ifnb1 expression in BMDMs pre-treated with DMSO or FHIN1 in the presence of NAC (n = 3; LPS 4 h). f, TRAF3 levels in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). g, IL-1β levels in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 3). h, p-p65 levels in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 3). i, D-loop and Non-NUMT DNA fold expression in ethidium bromide (EtBr)-treated BMDMs (n = 5; D-loop (P = 000000000031, Non-NUMT (P = 0.0000000012). j, Lamin B1 and α-tubulin in cytosolic and membrane-bound organelle fractions following digitonin fractionation (n = 3). k, IFN-β release from 2’,3’ cGAMP- or CpG-transfected BMDMs pre-treated (1 h) with C-178 or ODN2088 (n = 3 (cGAMP); n = 4(CpG); 3 h). l, Ifnb1 expression in BMDMs pre-treated with DMSO or FHIN1 in conjunction with C-178 or ODN2088 (1 h) respectively (n = 3; LPS 4 h). m, Cgas, Tmem173 and Tlr9 expression with silencing of Cgas, Tmem173 and Tlr9 respectively in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). n, IFN-β release with silencing of Cgas, Tmem173 and Tlr9 respectively from BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). o, Tmem173 expression in BMDMs pre-treated with DMSO, FHIN1 or DMF (n = 3, LPS 4 h). p, ND4, ND5 and ND6 RNA levels in whole cell extracts of BMDMs pre-treated with DMSO or FHIN1 in the presence of IMT1 (n = 5; LPS 4 h; ND5 (P = 0.000052)). q, ND4, ND5 and ND6 RNA levels in cytosolic extracts of BMDMs pre-treated with DMSO or FHIN1 in the presence or absence of IMT1 (n = 5; LPS 4 h). r, IFN-β release in BMDMs pre-treated with DMSO or FHIN1 in the presence of IMT1 (n = 3; LPS 4 h). c-e,i,k-r, Data are mean ± s.e.m. b,f-h,j, 1 representative blot of 3 shown. n = biological replicates. P values calculated using two-tailed Student’s t-test for paired comparisons or one-way ANOVA for multiple comparisons.
Extended Data Fig. 9 Mitochondrial membrane potential modifiers increase mtRNA and trigger IFN-β release.
a, Tlr7 expression with silencing of Tlr7 in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). b, Ddx58 and Ifih1 expression with silencing of Ddx58 and Ifih1 respectively in BMDMs pre-treated with DMSO or FHIN1 (n = 5; LPS 4 h; DMSO/Ddx58 (P = 0.000000000002), FHIN1/Ddx58 (P = 0.000000813792), DMSO/Ifih1 (P = 0.00000009), FHIN1/Ifih1 (P = 0.00000014)). c, Tlr3 expression and IFN-β release with silencing of Tlr3 in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h; DMSO/Tlr3 (P = 0.000000007), FHIN1/Tlr3 (P = 0.000013487)). d, TBK1 and p-TBK1 in BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). e, Ifnb1 expression in WT and Mavs–/– BMDMs pre-treated with DMSO or FHIN1 (n = 3; LPS 4 h). f, MFI of TMRM staining in BMDMs pre-treated with DMSO, FHIN1, oligomycin or valinomycin (n = 3, LPS 4 h). g, IFN-β release from BMDMs pre-treated with DMSO, FHIN1, oligomycin or valinomycin (n = 4; LPS 4 h; oligomycin (P = 0.0000003)). h, MFI of TMRM staining and IFN-β release from BMDMs pre-treated with DMSO or CCCP (n = 4 (TMRM), n = 3 (IFN-β); LPS 4 h; CCCP/IFN-β (P = 0.00000008)). i, MFI of TMRM staining in BMDMs pre-treated with DMSO or MMF (n = 3, LPS 4 h). Immunofluorescence (j) and quantification (k) of dsRNA in BMDMs pre-treated with DMSO, FHIN1 or oligomycin or transfected with poly (I:C) (n = 8; LPS 4 h). n = technical replicates from representative experiment. Data are mean ± s.d. Scale bar = 20 μm. l, D-loop fold expression in DNA and RNA isolated from cytosolic fractions of digitonin-fractionated BMDMs pre-treated with DMSO or oligomycin (n = 4 for mtDNA, n = 5 for mtRNA). Immunofluorescence (m) and quantification (n) of dsRNA in BMDMs pre-treated with DMSO or valinomycin (n = 9 (DMSO); n = 6 (Valinomycin); LPS 4 h). n = technical replicates from representative experiment. Data are mean ± s.d. Scale bar = 20 μm. o, Quantification of dsRNA immunofluorescence in Fh1+/+ and Fh1–/– BMDMs (n = 7 (Fh1+/+ Control); n = 6 (Fh1+/+ LPS); n = 12 (Fh1–/– Control); n = 10 (Fh1–/– LPS); EtOH/TAM 72 h; LPS 4 h). n = technical replicates from representative experiment. Data are mean ± s.d. a-c,e-i,l Data are mean ± s.e.m. d,j,m, 1 representative blot or image of 3 experiments shown. n = biological replicates unless stated otherwise. P values calculated using two-tailed Student’s t-test for paired comparisons, one-way ANOVA for multiple comparisons.
Extended Data Fig. 10 Prolonged LPS stimulation increases mitochondrial membrane potential and dsRNA.
a, MFI of TMRM staining in BMDMs (n = 3). Immunofluorescence (b) and quantification (c) of dsRNA in BMDMs (n = 8 (0/48 h); n = 9 (24 h)). n = technical replicates from representative experiment. Data are mean ± s.d. Scale bar = 20 μm. d, Ddx58 and Ifih1 expression in BMDMs (n = 4; LPS 4 h; Ddx58 (P = 0.0000000010), Ifih1 (P=0.00000012)). e, Fh1 expression in IFN-β-stimulated BMDMs (n = 3). a,d,e, Data are mean ± s.e.m. b, 1 representative image of 3 experiments shown. n = biological replicates unless stated otherwise. P values calculated using two-tailed Student’s t-test for paired comparisons, one-way ANOVA for multiple comparisons.
Supplementary information
Supplementary Figures
Supplementary Figs. 1 and 2 and uncropped blots.
Supplementary Table 1
A list of antibodies used in the study.
Supplementary Table 2
A list of primer sequences used in the study.
Source data
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hooftman, A., Peace, C.G., Ryan, D.G. et al. Macrophage fumarate hydratase restrains mtRNA-mediated interferon production. Nature 615, 490–498 (2023). https://doi.org/10.1038/s41586-023-05720-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-023-05720-6
This article is cited by
-
Metabolic regulation of tumor-associated macrophage heterogeneity: insights into the tumor microenvironment and immunotherapeutic opportunities
Biomarker Research (2024)
-
A break in mitochondrial endosymbiosis as a basis for inflammatory diseases
Nature (2024)
-
Blocking reverse electron transfer-mediated mitochondrial DNA oxidation rescues cells from PANoptosis
Acta Pharmacologica Sinica (2024)
-
Prognostic and immune predictive roles of a novel tricarboxylic acid cycle-based model in hepatocellular carcinoma
Scientific Reports (2024)
-
Mitochondria, Autophagy and Inflammation: Interconnected in Aging
Cell Biochemistry and Biophysics (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
