
Abstract
Birds and mammals independently evolved the highest metabolic rates among living animals1. Their metabolism generates heat that enables active thermoregulation1, shaping the ecological niches they can occupy and their adaptability to environmental change2. The metabolic performance of birds, which exceeds that of mammals, is thought to have evolved along their stem lineage3,4,5,6,7,8,9,10. However, there is no proxy that enables the direct reconstruction of metabolic rates from fossils. Here we use in situ Raman and Fourier-transform infrared spectroscopy to quantify the in vivo accumulation of metabolic lipoxidation signals in modern and fossil amniote bones. We observe no correlation between atmospheric oxygen concentrations11 and metabolic rates. Inferred ancestral states reveal that the metabolic rates consistent with endothermy evolved independently in mammals and plesiosaurs, and are ancestral to ornithodirans, with increasing rates along the avian lineage. High metabolic rates were acquired in pterosaurs, ornithischians, sauropods and theropods well before the advent of energetically costly adaptations, such as flight in birds. Although they had higher metabolic rates ancestrally, ornithischians reduced their metabolic abilities towards ectothermy. The physiological activities of such ectotherms were dependent on environmental and behavioural thermoregulation12, in contrast to the active lifestyles of endotherms1. Giant sauropods and theropods were not gigantothermic9,10, but true endotherms. Endothermy in many Late Cretaceous taxa, in addition to crown mammals and birds, suggests that attributes other than metabolism determined their fate during the terminal Cretaceous mass extinction.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others
Data availability
The Supplementary Information and Supplementary Data 1 and 2 contain all methods and source data: specimen information, method details, extracted S- and N-crosslink Raman intensities, formulae and conversions used to calculate the metabolic rates, details of regressions, data used for principal component and discriminant analyses, published body masses or body-mass estimates, correlation analyses, assessments of the prediction performance of various used regression models, assessments of potential data biases, labelled plots of all the analyses performed, data for the ancestral reconstruction. ATR FT-IR spectra are plotted in Extended Data Figs. 4, 5 and 6.
Code availability
R code for phylogeny time-scaling and ancestral state reconstruction is available at GitHub (https://github.com/IrisMenendez/Dinosaur_metabolism).
References
Grigg, G. C., Beard, L. A. & Augee, M. L. The evolution of endothermy and its diversity in mammals and birds. Physiol. Biochem. Zool. 77, 982–997 (2004).
Rolland, J. et al. The impact of endothermy on the climatic niche evolution and the distribution of vertebrate diversity. Nat. Ecol. Evol. 2, 459–464 (2018).
Bakker, R. T. Anatomical and ecological evidence of endothermy in dinosaurs. Nature 238, 81–85 (1972).
Köhler, M., Marín-Moratalla, N., Jordana, X. & Aanes, R. Seasonal bone growth and physiology in endotherms shed light on dinosaur physiology. Nature 487, 358–361 (2012).
Legendre, L. J., Guénard, G., Botha-Brink, J. & Cubo, J. Palaeohistological evidence for ancestral high metabolic rate in archosaurs. Syst. Biol. 65, 989–996 (2016).
Amiot, R. et al. Oxygen isotopes from biogenic apatites suggest widespread endothermy in Cretaceous dinosaurs. Earth Planet. Sci. Lett. 246, 41–54 (2006).
Eagle, R. A. et al. Dinosaur body temperatures determined from isotopic (13C-18O) ordering in fossil biominerals. Science 333, 443–445 (2011).
Rezende, E. L., Bacigalupe, L. D., Nespolo, R. F. & Bozinovic, F. Shrinking dinosaurs and the evolution of endothermy in birds. Sci. Adv. 6, eaaw4486 (2020).
Paladino, F. V., O’Connor, M. P. & Spotila, J. R. Metabolism of leatherback turtles, gigantothermy, and thermoregulation of dinosaurs. Nature 344, 858–860 (1990).
Grady, J. M., Enquist, B. J., Dettweiler-Robinson, E., Wright, N. A. & Smith, F. A. Evidence for mesothermy in dinosaurs. Science 344, 1268–1272 (2014).
Berner, R. A. GEOCARBSULF: a combined model for Phanerozoic atmospheric O2 and CO2. Geochim. Cosmochim. Acta 70, 5653–5664 (2006).
Weiss, B. & Laties, V. G. Behavioral thermoregulation. Science 133, 1338–1344 (1961).
Walter, I. & Seebacher, F. Endothermy in birds: underlying molecular mechanisms. J. Exp. Biol. 212, 2328–2336 (2009).
Anderson, K. J. & Jetz, W. The broad‐scale ecology of energy expenditure of endotherms. Ecol. Lett. 8, 310–318 (2005).
Buckley, L. B., Hurlbert, A. H. & Jetz, W. Broad‐scale ecological implications of ectothermy and endothermy in changing environments. Glob. Ecol. Biogeogr. 21, 873–885 (2012).
Ramos, E. K. S., Freitas, L. & Nery, M. F. The role of selection in the evolution of marine turtles mitogenomes. Sci Rep. 10, 16953 (2020).
Christian, K. A. & Conley, K. E. Activity and resting metabolism of varanid lizards compared with typical lizards. Aust. J. Zool. 42, 185–193 (1994).
Tattersall, G. J. et al. Seasonal reproductive endothermy in tegu lizards. Sci. Adv. 2, e1500951 (2016).
Pamplona, R. Advanced lipoxidation end-products. Chem. Biol. Interact. 192, 14–20 (2011).
Miyata, T., Kurokawa, K. & van Ypersele de Strihou, C. Advanced glycation and lipoxidation end products: role of reactive carbonyl compounds generated during carbohydrate and lipid metabolism. J. Am. Soc. Nephrol. 11, 1744–1752 (2000).
Vistoli, G. et al. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): an overview of their mechanisms of formation. Free Radic. Res. 47, 3–27 (2013).
Wiemann, J. et al. Fossilization transforms vertebrate hard tissue proteins into N-heterocyclic polymers. Nat. Commun. 9, 4741 (2018).
Wiemann, J., Crawford, J. M. & Briggs, D. E. G. Phylogenetic and physiological signals in metazoan fossil biomolecules. Sci. Adv. 6, eaba6883 (2020).
Wiemann, J. & Briggs, D. E. G. Raman spectroscopy is a powerful tool in molecular paleobiology: an analytical response to Alleon et al. (https://doi.org/10.1002/bies.202000295). Bioessays 44, 2100070 (2022).
Barrick, R. E. & Showers, W. J. Thermophysiology of Tyrannosaurus rex: evidence from oxygen isotopes. Science 265, 222–224 (1994).
Capellini, I., Venditti, C. & Barton, R. A. Phylogeny and metabolic scaling in mammals. Ecology 91, 2783–2793 (2010).
Benson, R. B. J. et al. Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage. PLoS Biol. 12, e1001853 (2014).
Erickson, G. M. On dinosaur growth. Annu. Rev. Earth Planet. Sci. 42, 675–697 (2014).
Wiemann, J., Yang, T.-R. & Norell, M. A. Dinosaur egg colour had a single evolutionary origin. Nature 563, 555–558 (2018).
Norell, M. A. et al. The first dinosaur egg was soft. Nature 583, 406–410 (2020).
Schaefer, B. et al. Microbial life in the nascent Chicxulub crater. Geology 48, 328–332 (2020).
Sepúlveda, J., Wendler, J. E., Summons, R. E. & Hinrichs, K. U. Rapid resurgence of marine productivity after the Cretaceous–Paleogene mass extinction. Science 326, 129–132 (2009).
Hull, P. M. et al. On impact and volcanism across the Cretaceous–Paleogene boundary. Science 367, 266–272 (2020).
Plet, C. et al. Palaeobiology of red and white blood cell-like structures, collagen and cholesterol in an ichthyosaur bone. Sci Rep. 7, 13776 (2017).
Barthel, H. J., Fougerouse, D., Geisler, T. & Rust, J. Fluoridation of a lizard bone embedded in Dominican amber suggests open-system behavior. PLoS ONE 15, e0228843 (2020).
Kourkoumelis, N., Polymeros, A. & Tzaphlidou, M. Background estimation of biomedical Raman spectra using a geometric approach. J. Spectrosc. 27, 311–317 (2012).
Acknowledgements
V. Rhue, K. Zyskowski and G. Watkins-Colwell facilitated specimen selection; T. Wu and M. Ghosh provided guidance for ATR FT-IR spectroscopy at the Yale West Campus Analytical Core. J.W. was supported by a Dissertation Improvement Grant from the Yale Institute for Biospheric Studies and a Graduate Student Research Grant from the Geological Society of America. I.M. was funded by a predoctoral grant from the Universidad Complutense de Madrid (CT27/16-CT28/16) and partially supported by project PGC2018-094955-A-I00 funded by the Spanish Ministerio de Ciencia, Innovación y Universidades. P.M.H. was funded by a Sloan Research Fellowship.
Author information
Authors and Affiliations
Contributions
J.W., J.M.C. and D.E.G.B. designed the project. M.F., J.A.G. and M.A.N. helped with specimen selection and sample materials preparation. J.W. designed the analytical strategy, performed Raman and FT-IR spectroscopy, processed and analysed the data and prepared the figures. J.W., P.M.H. and I.M. discussed statistical analyses. I.M. ran the time-calibrated ancestral state reconstruction. All authors discussed the data. J.W. and D.E.G.B. wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Kliti Grice, Marcin Kozanecki, Nick Stone and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Multivariate spectral analyses of biological signals in fossil amniote hard tissues.
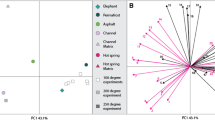
a ChemoSpace Principal Component Analysis for avascular and vascular amniote hard tissues (n = 51 data points, each based on n = 1 biological replicate). Each data point (circle) represents relative intensities at 24 band positions (Supplementary Data 1, tab 12) for in situ Raman spectra ranging from 500–1800 cm−1 Raman shift (averaged over n = 10 technical replicates which were mean-averaged). The ChemoSpace separates a grouping of avascular, acellular fossil eggshell tissues (grey convex hull) from vascular, cellular fossil bones and teeth (turquoise/orange convex hull). Eigenvectors for S- (blue) and N-crosslinks (orange) suggest that the separation of avascular and vascular fossil tissues follows primarily the trajectory of the S-crosslink vector, which represents metabolic and diagenetic lipoxidation products. PC 1 = 43%, PC 2 = 36%. b Discriminant analysis of metabolic signals in fossil amniote long bones (n = 30 subsampled from a; Supplementary Data 1, tab 12). Taxa with higher (orange circles) and lower metabolic rates (blue circles) separate on Discriminant Axis 2 and are contained within the turquoise/orange convex hull (metabolic assignments rely on published literature, see Supplementary Information section 2b). Black outlines = known metabolism (phylogenetic inference), red outlines = unknown metabolism. Specimens with evidence for elevated metabolic rates follow discriminant vectors for increased concentrations of metabolic S- (blue vector) and N-crosslinks (orange vector). Metabolic rates and resulting thermal strategies of modern and fossil amniote femora analysed in Fig. 2 were calculated solely from spectral MCin vivo values (Supplementary Data 1, tabs 1, 2) and their placement relative to the endothermy confidence interval in Fig. 1d. Thermal strategies were not inferred from the ChemoSpace plots.
Extended Data Fig. 2 Intraskeletal variation of MCin vivo values (spectral crosslink-to-peptide ratios).
This box-and-whiskers plot with integrated outlier analysis shows the range of MCin vivo values calculated from different skeletal elements of the endothermic mammal Chironectes (pink) and the ectothermic lizard Iguana (blue). Intraskeletal variation of metabolic signals recorded in ALEs is substantially larger in Chironectes than in Iguana. Metabolic rates in this study were calculated from modern (adult, except for the juvenile Gavialis) and fossil femora to ensure comparability. Analyses of intraskeletal signal variation suggest that ectotherm lizard femora, vertebrae, skull bones (frontals) and ribs yield comparable metabolic signals, whereas metabolic rate calculations for endotherms require individual calibrations for each type of bone used. Whisker length: one sigma, standard error (s.e.m.); quartile method: interpolation; this analysis was performed in PAST (see Supplementary Data 1, tab 8).
Extended Data Fig. 3 Modern and fossil crosslink-to-peptide (MCin vivo) ratios versus corresponding amniote body masses plotted from raw data.
MCin vivo ratios were directly calculated from baselined and normalized spectra of modern and fossil amniote samples (total n = 53 femora, n = 1 humerus, n = 1 vertebra, 10 s exposure time, 10 technical replicates mean-averaged; see Supplementary Information ‘Method details’). MCin vivo values of fossil samples (black outlines) are consistently higher than those of modern samples (red outlines), due to the formation of additional S- and N-crosslinks during diagenesis (Fig. 1c). MCin vivo data for modern taxa separate known endotherms with elevated metabolic rates (orange dots) from ectotherms with lower metabolic rates (blue dots), a pattern mirrored in the fossil MCin vivo values. Fossil thermal strategies were assigned based on the literature for crown lineage taxa, or inferred from the endothermy assessment (Fig. 1d). The sample separation relies exclusively on the spectral MCin vivo data, and thermal strategies are only highlighted for visual purposes. Both modern and fossil varanids (grey data points) plot among endotherms with elevated metabolic rates.
Extended Data Fig. 4
Comparison of the molecular signal coverage in in situ Raman spectroscopic and Attenuated-reflectance Fourier-Transform Infrared spectra (ATR FT-IR) for a Allosaurus, b Furcifer, c Chironectes, and d Puma. Raman spectra analyzed for metabolic rates (solid blue lines) and ATR FT-IR spectra (solid orange lines) obtained for representative modern (n = 3) and fossil (n = 1) bones are compared to published reference spectra35,36 (dotted lines). The plots show on the top half normalized spectral intensities (arbitrary units = a.u.), and on the bottom half normalized reflectance. The statistically analyzed organic fingerprint region (500–1800 cm−1) is shown in the large plots. The smaller corner plots show signals beyond the fingerprint region (red squares) illustrated in the large plots (full range: 500–3000 cm−1). In situ Raman spectra were obtained with n = 10 mean-averaged technical replicates, and ATR FT-IR spectra were acquired with n = 32 mean-averaged technical replicates. Arrows highlight significant peaks and bands in the spectra produced by molecular heterogeneities; blue: Raman spectroscopy, orange: ATR FT-IR.
Extended Data Fig. 5
Comparison of S-crosslinks (arrows; other signals may contribute to the total peak intensity) in the organic fingerprint region (500–1800 cm−1) of normalized Attenuated-reflectance Fourier-Transform Infrared (ATR FT-IR) spectra obtained for femora of two endotherms (orange: Chironectes, red: Puma) and an ectotherm (blue: Furcifer). Each ATR FT-IR spectrum was acquired with n = 32 mean-averaged technical replicates.
Extended Data Fig. 6 Net enrichment plot identifying Attenuated-reflectance Fourier-Transform Infrared (ATR FT-IR) spectral signatures that represent reaction educts (blue) and products (orange) of biomolecule fossilization in amniote bone.
Normalized spectra of modern (n = 3 biological replicates, n = 32 technical replicates each, all mean-averaged) and fossil (n = 1 biological sample, n = 32 technical replicates, all mean-averaged) bone were subtracted to identify the net differences over the organic fingerprint region (500–1800 cm−1). All major bands are labelled. This net enrichment plot recovers fossilization reactions identical to those revealed in previously published in situ Raman net enrichment plots22,23, thereby corroborating the impact of organic diagenesis on the preservation potential and additional formation of S-crosslinks in bone organic matter.
Extended Data Fig. 7 Linear regression (dotted line) of MCin vivo values and published metabolic rates in (n=25) extant amniote bones.
Data points are coloured according to phylogenetic affinity (see Supplementary Data 1, tab 2). The orange area represents the 95%-confidence interval for the regression function. Statistics: Pearson’s r = 0.82, r2 = 0.67, rs = 0.74. Spectra used to calculate MCin vivo values were acquired for each n = 1 biological replicate with n = 10 mean-averaged technical replicates.
Extended Data Fig. 8 Ranges and outlier analysis of calculated metabolic rates in key clades.
This box-and-whiskers plot with an integrated outlier analysis highlights the variation of spectral metabolic rates obtained for non-avian ornithodirans (grey), crown mammals (pink), crown birds (orange), and crown lepidosaurs (blue). The upper limits of non-avian ornithodiran metabolism exceed the limits of mammalian endothermy, providing further support for an ornithodiran origin of avian metabolism. Ectothermic dinosaur metabolic rates align with those of crown lepidosaurs. Whisker length: one sigma, standard error (s.e.m.); quartile method: interpolation; this analysis was performed in PAST (see Supplementary Data 1, tab 7).
Supplementary information
Supplementary Information
Details regarding specimen selection, organic matter endogeneity, a comparison of Raman metabolic rates and palaeohistological metabolic inferences, and the Supplementary Methods, Supplementary References, Supplementary Tables 1–4 and Supplementary Figs 1, 2.
Supplementary Data 1
Supplementary Data including: Fossil taxa: specimen information and spectral data; Extant taxa: specimen information and spectral data; source data for Extended Data Fig. 7; Calibration: calibration function and statistics; source data for Fig. 1b; Allometry: mass-specific metabolic rates for modern and fossil taxa; source data for Fig. 1d; Modern and fossil metabolic rates (MRs): source data for Fig. 1d, e and Extended Data Fig. 3; Endothermy confidence interval, source data for Fig. 1d; MR ranges: intra-clade variation of metabolic rates; source data for Extended Data Fig. 8; Intraskeletal variation: endotherm versus ectotherm, source data for Extended Data Fig. 2; Correlation analyses: behavioural and environmental correlates; Oxygen (O2) through time: correlation analysis; Summary of statistical results: Pearson’s r, r2, Spearman ranks; ChemoSpace data, variance–covariance matrix; source data for Extended Data Fig. 1; Sediments, sample information; Ancestral state reconstruction (ACS) codings: alternative trees; source data for Fig. 2; Additional band assignments: signals beyond the organic fingerprint region in Raman and IR.
Supplementary Data 2
Node and tip IDs and metabolic rates (MRs): explanation and source data for Figs. 1 and 2. The source code used to perform maximum-likelihood ancestral state reconstructions is available at Github (https://github.com/IrisMenendez/Dinosaur_metabolism).
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wiemann, J., Menéndez, I., Crawford, J.M. et al. Fossil biomolecules reveal an avian metabolism in the ancestral dinosaur. Nature 606, 522–526 (2022). https://doi.org/10.1038/s41586-022-04770-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-04770-6
This article is cited by
-
Global latitudinal gradients and the evolution of body size in dinosaurs and mammals
Nature Communications (2024)
-
Molecular fingerprints resolve affinities of Rhynie chert organic fossils
Nature Communications (2023)
-
Reply to: Amniote metabolism and the evolution of endothermy
Nature (2023)
-
Amniote metabolism and the evolution of endothermy
Nature (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
