
Abstract
The mutualistic relationship of gut-resident microbiota and the host immune system promotes homeostasis that ensures maintenance of the microbial community and of a largely non-aggressive immune cell compartment1,2. The consequences of disturbing this balance include proximal inflammatory conditions, such as Crohn’s disease, and systemic illnesses. This equilibrium is achieved in part through the induction of both effector and suppressor arms of the adaptive immune system. Helicobacter species induce T regulatory (Treg) and T follicular helper (TFH) cells under homeostatic conditions, but induce inflammatory T helper 17 (TH17) cells when induced Treg (iTreg) cells are compromised3,4. How Helicobacter and other gut bacteria direct T cells to adopt distinct functions remains poorly understood. Here we investigated the cells and molecular components required for iTreg cell differentiation. We found that antigen presentation by cells expressing RORγt, rather than by classical dendritic cells, was required and sufficient for induction of Treg cells. These RORγt+ cells—probably type 3 innate lymphoid cells and/or Janus cells5—require the antigen-presentation machinery, the chemokine receptor CCR7 and the TGFβ activator αv integrin. In the absence of any of these factors, there was expansion of pathogenic TH17 cells instead of iTreg cells, induced by CCR7-independent antigen-presenting cells. Thus, intestinal commensal microbes and their products target multiple antigen-presenting cells with pre-determined features suited to directing appropriate T cell differentiation programmes, rather than a common antigen-presenting cell that they endow with appropriate functions.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
Code availability
All code used for analysis in this manuscript is available at https://github.com/nygctech/Kedmi-CITEseq.
Change history
23 September 2022
A Correction to this paper has been published: https://doi.org/10.1038/s41586-022-05358-w
References
Hooper, L. V., Littman, D. R. & Macpherson, A. J. Interactions between the microbiota and the immune system. Science 336, 1268–1273 (2012).
Ansaldo, E., Farley, T. K. & Belkaid, Y. Control of immunity by the microbiota. Annu. Rev. Immunol. 39, 449–479 (2021).
Xu, M. et al. c-MAF-dependent regulatory T cells mediate immunological tolerance to a gut pathobiont. Nature 554, 373–377 (2018).
Chai, J. N. et al. Helicobacter species are potent drivers of colonic T cell responses in homeostasis and inflammation. Sci. Immunol. 2, eaal5068 (2017).
Wang, J. et al. Single-cell multiomics defines tolerogenic extrathymic Aire-expressing populations with unique homology to thymic epithelium. Sci. Immunol. 6, eabl5053 (2021).
Russler-Germain, E. V. et al. Gut Helicobacter presentation by multiple dendritic cell subsets enables context-specific regulatory T cell generation. eLife 10, e54792 (2021).
Darrasse-Jèze, G. et al. Feedback control of regulatory T cell homeostasis by dendritic cells in vivo. J. Exp. Med. 206, 1853–1862 (2009).
Durai, V. & Murphy, K. M. Functions of murine dendritic cells. Immunity 45, 719–736 (2016).
Esterházy, D. et al. Classical dendritic cells are required for dietary antigen-mediated induction of peripheral Treg cells and tolerance. Nat. Immunol. 17, 545–555 (2016).
Nussenzweig, M. C., Steinman, R. M., Gutchinov, B. & Cohn, Z. A. Dendritic cells are accessory cells for the development of anti-trinitrophenyl cytotoxic T lymphocytes. J. Exp. Med. 152, 1070–1084 (1980).
Sefik, E. et al. Individual intestinal symbionts induce a distinct population of RORγ+ regulatory T cells. Science 349, 993–997 (2015).
Esterhazy, D. et al. Compartmentalized gut lymph node drainage dictates adaptive immune responses. Nature 569, 126–130 (2019).
Worbs, T. et al. Oral tolerance originates in the intestinal immune system and relies on antigen carriage by dendritic cells. J. Exp. Med. 203, 519–527 (2006).
Koscso, B. et al. Gut-resident CX3CR1hi macrophages induce tertiary lymphoid structures and IgA response in situ. Sci. Immunol. 5, eaax0062 (2020).
Mildner, A. & Jung, S. Development and function of dendritic cell subsets. Immunity 40, 642–656 (2014).
Anderson, D. A. 3rd, Dutertre, C. A., Ginhoux, F. & Murphy, K. M. Genetic models of human and mouse dendritic cell development and function. Nat. Rev. Immunol. 21, 101–115 (2021).
Coombes, J. L. et al. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-β and retinoic acid-dependent mechanism. J. Exp. Med. 204, 1757–1764 (2007).
Persson, E. K. et al. IRF4 transcription-factor-dependent CD103+CD11b+ dendritic cells drive mucosal T helper 17 cell differentiation. Immunity 38, 958–969 (2013).
Pool, L., Rivollier, A. & Agace, W. W. Deletion of IRF4 in dendritic cells leads to delayed onset of T cell-dependent colitis. J. Immunol. 204, 1047–1055 (2020).
Wohn, C. et al. Absence of MHC class II on cDC1 dendritic cells triggers fatal autoimmunity to a cross-presented self-antigen. Science Immunol. 5, eaba1896 (2020).
Yamano, T. et al. Aire-expressing ILC3-like cells in the lymph node display potent APC features. J. Exp. Med. 216, 1027–1037 (2019).
Hepworth, M. R. et al. Group 3 innate lymphoid cells mediate intestinal selection of commensal bacteria-specific CD4+ T cells. Science 348, 1031–1035 (2015).
Bartleson, J. M. et al. Strength of tonic T cell receptor signaling instructs T follicular helper cell-fate decisions. Nat. Immunol. 21, 1384–1396 (2020).
Mackley, E. C. et al. CCR7-dependent trafficking of RORγ+ ILCs creates a unique microenvironment within mucosal draining lymph nodes. Nat. Commun. 6, 5862 (2015).
Kim, M. H., Taparowsky, E. J. & Kim, C. H. Retinoic acid differentially regulates the migration of innate lymphoid cell subsets to the gut. Immunity 43, 107–119 (2015).
Wang, R. et al. GARP regulates the bioavailability and activation of TGFβ. Mol. Biol. Cell 23, 1129–1139 (2012).
Lienart, S. et al. Structural basis of latent TGF-β1 presentation and activation by GARP on human regulatory T cells. Science 362, 952–956 (2018).
Qin, Y. et al. A milieu molecule for TGF-β required for microglia function in the nervous system. Cell 174, 156–171.e116 (2018).
Lacy-Hulbert, A. et al. Ulcerative colitis and autoimmunity induced by loss of myeloid alphav integrins. Proc. Natl Acad. Sci. USA 104, 15823–15828 (2007).
Paidassi, H. et al. Preferential expression of integrin αVβ8 promotes generation of regulatory T cells by mouse CD103+ dendritic cells. Gastroenterology 141, 1813–1820 (2011).
Travis, M. A. et al. Loss of integrin αVβ8 on dendritic cells causes autoimmunity and colitis in mice. Nature 449, 361–365 (2007).
Gardner, J. M. et al. Deletional tolerance mediated by extrathymic Aire-expressing cells. Science 321, 843–847 (2008).
Nakawesi, J. et al. αVβ8 integrin-expression by BATF3-dependent dendritic cells facilitates early IgA responses to Rotavirus. Mucosal Immunol. 14, 53–67 (2021).
Brown, C. C. et al. Transcriptional basis of mouse and human dendritic cell heterogeneity. Cell 179, 846–863.e824 (2019).
Barnett, L. G. et al. B cell antigen presentation in the initiation of follicular helper T cell and germinal center differentiation. J. Immunol. 192, 3607–3617 (2014).
Hepworth, M. R. et al. Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria. Nature 498, 113–117 (2013).
Goto, Y. et al. Segmented filamentous bacteria antigens presented by intestinal dendritic cells drive mucosal Th17 cell differentiation. Immunity 40, 594–607 (2014).
Yin, X., Chen, S. & Eisenbarth, S. C. Dendritic cell regulation of T helper cells. Annu. Rev. Immunol. 39, 759–790 (2021).
Eberl, G. & Littman, D. R. Thymic origin of intestinal αβ T cells revealed by fate mapping of RORγt+ cells. Science 305, 248–251 (2004).
Lochner, M. et al. In vivo equilibrium of proinflammatory IL-17+ and regulatory IL-10+Foxp3+RORγt+ T cells. J. Exp. Med. 205, 1381–1393 (2008).
Kaplan, D. H., Jenison, M. C., Saeland, S., Shlomchik, W. D. & Shlomchik, M. J. Epidermal Langerhans cell-deficient mice develop enhanced contact hypersensitivity. Immunity 23, 611–620 (2005).
Dow, L. E. et al. Conditional reverse Tet-transactivator mouse strains for the efficient induction of TRE-regulated transgenes in mice. PLoS ONE 9, e95236 (2014).
Metzger, T. C. et al. Lineage tracing and cell ablation identify a post-Aire-expressing thymic epithelial cell population. Cell Rep. 5, 166–179 (2013).
Stoeckius, M. et al. Simultaneous epitope and transcriptome measurement in single cells. Nat. Methods 14, 865–868 (2017).
Stoeckius, M. et al. Cell hashing with barcoded antibodies enables multiplexing and doublet detection for single cell genomics. Genome Biol. 19, 224 (2018).
van Buggenum, J. A. et al. A covalent and cleavable antibody–DNA conjugation strategy for sensitive protein detection via immuno-PCR. Sci Rep. 6, 22675 (2016).
Hafemeister, C. & Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 20, 296 (2019).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e3529 (2021).
Waltman, L., & Nees, J. v. E. A smart local moving algorithm for large-scale modularity-based community detection. Eur. Phys. J. B 86, 471 (2013).
Wolock, S. L., Lopez, R. & Klein, A. M. Scrublet: computational identification of cell doublets in single-cell transcriptomic data. Cell Syst. 8, 281–291.e289 (2019).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Lopez, R., Regier, J., Cole, M. B., Jordan, M. I. & Yosef, N. Deep generative modeling for single-cell transcriptomics. Nat. Methods 15, 1053–1058 (2018).
Acknowledgements
We thank members of the Littman laboratory, J. J. Lafaille and S, Schwab for valuable discussion and critical reading of the manuscript and G. Romero-Meza for assistance with experiments; D. Sheppard for advice and providing ADWA-11 blocking antibody; S. Y. Kim and the NYU Rodent Genetic Engineering Laboratory (RGEL) for rederivation of mutant mice; and C. Loomis and the Experimental Pathology Research Laboratory of NYULMC for histology of intestine samples. The Microscopy Core and the Genome Technology Core are partially supported by NYU Cancer Center Support Grant NIH/NCI P30CA016087 at the Laura and Isaac Perlmutter Cancer Center, S10 RR023704-01A1 and NIH S10 ODO019974-01A1. The Experimental Pathology Research Laboratory is supported by National Institutes of Health Shared Instrumentation grants S10OD010584-01A1 and S10OD018338-01. C.A.L., A.T.S. and J.M.G. are recipients of the IGVF award UM1HG012076. This work was supported by an Irvington Institute fellowship from the Cancer Research Institute (R.K.) and a Jane Coffin Childs Fund fellowship (K.R.M.), the Agence Nationale de la Recherche (ANR-20-CE15-0015) (H.P.), the Helen and Martin Kimmel Center for Biology and Medicine (D.R.L.); National Institutes of Health grants R01AI139540 (P.M.A.) and R01AI158687 (D.R.L.) and the Howard Hughes Medical Institute (D.R.L.).
Author information
Authors and Affiliations
Contributions
R.K., T.A.N., K.R.M. and D.R.L. designed the study and analysed the data. R.K. and T.A.N. performed mouse genetic experiments with assistance from A.G. M.P., M.X. and J.T. performed early experiments to launch the study. K.R.M. and R.K. performed intravital multiphoton microscopy. R.K., S.H. and M.S. performed CITE-seq studies. A.T.S. and C.A.L. performed scRNA-seq. R.K., L.K., Y.H. and J.M.G. did bioinformatics analyses. J.W., M.S.A. and J.M.G. provided biological samples, genomics data and experimental support. H.P., T.M.L., I.A., J.M.B. and P.M.A. contributed mouse strains. H.P. contributed phenotypic analysis. R.K. and D.R.L. wrote the manuscript, with input from the other authors. D.R.L. supervised the research.
Corresponding author
Ethics declarations
Competing interests
D.R.L. consults and has equity interest in Chemocentryx, Vedanta, Immunai and Pfizer Pharmaceuticals. All other authors declare no competing interests.
Peer review
Peer review information
Nature thanks the anonymous reviewer(s) for their contribution to the peer review of this work. Peer review reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Cells targeted by CD11c-Cre and consequences for Hh-specific T cell differentiation.
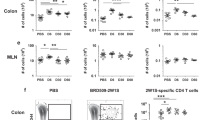
a, Phenotype of Hh7-2 TCR transgenic T cells in the colon lamina propria at 10 days after transfer into Hh-colonized MHCIIΔCD11c (n = 3) and control mice (n = 7), as indicated. b, Phenotype of host CD4+ T cells from mice in (a); MHCIIΔCD11c (n = 3) and control mice (n = 10), as indicated. c, Cytokine profile of Hh7-2 T cells shown in (a); MHCIIΔCD11c (n = 4) and control mice (n = 3). d, Proliferation and differentiation of Hh-specific iTreg and Th17 cells in the MLN of Ccr7 −/− (n = 5) and littermate control mice (n = 5). CFSE-labeled Hh7-2 T cells were analyzed at 3 days following their adoptive transfer into Hh-colonized mice. Data summarize two independent experiments. e–f, Transcription factor (e) and intracellular cytokine (f) profiles of Hh7-2 T cells in the large intestine of Ccr7ΔCD11c (n = 7 or 5, for transcription factors and cytokines, respectively) and littermate control (n = 5) mice, at 10 days after adoptive transfer. g–i, Proportion in MLN of Hh7-2 with the iTreg phenotype at 3 days after transfer into BATF3−/− (g) (n = 7), IRF4ΔCD11c (h) (n = 6), and huLangerin (CD207)-DTA (i) (n = 12) mice (red) and indicated littermate controls (black). Data summarize at least two independent experiments. Representative flow panels and aggregate data are shown for each analysis. All statistics were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 2 Identification of CITE-seq-assigned clusters of sorted tdTomato-ONCD11c fate-mapped cells.
a, Stacked violin plots for selected (curated) and top DEG (data-driven) of tdTomato+ cells sorted from MLN of Hh-colonized mice. b, Stacked violin plots for selected (curated) cell surface markers for each cluster.
Extended Data Fig. 3 Phenotypic discrimination of ILC3 and JC.
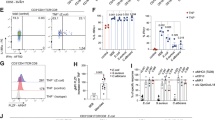
a, Dot plots for selected (curated) DEG and cell surface markers for the indicated clusters, obtained from CITE-seq analysis of tdTomato-ONCD11c fate-mapped cells. b, Flow cytometry profiling of CXCR6, CD127(IL-7R), CCR6 CD25 and CD40 on ILC3 (red) and JC (blue), pre-gated on TCRβ−, TCRγδ−, B220−, RORγt+, MHCII+. c, gating strategy for JC using cell surface staining as indicated. d, Flow cytometry profiling of JC and DC markers, showing that migratory cDC are excluded from CD11clow CCR6+ gating. e, TdTomato levels in ILC3 (TCRβ−, TCRγδ−, B220−, MHCII+ CCR6+, Il7R+) and JC (TCRβ−, TCRγδ−, B220−, MHCII+ CCR6+, Il7R−) from the MLN of Hh-colonized tdTomato-ONCD11c fate-map mice.
Extended Data Fig. 4 Antigen presentation by RORγt+ cells is required for microbiota-induced iTreg cell differentiation.
a, MHCII expression in RORγt+ cells (top) and DC (bottom) from the MLN of Hh-colonized MHCII ΔCD11c mice (n = 6 and 5) and littermate controls (n = 10 and 8). RORγt+ cells were gated as TCRβ−, TCRγδ−, B220−, RORγt+; DC were gated as TCRβ−, TCRγδ−, B220−, CD90−, CD11chigh. b, Bar graph showing frequency of iTreg among Hh7-2 T cells, measured as in Fig. 2e. c-d, Representative dot plots showing Hh7-2 T cell differentiation (c) and cytokine (d) profiles in colon lamina propria at 22 days after adoptive transfer into Hh-colonized MHCII ΔRORγt and littermate controls. e, Representative and aggregate data of transcription factor profiles of host CD4+ T cells in colon lamina propria of mice shown in (c) and (d). f, Hh7-2 cell proliferation and differentiation in the MLN of H2-DMaΔRORγt (RORγt-Cre; H2-Dma f/f) (n = 11) and littermate controls (RORγt-Cre; H2-DMa+/f) (n = 5) at 3 days after transfer of CFSE labeled naïve Hh7-2, cell proliferation and FoxP3 were assessed in cells isolated from C1 MLN. Representative flow cytometry (left) and aggregate data from multiple animals (right). Data summarize two independent experiments. All statistics were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 5 Differential requirements for CCR7 in iTreg and effector Th17 cell differentiation and analysis of Aire+ JC function in differentiation of Hh-specific iTreg cells.
a, Cell surface expression of CCR7 on CD11c-Cre fate-mapped ILC3 (TCRβ−, TCRγδ−, B220−, MHCII+, CCR6+, IL-7R+) and JC (TCRβ−, TCRγδ−, B220−, MHCII+, CCR6+, IL-7R−) in the MLN. b-c, Analysis of DC counts in MLN (b) and large intestine (c) of WT and Ccr7ΔRORγt mixed bone marrow chimeric mice described in Fig. 3c. Counts in the MLN of DC subsets derived from bone marrow (b); frequencies of CCR7+ among total colonic DCs (c) (n = 4). Statistics were calculated using paired two-sided t-test. d, Analysis of CD45.2 frequencies within donor cells is presented for ILC3 (TCRβ−, TCRγδ−, B220−, MHCII+, RORγt +, IL-7R+) and JC (TCRβ−, TCRγδ−, B220−, MHCII+, RORγt +, IL-7R−) in the MLN of WT and Ccr7ΔRORγt mixed bone marrow chimeric mice described in Fig. 3c. e, Cell surface expression of CCR7 in colonic ILC3 (TCRβ−, TCRγδ−, B220−, CD90+, RORγt+, CD25+, CD4+) from Ccr7ΔRORγt (n = 3), Ccr7ΔCD11c (n = 2) and control Hh-colonized mice (n = 2). f, Cell surface expression of CCR7 and CD11b in ILC3-gated MLN cells (TCRβ−, TCRγδ−, B220−, IL-7R+, CCR6+, CD25+) from Ccr7ΔCD11c (n = 4) and control Hh-colonized mice (n = 4). g, Lethally irradiated mice were reconstituted with BM cells from CD45.2 Aire-DTR or CD45.2 WT mice. One month after reconstitution, mice were colonized with Hh, and one week later were treated with Diphtheria toxin (DT, Sigma-Aldrich) for 3 sequential days (at a dose of 25 ng/g mice). CD45.1/CD45.2 CFSE-labeled Hh7-2 T cells (1 x 105) were transferred intravenously into the mice on the first day of DT treatment. Bar graph of proportion of proliferating Foxp3+ Hh7-2 T cells in the MLN of mice reconstituted with Aire-DTR BM (n = 5) or with WT BM (n = 4) (left); Aire mRNA in the spleen of the treated mice was blindly scored using RNAscope analysis. The experiment was performed once. h, Proliferation and differentiation of CFSE-labeled Hh7-2 T cells in the MLN of RORγt-Cre;Aire f/f (n = 5) and control Aire+/f littermates (n = 7) at 3 days after adoptive transfer. Data summarize three independent experiments. All statistics, except for b and c, were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 6 Effect of integrin αvβ8 blockade or αv inactivation on microbiota-dependent T cell differentiation.
a, Expression of integrin αv (CD51) in fate-mapped RORγt+ cell subsets from MLN of wild type and Itgav ΔRORγt mice. b, C1 MLN from ItgavΔCD11c (n = 3) and littermate controls (n = 4), 10 days after Hh colonization. c-d, Flow cytometry profiling of transcription factors and CCR6 in proliferating CFSE-labeled Hh7-2 in the MLN at 3 days after adoptive transfer into Itgav ΔRORγt (n = 3) and littermate control mice (n = 3) (b) or into mice treated with ADWA11(n = 4) (as in Fig. 4a) or untreated control littermates (n = 4) (c). Summary data of results in (b) and (c) are shown below. e, Intracellular IFNγ and T-BET expression in PMA/Ionomycin-stimulated Hh7-2 T cells isolated from colon lamina propria of Itgav ΔRORγt(n = 3) and control littermates (n = 5), 10 days after adoptive transfer. f, Frequency of iTreg cells among proliferating Hh7-2 in the MLN at 3 days after adoptive transfer into Itgav ΔCD4 (n = 4) and control littermates (n = 7). Data summarize two independent experiments. g, Integrin αv and MHCII cell surface expression in ILC3 (gated as TCRβ−, TCRγδ−, B220−, RORγt+, CD90+, CD25+ CD45.2+) isolated from MLN of bone marrow chimeric mice, reconstituted with different combinations of donor cells as indicated and colonized with Hh for 10 days. Data summarized below for control (n = 3), MHCIIΔCD11c (n = 3), MHCIIΔCD11c and WT (n = 4), and MHCIIΔCD11c and ItgavΔRORγt (n = 4) reconstituted mice. All Statistics were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 7 Itgav and Itgb8 expression in ILC3 and JC.
a, tSNE plot with Leiden clustering of scRNAseq of pooled GFP+ sorted and unsorted cells, as indicated, from pooled lymph nodes of Adig mice32. b, tSNE feature plots showing Aire, Itgav, and Itgb8 levels in the cell clusters. c, UMAP plot of Aire+ JC and ILC3 populations from pooled datasets as indicated with associated feature plots. d, top differentially expressed genes per pseudobulk cluster in (c), shown by heatmap. e, dot plot of selected genes in JC and ILC3 clusters. f-g, aggregate results (f) and representative flow cytometry (g) of tdTomato+ JC and ILC3, gated as indicated, in C1 MLN of Itgb8-IRES-tdTomato mice33 (n = 4) and littermate controls (n = 4). Aggregate data (right) show percent tdTomato+ cells among total ILC3 and JC and number of reporter-positive cells in the C1 MLN of each mouse. We performed two independent experiments and data shown are from one representative experiment. All statistics were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 8 Analysis of RORγt-expressing cells in the MLN and intravital tracking of RORγt-expressing cells and DC interactions with Hh-specific T cells during priming in the MLN.
a, Hh7-2 proliferation and differentiation in MLN of Ccr7Δzbtb46 (n = 4) and control littermates (n = 5), at 3 days after transfer of the naïve cells. Data in the right panel summarize three independent experiments. b, UMAP visualization of CITE-seq datasets obtained from 3 distinct sorted populations (GFP+, GFP+ tdTomato+ and tdTomato+) isolated from C1 MLN of Zbtb46-eGFP ; tdTomato-ONRORγt mice (n = 2), analyzed by the WNN method. c, Flow cytometry analysis of fate-mapped C1 MLN cells from RORγt-eGFP;mKate2-ONzbtb46 mice, gated for the indicated cell subsets. d-e, Feature plot showing Rorc (d) and integrin αv (e) levels in the cell clusters identified in the CITE-seq analysis shown in (b); Positive cells are layered in front. g, Flow cytometry analysis of fate-mapped C1 MLN cells from RORγt-eGFP;mKate2-ONzbtb46 mice, gated for the indicated cell subsets (ILC3 were gated as TCRβ−, TCRγδ−, B220−, MHCII+, RORγt-eGFP+, CCR6+, CD25+ and JC as TCRβ−, TCRγδ−, B220−, MHCII+, RORγt-eGFP+, CD25−). Note that there is incomplete excision of the transcriptional stop signal by zbtb46-Cre. h, Representative image of cell-cell interactions of recently primed Hh-specific T cells with DC and RORγt-expressing cells. Nur77-eGFP tracer-labeled Hh7-2 T cells were transferred into of RORγt-eGFP;mKate2-ONzbtb46 Hh-colonized mice. Cell colocalization of primed Hh7-2 (tracer dye+, GFP+) or naïve Hh7-2 (tracer dye+, GFP−) T cells with cDC (mKate2+ with dendritic morphology), RORγt-expressing cells (eGFP+, mKate2+ or eGFP+ alone with amoeboid morphology), or both were visualized using intravital multiphoton microscopy of the C1 MLN at 15 h after transfer. Note that Cell Tracer fluorescent labeling provides clear spatial discrimination of RORγt-eGFP and Nur77-eGFP expressing cells. i, Quantification and graphical representation of the total (left) and individual rates of interaction of RORγt-expressing cells or cDC populations (right) with primed or naïve Hh7-2 T cells. Data summarize cell-cell interactions from six 0.25mm3 three-dimensional regions of C1 MLN, (n = 72 total Hh7-2 T cells), (n = 49 primed and 23 naïve Hh7-2 T cells). All statistics were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 9 Gain-of-function expression of MHCII in RORγt+ cells rescues bone marrow-derived iTreg cell differentiation.
a, Aggregate data showing MHCII frequency on donor-derived DC and RORγt+ cells in MLN and colon lamina propria from chimeric mice reconstituted with combinations of donor BM cells as indicated, with representative flow cytometry panel in Fig. 5b. MLN: WT (n = 5), MHCIIΔCD11c (n = 5), MHCIIΔCD11c and WT (n = 8), MHCIIΔCD11c and MHCII-ONRORγt (n = 7). Colon: WT (n = 4), MHCIIΔCD11c (n = 6), MHCIIΔCD11c and WT (n = 8), MHCIIΔCD11c and MHCII-ONRORγt (n = 6). b, Donor bone marrow-derived CD4+ T cell differentiation in colon lamina propria from chimeric mice reconstituted with combinations of BM cells as indicated. Representative flow panels (left) and aggregate data (right). WT (n = 10), MHCIIΔCD11c (n = 11), MHCIIΔCD11c and WT (n = 8), MHCIIΔCD11c and MHCII-ONRORγt (n = 9). Colon: WT (n = 4), MHCIIΔCD11c (n = 6), MHCIIΔCD11c and WT (n = 8), MHCIIΔCD11c and MHCII-ONRORγt (n = 7). c, Representative flow cytometry (left) and aggregate data (right) of Hh7-2 T cell differentiation in colon lamina propria of Hh-colonized bone marrow chimeric mice reconstituted with cells of indicated genotypes, 12 days after transfer of naive TCR transgenic T cells. WT (n = 8), MHCIIΔCD11c (n = 7), MHCII-ONCD11c (n = 5), and MHCII-ONRORγt (n = 4). Data summarize two or three independent experiments. All statistics were calculated by unpaired two-sided Welch’s t-test. Error bars denote mean ± s.d. p-values are indicated in the figure.
Extended Data Fig. 10 Schematic of the requirement of distinct APC subsets for T cell differentiation.
CCR7 and integrin αvβ8 are required in RORγt+ APCs for iTreg cell differentiation. Note that other APCs, with differential requirements for CCR7 expression, are involved in the priming and differentiation of pathogenic Th17 and Tfh cells.
Supplementary information
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kedmi, R., Najar, T.A., Mesa, K.R. et al. A RORγt+ cell instructs gut microbiota-specific Treg cell differentiation. Nature 610, 737–743 (2022). https://doi.org/10.1038/s41586-022-05089-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05089-y
This article is cited by
-
Impaired immune tolerance mediated by reduced Tfr cells in rheumatoid arthritis linked to gut microbiota dysbiosis and altered metabolites
Arthritis Research & Therapy (2024)
-
The dynamic shifts of IL-10-producing Th17 and IL-17-producing Treg in health and disease: a crosstalk between ancient "Yin-Yang" theory and modern immunology
Cell Communication and Signaling (2024)
-
Immune microniches shape intestinal Treg function
Nature (2024)
-
The emerging family of RORγt+ antigen-presenting cells
Nature Reviews Immunology (2024)
-
Innate lymphoid cells and innate-like T cells in cancer — at the crossroads of innate and adaptive immunity
Nature Reviews Cancer (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
