
Abstract
Bestrophin-2 (BEST2) is a member of the bestrophin family of calcium-activated anion channels that has a critical role in ocular physiology1,2,3,4. Here we uncover a directional permeability of BEST2 to glutamate that heavily favours glutamate exit, identify glutamine synthetase (GS) as a binding partner of BEST2 in the ciliary body of the eye, and solve the structure of the BEST2–GS complex. BEST2 reduces cytosolic GS activity by tethering GS to the cell membrane. GS extends the ion conducting pathway of BEST2 through its central cavity and inhibits BEST2 channel function in the absence of intracellular glutamate, but sensitizes BEST2 to intracellular glutamate, which promotes the opening of BEST2 and thus relieves the inhibitory effect of GS. We demonstrate the physiological role of BEST2 in conducting chloride and glutamate and the influence of GS in non-pigmented ciliary epithelial cells. Together, our results reveal a novel mechanism of glutamate release through BEST2–GS.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


References
Bakall, B. et al. Bestrophin-2 is involved in the generation of intraocular pressure. Invest. Ophthalmol. Vis. Sci. 49, 1563–1570 (2008).
Owji, A. P., Kittredge, A., Zhang, Y. & Yang, T. Structure and function of the Bestrophin family of calcium-activated chloride channels. Channels 15, 604–623 (2021).
Zhang, Y. et al. Enhanced inflow and outflow rates despite lower IOP in bestrophin-2-deficient mice. Invest. Ophthalmol. Vis. Sci. 50, 765–770 (2009).
Zhang, Y., Patil, R. V. & Marmorstein, A. D. Bestrophin 2 is expressed in human non-pigmented ciliary epithelium but not retinal pigment epithelium. Mol. Vis. 16, 200–206 (2010).
Johnson, A. A. et al. Bestrophin 1 and retinal disease. Prog. Retin. Eye Res. 58, 45–69 (2017).
Lee, S. et al. Channel-mediated tonic GABA release from glia. Science 330, 790–796 (2010).
Li, Y. et al. Patient-specific mutations impair BESTROPHIN1’s essential role in mediating Ca2+-dependent Cl− currents in human RPE. eLife 6, e29914 (2017).
Heijl, A., Leske, M. C., Bengtsson, B., Hyman, L. & Hussein, M. Reduction of intraocular pressure and glaucoma progression: results from the Early Manifest Glaucoma Trial. Arch. Ophthalmol. 120, 1268–1279 (2002).
Kass, M. A. et al. The Ocular Hypertension Treatment Study: a randomized trial determines that topical ocular hypotensive medication delays or prevents the onset of primary open-angle glaucoma. Arch. Ophthalmol. 120, 701–713 (2002).
Lichter, P. R. et al. Interim clinical outcomes in the Collaborative Initial Glaucoma Treatment Study comparing initial treatment randomized to medications or surgery. Ophthalmology 108, 1943–1953 (2001).
Owji, A. P. et al. Structural and functional characterization of the bestrophin-2 anion channel. Nat. Struct. Mol. Biol. 27, 382–391 (2020).
Qu, Z., Fischmeister, R. & Hartzell, C. Mouse bestrophin-2 is a bona fide Cl− channel: identification of a residue important in anion binding and conduction. J. Gen. Physiol. 123, 327–340 (2004).
Qu, Z. & Hartzell, H. C. Bestrophin Cl− channels are highly permeable to HCO3. Am. J. Physiol. 294, C1371–C1377 (2008).
Yu, K., Lujan, R., Marmorstein, A., Gabriel, S. & Hartzell, H. C. Bestrophin-2 mediates bicarbonate transport by goblet cells in mouse colon. J. Clin. Invest. 120, 1722–1735 (2010).
Han, K. S. et al. Channel-mediated astrocytic glutamate release via Bestrophin-1 targets synaptic NMDARs. Mol. Brain 6, 4 (2013).
Jo, S. et al. GABA from reactive astrocytes impairs memory in mouse models of Alzheimer’s disease. Nat. Med. 20, 886–896 (2014).
Koh, W. et al. Astrocytes render memory flexible by releasing d-serine and regulating NMDA receptor tone in the hippocampus. Biol. Psychiatry 91, 740–752 (2022).
Kwak, H. et al. Astrocytes control sensory acuity via tonic inhibition in the thalamus. Neuron 108, 691–706.e610 (2020).
Oh, S. J. et al. Protease activated receptor 1-induced glutamate release in cultured astrocytes is mediated by Bestrophin-1 channel but not by vesicular exocytosis. Mol. Brain 5, 38 (2012).
Park, H. et al. High glutamate permeability and distal localization of Best1 channel in CA1 hippocampal astrocyte. Mol. Brain 6, 54 (2013).
Woo, D. H. et al. TREK-1 and Best1 channels mediate fast and slow glutamate release in astrocytes upon GPCR activation. Cell 151, 25–40 (2012).
Kane Dickson, V., Pedi, L. & Long, S. B. Structure and insights into the function of a Ca2+-activated Cl− channel. Nature 516, 213–218 (2014).
Owji, A. P. et al. Structures and gating mechanisms of human bestrophin anion channels. Nat. Commun. 13, 3836 (2022).
Yang, T. et al. Structure and selectivity in bestrophin ion channels. Science 346, 355–359 (2014).
Miller, A. N., Vaisey, G. & Long, S. B. Molecular mechanisms of gating in the calcium-activated chloride channel bestrophin. eLife 8, e43231 (2019).
Hu, R. G., Lim, J. C., Kalloniatis, M. & Donaldson, P. J. Cellular localization of glutamate and glutamine metabolism and transport pathways in the rat ciliary epithelium. Invest. Ophthalmol. Vis. Sci. 52, 3345–3353 (2011).
Langford, M. P. et al. Apical localization of glutamate in GLAST-1, glutamine synthetase positive ciliary body nonpigmented epithelial cells. Clin. Ophthalmol. 1, 43–53 (2007).
Hu, R. G., Zhu, Y., Donaldson, P. & Kalloniatis, M. Alterations of glutamate, glutamine, and related amino acids in the anterior eye secondary to ischaemia and reperfusion. Curr. Eye Res. 37, 633–643 (2012).
Moreno, M. C. et al. Effect of glaucoma on the retinal glutamate/glutamine cycle activity. FASEB J. 19, 1161–1162 (2005).
Krajewski, W. W. et al. Crystal structures of mammalian glutamine synthetases illustrate substrate-induced conformational changes and provide opportunities for drug and herbicide design. J. Mol. Biol. 375, 217–228 (2008).
Yu, D. Y., Noh, S. M. & Lee, G. M. Limitations to the development of recombinant human embryonic kidney 293E cells using glutamine synthetase-mediated gene amplification: Methionine sulfoximine resistance. J. Biotechnol. 231, 136–140 (2016).
Ji, C. et al. Dual Ca2+-dependent gates in human Bestrophin1 underlie disease-causing mechanisms of gain-of-function mutations. Commun. Biol. 2, 240 (2019).
Zhang, Y. et al. ATP activates bestrophin ion channels through direct interaction. Nat. Commun. 9, 3126 (2018).
Rivas, S., Hanif, K., Chakouri, N. & Ben-Johny, M. Probing ion channel macromolecular interactions using fluorescence resonance energy transfer. Methods Enzymol. 653, 319–347 (2021).
Kittredge, A., Ward, N., Hopiavuori, A., Zhang, Y. & Yang, T. Expression and purification of mammalian bestrophin ion channels. J. Vis. Exp. 138, 57832 (2018).
Ji, C. et al. Investigation and restoration of BEST1 activity in patient-derived RPEs with dominant mutations. Sci. Rep. 9, 19026 (2019).
Zhao, Q. et al. Distinct expression requirements and rescue strategies for BEST1 loss- and gain-of-function mutations. eLife 10, e67622 (2021).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Zivanov, J., Nakane, T. & Scheres, S. H. W. A Bayesian approach to beam-induced motion correction in cryo-EM single-particle analysis. IUCrJ 6, 5–17 (2019).
Punjani, A., Zhang, H. & Fleet, D. J. Non-uniform refinement: adaptive regularization improves single-particle cryo-EM reconstruction. Nat. Methods 17, 1214–1221 (2020).
Waterhouse, A. et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46, W296–W303 (2018).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D 60, 2126–2132 (2004).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D 74, 531–544 (2018).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D 66, 12–21 (2010).
Afonine, P. V. et al. New tools for the analysis and validation of cryo-EM maps and atomic models. Acta Crystallogr. D 74, 814–840 (2018).
Barad, B. A. et al. EMRinger: side chain-directed model and map validation for 3D cryo-electron microscopy. Nat. Methods 12, 943–946 (2015).
Pintilie, G. et al. Measurement of atom resolvability in cryo-EM maps with Q-scores. Nat. Methods 17, 328–334 (2020).
Acknowledgements
We thank W. Hendrickson and C. Hartzell for comments on the paper; A. Hsu for helping purify bGS; Y. Kong for helping set up quantitative PCR; M. Ben-Johny and P. J. del Rivero Morfin for helping set up FRET; R. Soni at the Proteomics and Macromolecular Crystallography Shared Resource of Columbia University for helping with mass spectrometry analysis; R. Bruni at Center on Membrane Protein Production and Analysis (COMPPÅ) for providing the bacterial expression vector, and the Unrestricted Grant from Research to Prevent Blindness (RPB) to the Department of Ophthalmology at Columbia University. Cryo-EM data were collected within the Columbia Cryo-EM Core. A.P.O. was supported by NIH grants R01GM107462 and F31EY030763, K.Y. was supported by NIH grants R01EY014852 and R01GM132598, and T.Y. was supported by NIH grants R01GM127652 and R24EY028758, the Irma T. Hirschl/Monique Weill-Caulier Research Award (CU20–4313) and Schaefer Research Award.
Author information
Authors and Affiliations
Contributions
A.P.O. designed research, performed protein purification and cryo-EM experiments, analysed data, made figures and wrote the paper. K.Y. and J.W. designed research, performed patch clamp recordings, analysed data and made figures. A.K. designed research, generated constructs, performed bovine ciliary lysate pull-down, immunoblotting and GS activity experiments, analysed data, made figures, and helped write the paper. Y.Z. and T.Y. designed research, performed experiments, analysed data, made figures and wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks C. Lee and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
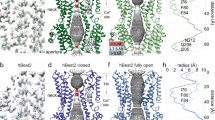
Extended Data Fig. 1 Identification of bBest2-GS interaction by mass spectrometry.
(a,b) Dot plots showing the signal intensities of positive hits from mass spectrometry analysis with ciliary body lysate pull-down samples. X axis, signal intensity in the bBest2 pull-down sample; Y axis, change of signal intensity calculated by subtracting the signal intensity of the control pull-down sample (bovine ciliary lysate with empty Ni+ beads) from that of the bBest2 pull-down sample. The dot representing bGS is labeled red in the total (a) and high intensity (b) data sets.
Extended Data Fig. 2 Immunoblotting of Best2 and GS in HEK293 cells.
(a) Indicated Best2-YFP-His and GS-CFP-Myc constructs were co-expressed and detected by an anti-GFP antibody in input (left) and co-immunoprecipitation (right) samples. The last lane on the IP gel is performed in the presence of 20 mM glutamate. (b) The overall, cytosolic and membrane-bound protein levels of WT hBest2-YFP-His and hGS-CFP-Myc upon individual- or co-expression were detected by an anti-GFP antibody. (c) Exogenous hGS-CFP-Myc and endogenous hGS in HEK293 cells were detected by an anti-GS antibody. (d) The cytosolic and membrane-bound levels of indicated hBest2-YFP-His and hGS-CFP-Myc constructs upon co-expression were detected by an anti-GFP antibody. (e) hBest2-YFP-His and hGS-CFP-Myc were individually co-transfected with hBest2 siRNA and hGS siRNA, respectively, or a scramble siRNA, and detected with an anti-GFP antibody (left). β-Actin (right) served as the loading control on the same blot. Each experiment was biologically replicated three times with similar results.
Extended Data Fig. 3 Cryo-EM single-particle analysis processing workflow for obtaining the bBest2-GS co-structure.
(a) Representative motion-corrected micrograph. (b) Representative 2D class averages after selecting for the complex of interest. The bBest2-GS-Best2 sandwich is solved in cryo-EM with protein overexpression, while only one Best2 to GS is physiologically relevant, as Best2 is a membrane protein and GS is a cytosolic protein that forms polymers (including decamer) in solution in the absence of Best2. (c) The 3DFSC curve. (d) Density map and model for the co-structure viewed perpendicular to the binding interface, with bBest2 (tan) on top (R197 is labeled) and bGS (blue) on the bottom; local filtered map is shown at sigma 7.0. (e) FSC curve for final refinement with D5 symmetry imposed. (f) The viewing angle distribution.
Extended Data Fig. 4 The catalytic region of bGS is not affected by bBest2 binding.
(a) Inner view of the bBest2-GS ion-conducting pathway depicted as a surface and colored by element (N = blue, O = red, C = grey) demonstrating the hydrophilic surface. Box 1 represents the active and ATP-binding site, while box 2 represents the hydrophilic central cavity of bGS. (b) HOLE graph of the ion permeation pathway radius as a function of distance along the central axis of bBest2 and bGS. (c) Cryo-EM density corresponding to box 1 from panel a depicting the model for the ATP/ADP-free active site; local filtered map is shown at sigma 6.8. (d) Cryo-EM density corresponding to box 2 from panel a depicting the hydrated hydrophilic central cavity with density corresponding to water molecules colored red; local filtered map is shown at sigma 6.8. (e) View from inside the bBest2 (tan) aperture looking away from the membrane towards bGS (blue). (f) Side view of the bGS active site with bBest2 sitting on top. The black box indicates R197 of bBest2 interacting with bGS. One protomer of bGS is colored pink with the beta grasp in dark pink. One molecule of ADP (green), taken from the original PDB 2OJW, is placed within the active site at the interface of two adjacent bGS protomers, illustrating the location of the active site in relation to the bBest2 binding site.
Extended Data Fig. 5 Anion currents from hBest2 variants and the influence of hGS.
(a-c) Bar chart showing the steady-state current densities at +100 mV from HEK293 cells transiently expressing the indicated hBest2 and hGS constructs with 1 μM [Ca2+]i under different conditions: (a) Cl− as the principal internal and external anion; n = 9, 5, 7, 7, 5, 8, 5 and 6 for the groups from left to right, respectively; *p = 0.04, 7E-3, 2E-3 and 2E-3 for the groups from left to right, respectively, compared to currents from cells expressing WT hBest2 alone. (b) Internal glutamate and external Cl− as the principal anions; n = 5, 8, 9, 11, 7, 11, 5 and 6 for the groups from left to right, respectively; *p = 3E-4, 9E-4, 0.01 and 0.02 for the groups from left to right, respectively, compared to currents from cells expressing WT hBest2 alone. (c) Internal Cl− and external glutamate as the principal anions; n = 6, 6, 12, 7, 6 and 8 for the groups from left to right, respectively; *p = 5E-4, 6E-6 and 0.047 for the groups from left to right, respectively, compared to currents from cells expressing WT hBest2 alone. All error bars are presented as mean values +/− SEM; p values are calculated by two-tailed unpaired Student’s t test.
Extended Data Fig. 6 Contributions of residues at the Best2-GS interacting interface.
(a) Bar chart showing GS activities in the whole-cell lysates of HEK293 cells transiently transfected with indicated hBest2 and hGS constructs; n = 3 for each group. (b) FRET two-hybrid analysis of the interaction between Cerulean-tagged hGS and Venus-tagged hBest2 constructs. Bar chart showing 1/Kd,EFF of the interaction; n = 3 for each group; *p = 0.01, 0.01, 0.02 and 0.01 for the WT-mutant pairs from left to right, respectively, compared to the WT-WT pair. (c) Bar chart showing GS activities in the cytosolic lysates of HEK293 cells transiently transfected with indicated hGS constructs; n = 3 for each group; *p = 9E-3 and 0.02 for untransfected and hGS R299E transfected cells, respectively, compared to cells transfected with WT hGS. (d) Population steady-state current density-voltage relationships of hBest2 in the presence of hGS G23A or K52A mutant (magenta triangles, n = 7 for each group), compared to hBest2 without (black triangle, n = 9) or with WT hGS (red circle, n = 5) at 1 μM [Ca2+]i in transiently transfected HEK293 cells; *p = 0.04 compared to currents from cells expressing hBest2 alone. All error bars are presented as mean values +/− SEM; p values are calculated by two-tailed unpaired Student’s t test.
Extended Data Fig. 7 Reversal potentials of hBest2 variants and the influence of hGS.
(a-b) Bar chart showing the reversal potentials from HEK293 cells transiently expressing the indicated hBest2 and hGS constructs at 1 μM [Ca2+]i under different conditions: (a) Internal glutamate and external Cl− as the principal anions; n = 5, 8, 9, 11, 7, 11, 5 and 6 for the groups from left to right, respectively; *p = 0.04 and 0.02 for 3A and K208A, respectively, compared to currents from cells expressing WT hBest2 alone. (b) Internal Cl− and external glutamate as the principal anions; n = 6, 6, 12, 7, 6, 8, 9, 6 and 6 for the groups from left to right, respectively. p = 3E-5, 3E-4, 0.01, 2E-3 and 2E-7 for the groups from left to right, respectively, compared to currents from cells expressing WT hBest2 alone. All error bars are presented as mean values +/− SEM; p values are calculated by two-tailed unpaired Student’s t test.
Extended Data Fig. 8 Influence of hGS on the permeability of hBest2 to glutamate and glutamine.
(a,b) Reversal potentials (a) and relative ion permeability ratios of glutamate to Cl− (b) from HEK293 cells transiently expressing hBest2 with 140 mM external glutamate and different concentrations of internal glutamate at 1 μM [Ca2+]i; n = 6, 9, 9, 11, 11 and 9 for the groups from left to right, respectively. (c) Population steady-state current density-voltage relationships in HEK293 cells transiently expressing hBest2 and WT (red, n = 8) or mutant (blue, n = 11 and 7 for G12D and G12W, respectively) hGS at 1 μM [Ca2+]i with internal glutamate and external Cl− as the principal anions; *p = 0.03 for both G12D and G12W, compared to WT GS. (d) Population steady-state current density-voltage relationships in HEK293 cells transiently expressing hBest2 alone (black, n = 10) or hBest2 + hGS (red, n = 9) at 1 μM [Ca2+]i with internal glutamine and external Cl− as the principal anions. (e) The same format as d, but with external glutamine and internal Cl− as the principal anions; n = 11 for each group; *p = 7E-3 compared to hBest2 alone. (f) Reversal potentials calculated from d, e. All error bars are presented as mean values +/− SEM; p values are calculated by two-tailed unpaired Student’s t test.
Extended Data Fig. 9 Knockdown of hBest2 and hGS in human NPE cells.
The relative mRNA levels of hBest2 and hGS in cells treated with a scramble, hBest2-specific or hGS-specific siRNA were analyzed by qRT-PCR and normalized to those in untreated cells; n = 3 for each group; *p = 5E-4 and 2E-3 for hBest2 and hGS mRNA levels in cells treated with hBest2- and hGS-specific siRNA, respectively, compared to those in cells treated with the scramble siRNA by two-tailed unpaired Student’s t test. All error bars are presented as mean values +/− SEM.
Supplementary information
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Owji, A.P., Yu, K., Kittredge, A. et al. Bestrophin-2 and glutamine synthetase form a complex for glutamate release. Nature 611, 180–187 (2022). https://doi.org/10.1038/s41586-022-05373-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05373-x
This article is cited by
-
GABA tone regulation and its cognitive functions in the brain
Nature Reviews Neuroscience (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
