
Abstract
Gut commensal bacteria with the ability to translocate across the intestinal barrier can drive the development of diverse immune-mediated diseases1,2,3,4. However, the key factors that dictate bacterial translocation remain unclear. Recent studies have revealed that gut microbiota strains can adapt and evolve throughout the lifetime of the host5,6,7,8,9, raising the possibility that changes in individual commensal bacteria themselves over time may affect their propensity to elicit inflammatory disease. Here we show that within-host evolution of the model gut pathobiont Enterococcus gallinarum facilitates bacterial translocation and initiation of inflammation. Using a combination of in vivo experimental evolution and comparative genomics, we found that E. gallinarum diverges into independent lineages adapted to colonize either luminal or mucosal niches in the gut. Compared with ancestral and luminal E. gallinarum, mucosally adapted strains evade detection and clearance by the immune system, exhibit increased translocation to and survival within the mesenteric lymph nodes and liver, and induce increased intestinal and hepatic inflammation. Mechanistically, these changes in bacterial behaviour are associated with non-synonymous mutations or insertion–deletions in defined regulatory genes in E. gallinarum, altered microbial gene expression programs and remodelled cell wall structures. Lactobacillus reuteri also exhibited broadly similar patterns of divergent evolution and enhanced immune evasion in a monocolonization-based model of within-host evolution. Overall, these studies define within-host evolution as a critical regulator of commensal pathogenicity that provides a unique source of stochasticity in the development and progression of microbiota-driven disease.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
The sequencing data generated in this study are available at the NCBI Sequence Read Archive (SRA) database. Genomic sequences are under the accession PRJNA743649 and RNA-seq data are under the accession PRJNA743979. Source data are provided with this paper.
References
Manfredo Vieira, S. et al. Translocation of a gut pathobiont drives autoimmunity in mice and humans. Science 359, 1156–1161 (2018).
Nakamoto, N. et al. Gut pathobionts underlie intestinal barrier dysfunction and liver T helper 17 cell immune response in primary sclerosing cholangitis. Nat. Microbiol. 4, 492–503 (2019).
Chow, J., Tang, H. & Mazmanian, S. K. Pathobionts of the gastrointestinal microbiota and inflammatory disease. Curr. Opin. Immunol. 23, 473–480 (2011).
Ruff, W. E., Greiling, T. M. & Kriegel, M. A. Host-microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 18, 521–538 (2020).
Schloissnig, S. et al. Genomic variation landscape of the human gut microbiome. Nature 493, 45–50 (2013).
Zhao, S. et al. Adaptive evolution within gut microbiomes of healthy people. Cell Host Microbe 25, 656–667 (2019).
Elhenawy, W., Tsai, C. N. & Coombes, B. K. Host-specific adaptive diversification of Crohn's disease-associated adherent-invasive Escherichia coli. Cell Host Microbe 25, 301–312 (2019).
Barroso-Batista, J. et al. Specific eco-evolutionary contexts in the mouse gut reveal Escherichia coli metabolic versatility. Curr. Biol. 30, 1049–1062 (2020).
Yilmaz, B. et al. Long-term evolution and short-term adaptation of microbiota strains and sub-strains in mice. Cell Host Microbe 29, 650–663 (2021).
Group, N. H. W. et al. The NIH Human Microbiome Project. Genome Res. 19, 2317–2323 (2009).
The Integrative HMP (iHMP) Research Network Consortium. The Integrative Human Microbiome Project. Nature 569, 641–648 (2019).
Plumbridge, J. Control of the expression of the manXYZ operon in Escherichia coli: Mlc is a negative regulator of the mannose PTS. Mol. Microbiol. 27, 369–380 (1998).
Honeyman, A. L. & Curtiss, R. 3rd Isolation, characterization and nucleotide sequence of the Streptococcus mutans lactose-specific enzyme II (lacE) gene of the PTS and the phospho-beta-galactosidase (lacG) gene. J. Gen. Microbiol. 139, 2685–2694 (1993).
Dubrac, S. & Msadek, T. Tearing down the wall: peptidoglycan metabolism and the WalK/WalR (YycG/YycF) essential two-component system. Adv. Exp. Med. Biol. 631, 214–228 (2008).
Fried, L., Behr, S. & Jung, K. Identification of a target gene and activating stimulus for the YpdA/YpdB histidine kinase/response regulator system in Escherichia coli. J. Bacteriol. 195, 807–815 (2013).
Dobihal, G. S., Brunet, Y. R., Flores-Kim, J. & Rudner, D. Z. Homeostatic control of cell wall hydrolysis by the WalRK two-component signaling pathway in Bacillus subtilis. eLife 8, e52088 (2019).
Auchtung, J. M., Lee, C. A., Garrison, K. L. & Grossman, A. D. Identification and characterization of the immunity repressor (ImmR) that controls the mobile genetic element ICEBs1 of Bacillus subtilis. Mol. Microbiol. 64, 1515–1528 (2007).
Donaldson, G. P., Lee, S. M. & Mazmanian, S. K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 14, 20–32 (2016).
Boneca, I. G. et al. A critical role for peptidoglycan N-deacetylation in Listeria evasion from the host innate immune system. Proc. Natl Acad. Sci. USA 104, 997–1002 (2007).
Benachour, A. et al. The lysozyme-induced peptidoglycan N-acetylglucosamine deacetylase PgdA (EF1843) is required for Enterococcus faecalis virulence. J. Bacteriol. 194, 6066–6073 (2012).
Thurlow, L. R., Thomas, V. C., Fleming, S. D. & Hancock, L. E. Enterococcus faecalis capsular polysaccharide serotypes C and D and their contributions to host innate immune evasion. Infect. Immun. 77, 5551–5557 (2009).
Dalton, J. E. et al. Intraepithelial γδ+ lymphocytes maintain the integrity of intestinal epithelial tight junctions in response to infection. Gastroenterology 131, 818–829 (2006).
Hoytema van Konijnenburg, D. P. et al. Intestinal epithelial and intraepithelial T cell crosstalk mediates a dynamic response to infection. Cell 171, 783–794 (2017).
Olivares-Villagomez, D. & Van Kaer, L. Intestinal intraepithelial lymphocytes: sentinels of the mucosal barrier. Trends Immunol. 39, 264–275 (2018).
McPherson, A. C., Pandey, S. P., Bender, M. J. & Meisel, M. Systemic immunoregulatory consequences of gut commensal translocation. Trends Immunol. 42, 137–150 (2021).
Zegarra-Ruiz, D. F. et al. A diet-sensitive commensal Lactobacillus strain mediates TLR7-dependent systemic autoimmunity. Cell Host Microbe 25, 113–127 (2019).
Lee, S. M. et al. Bacterial colonization factors control specificity and stability of the gut microbiota. Nature 501, 426–429 (2013).
Brown, S. P., Cornforth, D. M. & Mideo, N. Evolution of virulence in opportunistic pathogens: generalism, plasticity, and control. Trends Microbiol. 20, 336–342 (2012).
Culyba, M. J. & Van Tyne, D. Bacterial evolution during human infection: adapt and live or adapt and die. PLoS Pathog. 17, e1009872 (2021).
Ha, C. W. Y. et al. Translocation of viable gut microbiota to mesenteric adipose drives formation of creeping fat in humans. Cell 183, 666–683(2020).
Van Tyne, D. et al. Impact of antibiotic treatment and host innate immune pressure on enterococcal adaptation in the human bloodstream. Sci. Transl. Med. 11, eaat8418 (2019).
Young, B. C. et al. Severe infections emerge from commensal bacteria by adaptive evolution. eLife 6, e30637 (2017).
Palm, N. W. et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 158, 1000–1010 (2014).
Wick, R. R., Judd, L. M., Gorrie, C. L. & Holt, K. E. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13, e1005595 (2017).
Andrews, S. FastQC: a quality control tool for high throughput sequence data (2010); http://www.bioinformatics.babraham.ac.uk/projects/fastqc/
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
De Coster, W., D'Hert, S., Schultz, D. T., Cruts, M. & Van Broeckhoven, C. NanoPack: visualizing and processing long-read sequencing data. Bioinformatics 34, 2666–2669 (2018).
Aziz, R. K. et al. The RAST Server: rapid annotations using subsystems technology. BMC Genomics 9, 75 (2008).
Overbeek, R. et al. The SEED and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 42, D206–D214 (2014).
Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069 (2014).
Seemann, T. snippy: fast bacterial variant calling from NGS reads (2015); https://github.com/tseemann/snippy
Deatherage, D. E. & Barrick, J. E. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol. Biol. 1151, 165–188 (2014).
Treangen, T. J., Ondov, B. D., Koren, S. & Phillippy, A. M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 15, 524 (2014).
Bankevich, A. et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477 (2012).
Darling, A. C., Mau, B., Blattner, F. R. & Perna, N. T. Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 14, 1394–1403 (2004).
Okonechnikov, K., Golosova, O. & Fursov, M., team, U. Unipro UGENE: a unified bioinformatics toolkit. Bioinformatics 28, 1166–1167 (2012).
Letunic, I. & Bork, P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, W293–W296 (2021).
Page, A. J. et al. Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics 31, 3691–3693 (2015).
Eren, A. M. et al. Community-led, integrated, reproducible multi-omics with anvi’o. Nat. Microbiol. 6, 3–6 (2021).
Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17, 10–12 (2011).
Howe, K. L. et al. Ensembl 2021. Nucleic Acids Res. 49, D884–D891 (2021).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Cullen, T. W. et al. Antimicrobial peptide resistance mediates resilience of prominent gut commensals during inflammation. Science 347, 170–175 (2015).
Leenhouts, K. et al. A general system for generating unlabelled gene replacements in bacterial chromosomes. Mol. Gen. Genet. 253, 217–224 (1996).
Laute-Caly, D. L. et al. The flagellin of candidate live biotherapeutic Enterococcus gallinarum MRx0518 is a potent immunostimulant. Sci. Rep. 9, 801 (2019).
Acknowledgements
We thank R. Medzhitov, A. Wang and the members of the Palm laboratory for discussions and comments; Y. Cao, T. A. Rice and S. R. Leopold for experimental assistance; the staff at the Centre for Cellular and Molecular Imaging (CCMI) Electron Microscopy Facility at Yale University for electron micrograph acquisition; members of the Yale Research Histology Core for histological staining; and staff at the Yale Centre for Genome Analysis for next-generation sequencing. This work was supported by the National Institute on Aging of the National Institutes of Health under award number R01AG068863 (to N.W.P.). Approximately 50% of the funding for this research project (around US$300,000) was financed with NIH funds; the remainder was financed by non-governmental sources. N.W.P. also acknowledges support from the Common Fund of the NIH (DP2DK125119), the Leona M. and Henry B. Helmsley Charitable Trust (3083), the Chan Zuckerberg Initiative, Aligning Science Across Parkinson's through the Michael J. Fox Foundation for Parkinson’s Research, the Ludwig Family, the Mathers Foundation, the Pew Charitable Trust, the NIGMS (RM1GM141649) and F. Hoffmann-La Roche. M.A.K. acknowledges support from the Lupus Research Alliance. The funders of this work had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. Schematic figures were created using BioRender.com.
Author information
Authors and Affiliations
Contributions
N.W.P. and Y.Y. conceived the project, designed the experiments and wrote the manuscript. Y.Y., M.N., V.K., N.D.S. and H.C. designed and performed experiments. M.N. isolated E. gallinarum isolates from SPF NZW × BXSB F1 mice. V.K. generated the pgdA mutant strain. N.D.S. and H.C. assessed potential E. gallinarum phenotypes. Y.Y. performed all of the other experiments and all data analyses. A.L.M. assisted with gnotobiotic mouse experiments. M.A.K. participated in the conceptualization of the project and provided NZW × BXSB F1 mice, the original liver isolate of E. gallinarum from autoimmune-prone SPF mice and critical intellectual input. All of the authors edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
N.W.P. is a co-founder of Artizan Biosciences and Design Pharmaceuticals and has received research funding from Artizan Biosciences and F. Hoffmann-La Roche. M.A.K. holds a patent on the use of microbiota manipulations to treat immune-mediated diseases (US patent no. PCT/US18/14368) and consults for Eligo Biosciences. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks Kenya Honda and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Mutations detected in E. gallinarum isolates from SPF NZW × BXSB F1 mice.
a, Pangenome of eighteen E. gallinarum strains. b, Schematic of the complete genome of E. gallinarum strain EGF1-FE1. c, Number of singleton or shared variants detected in E. gallinarum faecal or liver isolates. d, Functional classes of genes with nonsynonymous mutations or indels. n = 13 genes. e, Relative abundance of liver and faecal E. gallinarum isolates in the faeces of co-colonized SPF C57BL/6 mice. Mice were gavaged with equal doses of EGF1-LV1 and EGF1-FE4 and faecal samples were analysed one week after colonization. n = 6 mice across two independent experiments. Data represent mean ± SEM. Unpaired two-tailed t-test (e). NSY, nonsynonymous SNVs; SYN, synonymous SNVs; Indel, small insertion/deletion variant; Inter, intergenic variant. EG, E. gallinarum.
Extended Data Fig. 2 Mutations detected in E. gallinarum isolates from monocolonized C57BL/6 mice.
a, Schematic of cage setup of the experimental evolution in monocolonized mice. b, Distribution of detected mutations across the E. gallinarum genome. n = 159 unique mutations detected in 153 isolates. c, Number of singleton or shared variants. d, Functional classes of genes with nonsynonymous mutations or indels. n = 30 genes. NSY, nonsynonymous SNVs; SYN, synonymous SNVs; Indel, small insertion/deletion variant; Inter, intergenic variant. The schematic in a was created using BioRender.
Extended Data Fig. 3 Within-host evolution of E. gallinarum in monocolonized mice.
a–c, Phylogenetic tree of liver and faecal E. gallinarum isolates sampled from mouse 1 (a), mouse 2 (b) or mice 3-5 (c). Reference-based alignments. d, Position, effects, and frequency of mutations at walK, manX, manY, lacE, ypdA, and immR in liver or faecal isolates sampled from each individual mouse. Rows represent mutations and columns represent individual mice. e, Relative abundance of E. gallinarum isolates in the faeces of bi-colonized mice. EGF1-FE4, EGmono6-LV1, or EGmono6-LV10 was co-gavaged with an equal dose of EGmono7-FE2 and samples were collected 4 weeks after co-colonization. See Supplementary Table 7 for a summary of all mutants. n = 3 mice. Representative of two independent experiments. Data represent mean ± SEM. Two-way ANOVA with Sidak’s post hoc test. f, Colony morphologies of EGmono6-LV1, EGmono6-LV10, and EGmono7-FE2 after 48-hour aerobic culture on GAM agar plates.
Extended Data Fig. 4 Reconstructed evolutionary paths of E. gallinarum within individual hosts.
Reconstructed phylogenetic histories of E. gallinarum isolates from mouse 1 (a), mouse 2 (b), mice 3–5 (c), and mice 6–8 (d). The mouse 7 is also presented in Fig. 2a. Bar plots show bacterial load recovered from the liver of each monocolonized mouse. n = 1 mouse. Circles represent sequenced isolates and squares represent hypothetical intermediate genotypes. Different colours indicate distinct genotypes. Arrows connect related genotypes and dashed lines connect genetically identical isolates. Liver and faecal populations are separated by a blue dashed line. The schematics in a-d were created using BioRender.
Extended Data Fig. 5 Transcriptomic comparisons of liver and faecal E. gallinarum isolates, and susceptibility of E. gallinarum pgdA mutant to liver clearance.
a, The minimum inhibitory concentrations (MIC) of mCRAMP against E. gallinarum isolates. b, Gating strategy used to select live macrophages and quantify CFSE signal in Fig. 3c. c–g, Volcano plots showing adjusted p-value versus fold change for 3575 genes expressed in EGmono6-LV1 (c), EGmono6-LV10 (d) as compared to EGmono7-FE2, or in EGmono6-LV1 (e), EGmono6-LV10 (f) and EGmono7-FE2 (g) isolate as compared to the ancestral strain EGF1-FE4. Horizontal lines show the p-value cut-off for significance after correction for multiple testing (adjust p-value < 0.05). Differentially expressed genes are coloured as red. n = 3 independent cultures. h, Overrepresented Pfam functional classes (EGmono6-LV10 vs. EGmono7-FE2). i, Expressions of pgdA gene in cultured E. gallinarum isolates. n = 3 independent cultures. j, Liver clearance of intravenously injected wild-type or pgdA mutant strains of E. gallinarum. Liver bacterial load 5 days post-injection. n = 4 mice. Representative of two independent experiments. Data in (i, j) represent mean ± SEM. Two-tailed Wald test with Benjamini-Hochberg correction (FDR = 0.05) (c–g), one-way ANOVA with Benjamini-Hochberg correction (FDR = 0.05) (i), unpaired two-tailed t-test (j).
Extended Data Fig. 6 Liver and faecal E. gallinarum isolates elicit distinct immune responses in the intestine and liver.
a–e, Transcriptomics of ileal epithelia and ileal tissues. a, b, Eight-hour monocolonization. Enriched pathways (a) and differentially expressed genes (b) in IECs between mice colonized by EGmono6-LV10 or EGmono7-FE2. n = 4 mice. c–e, Two-week monocolonization. c, Pathways enriched in IECs or ileal tissues induced by EGmono6-LV10 or EGmono7-FE2 colonization. d, e, Enriched pathways (d) and differentially expressed genes (e) in IECs or ileal tissues between mice colonized by EGmono6-LV1 or EGmono7-FE2. IECs (n = 4), ileal tissues (n = 3). Left and right heatmaps display distinct gene sets. f, Density of E. gallinarum in distal ileum after 8-hour or 2-week monocolonization. 8 h: EGmono6-LV10 (n = 6), EGmono7-FE2 (n = 8). 2 weeks: n = 4 mice. g, h, PAS staining (g) or CD3 immunohistochemistry staining (h) showing goblet cells (g, n = 40 villi) or IELs (h, n = 90 villi) in distal ileum of monocolonized mice 2 weeks (g) or 18 h (h) post-gavage. Scale bars, 50 μm. i, Gut permeability of monocolonized mice. MyD88-/-Trif-/-: EGmono6-LV1 (n = 6), EGmono6-LV10 and EGmono7-FE2 (n = 7). Rag1-/-: EGmono6-LV1 (n = 7), EGmono6-LV10 and EGmono7-FE2 (n = 6). j, Percentage of monocolonized mice showing liver translocation. k, Pathways enriched in livers induced by EGmono6-LV10 or EGmono7-FE2 monocolonization. Results are representative of n =2 (a-i) or 5 (j) independent experiments. Data represent mean ± SEM (f, i), violin plots (g, h) showing median (red) and quartiles (blue). Two-tailed Wald test with Benjamini-Hochberg correction (FDR = 0.05) (b, e), two-tailed Wilcoxon test (f, left panel), Kruskal-Wallis test with Benjamini-Hochberg correction (FDR = 0.05) (f, right panel; h, i), one-way ANOVA with Tukey’s post hoc test (g), paired two-tailed t-test (j). NES, normalized enrichment score.
Extended Data Fig. 7 Liver E. gallinarum isolates from SPF NZW × BXSB F1 mice exhibit similar features as experimentally evolved liver isolates from monocolonizations.
a, TEM images of cultured E. gallinarum isolates from NZW × BXSB F1 mice. Two bacterial cells of each isolate were shown. b, Minimum inhibitory concentration for mCRAMP against NZW × BXSB F1 E. gallinarum isolates. c, Liver clearance of intravenously injected NZW × BXSB F1 E. gallinarum isolates. Liver bacterial load 5 days post-injection. n = 9 mice. d, Differentially expressed genes in IECs (EGF1-LV1: n = 4, EGF1-FE4: n = 3) or ileal tissues (n = 4) between mice monocolonized with EGF1-LV or EGF1-FE4 for 2 weeks. Left and right heatmaps display distinct gene sets. e, Gut permeability of mice monocolonized with EGF1-LV1 (n = 11) or EGF1-FE4 (n = 9) for 2 weeks. f–i, Imiquimod-induced autoimmune phenotypes in mice monocolonized with EGF1-LV1 or EGF1-FE4. f, Schematic of imiquimod-induced autoimmunity. E. gallinarum translocation to the liver (g), mLNs (h) and spleen (i). n = 9 mice. Results in (a–i) are representative of two independent experiments. Data in (c, e, g–i) represent mean ± SEM. Two-tailed Mann-Whitney test (c, g–i), unpaired two-tailed t-test (e), two-tailed Wald test with Benjamini-Hochberg correction (FDR = 0.05) (d). The schematic in f was created using BioRender.
Extended Data Fig. 8 Divergent evolution of B. fragilis in monocolonized mice.
a, Schematic of experimental evolution of B. fragilis in monocolonized mice. Pie charts showing proportions of mice with liver translocation. n = 5 mice. b, Reconstructed phylogenetic histories of B. fragilis isolates sampled from two co-housed mice (BF1 and BF2). Circles represent sequenced isolates and squares represent hypothetical intermediate genotypes. Arrows connect related genotypes and dashed lines connect genetically identical isolates. SIM and faecal populations are separated by a blue dashed line. c, Liver clearance of intravenously injected B. fragilis. Representative SIM (BFmono1-LV5, BFmono2-LV8) and faecal (BFmono1-LV6) isolates. Liver bacterial load 1 or 3 dpi. n = 4 mice. Representative of two independent experiments. Data represent mean ± SEM. One-way ANOVA with Tukey’s post hoc test (c). SIM, small intestinal mucosa. dpi, days post-injection. The schematic in a was created using BioRender.
Extended Data Fig. 9 Impacts of within-host evolution of E. gallinarum on bacterial translocation and initiation of inflammation in the intestine and liver.
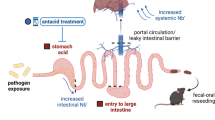
Within-host evolution of E. gallinarum facilitates divergence into independent lineages with distinct niche preferences and capacities for translocation. Faecal lineages preferentially colonize the intestinal lumen and are highly susceptible to immune recognition and clearance. By contrast, mucosa-associated lineages are adapted to colonize mucosal niches and exhibit enhanced resistance to immune recognition and clearance via remodelling of the bacterial cell wall. Functionally, the faecal lineage elicits robust immune responses at the intestinal epithelium, including enhanced mucus production and increased IEL recruitment; these responses reinforce the integrity of the gut barrier and limit bacterial translocation. By contrast, mucosally adapted E. gallinarum lineages evade initial detection at the epithelial surface, translocate across the ‘leaky’ gut barrier, exhibit increased survival after translocation, and trigger inflammation in the intestinal lamina propria and liver. The schematic was created using BioRender.
Supplementary information
Supplementary Table 1
Mutations detected in NZW × BXSB F1 mice-derived E. gallinarum isolates. This table lists mutations detected in 11 E. gallinarum isolates sampled from the livers or faeces of 18-week-old NZW × BXSB F1 hybrid mice. The EGF1-FE1 strain was used as the reference genome.
Supplementary Table 2
Mutations detected in E. gallinarum isolates evolved in monocolonized C57BL/6 mice. This table lists mutations detected in 153 E. gallinarum isolates sampled from the livers or faeces of eight monocolonized C57BL/6 mice. E. gallinarum isolates were collected after colonization for 3 months. EGF1-FE4 was the ancestral strain and was used as the reference genome.
Supplementary Table 3
Mutations detected in E. gallinarum isolates evolved in a simplified mock microbial community. This table lists mutations detected in 20 E. gallinarum isolates sampled from the livers or faeces of a gnotobiotic C57BL/6 mouse colonized by E. gallinarum and a nine-species mock community. E. gallinarum isolates were collected after colonization for 7 weeks. EGF1-FE4 was the ancestral strain and was used as the reference genome.
Supplementary Table 4
Mutations detected in E. gallinarum isolates evolved in SPF C57BL/6 mice. This table lists mutations detected in 39 E. gallinarum isolates sampled from the SIM or faeces of two cohoused SPF C57BL/6 mice. E. gallinarum-free C57BL/6 mice were purchased from Jackson Laboratory and were inoculated with EGF1-FE4 strain. E. gallinarum isolates were collected after colonization for 1.5 years. EGF1-FE4 was the ancestral strain and was used as the reference genome.
Supplementary Table 5
Mutations detected in L. reuteri isolates evolved in monocolonized C57BL/6 mice. This table lists mutations detected in 20 L. reuteri isolates sampled from the livers or faeces of a monocolonized C57BL/6 mouse. L. reuteri isolates were collected after colonization for 3 months. The draft genome of the ancestral strain was used as the reference genome.
Supplementary Table 6
Mutations detected in B. fragilis isolates evolved in monocolonized C57BL/6 mice. This table lists mutations detected in 48 B. fragilis isolates sampled from the SIM or faeces of two cohoused monocolonized C57BL/6 mouse. B. fragilis isolates were collected after colonization for 3 months. The draft genome of the ancestral strain was used as the reference genome.
Supplementary Table 7
Metadata of selected bacterial strains used in this study. This table lists the identifier, species, host of origin, duration of colonization, source location and bacterial genotype of bacterial strains assessed in this study.
Supplementary Table 8
Annotation of select mutated genes in this study. This table lists annotations of select mutated genes by RAST server v.2.0 (https://rast.nmpdr.org/rast.cgi) or Prokka v.1.12.
Source data
Rights and permissions
About this article

Cite this article
Yang, Y., Nguyen, M., Khetrapal, V. et al. Within-host evolution of a gut pathobiont facilitates liver translocation. Nature 607, 563–570 (2022). https://doi.org/10.1038/s41586-022-04949-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-04949-x
This article is cited by
-
Quantifying the adaptive landscape of commensal gut bacteria using high-resolution lineage tracking
Nature Communications (2024)
-
The gut microbiome in systemic lupus erythematosus: lessons from rheumatic fever
Nature Reviews Rheumatology (2024)
-
Contributing roles of mitochondrial dysfunction and hepatocyte apoptosis in liver diseases through oxidative stress, post-translational modifications, inflammation, and intestinal barrier dysfunction
Cellular and Molecular Life Sciences (2024)
-
Akkermansia muciniphila-induced trained immune phenotype increases bacterial intracellular survival and attenuates inflammation
Communications Biology (2024)
-
Exposing new taxonomic variation with inflammation — a murine model-specific genome database for gut microbiome researchers
Microbiome (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
