
Abstract
In the eighteenth century, Daniel Bernoulli, Adam Smith and Jeremy Bentham proposed that economic choices rely on the computation and comparison of subjective values1. This hypothesis continues to inform modern economic theory2 and research in behavioural economics3, but behavioural measures are ultimately not sufficient to verify the proposal4. Consistent with the hypothesis, when agents make choices, neurons in the orbitofrontal cortex (OFC) encode the subjective value of offered and chosen goods5. Value-encoding cells integrate multiple dimensions6,7,8,9, variability in the activity of each cell group correlates with variability in choices10,11 and the population dynamics suggests the formation of a decision12. However, it is unclear whether these neural processes are causally related to choices. More generally, the evidence linking economic choices to value signals in the brain13,14,15 remains correlational16. Here we show that neuronal activity in the OFC is causal to economic choices. We conducted two experiments using electrical stimulation in rhesus monkeys (Macaca mulatta). Low-current stimulation increased the subjective value of individual offers and thus predictably biased choices. Conversely, high-current stimulation disrupted both the computation and the comparison of subjective values, and thus increased choice variability. These results demonstrate a causal chain linking subjective values encoded in OFC to valuation and choice.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


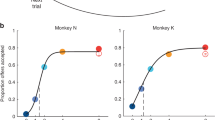
Data availability
The complete dataset is available at https://github.com/PadoaSchioppaLab/2020_Ballesta_etal_Nature. Source data are provided with this paper.
Code availability
The MATLAB code used for the analysis is available at https://github.com/PadoaSchioppaLab/2020_Ballesta_etal_Nature.
References
Niehans, J. A History of Economic Theory: Classic Contributions, 1720–1980 (Johns Hopkins Univ. Press, 1990).
Kreps, D. M. A Course in Microeconomic Theory (Princeton Univ. Press, 1990).
Kahneman, D. & Tversky, A. (eds) Choices, Values and Frames (Russell Sage Foundation–Cambridge Univ. Press, 2000).
Camerer, C. F., Loewenstein, G. & Prelec, D. Neuroeconomics: how neuroscience can inform economics. J. Econ. Lit. 43, 9–64 (2005).
Padoa-Schioppa, C. & Assad, J. A. Neurons in the orbitofrontal cortex encode economic value. Nature 441, 223–226 (2006).
Roesch, M. R. & Olson, C. R. Neuronal activity in primate orbitofrontal cortex reflects the value of time. J. Neurophysiol. 94, 2457–2471 (2005).
Kennerley, S. W., Dahmubed, A. F., Lara, A. H. & Wallis, J. D. Neurons in the frontal lobe encode the value of multiple decision variables. J. Cogn. Neurosci. 21, 1162–1178 (2009).
Pastor-Bernier, A., Stasiak, A. & Schultz, W. Orbitofrontal signals for two-component choice options comply with indifference curves of Revealed Preference Theory. Nat. Commun. 10, 4885 (2019).
Hare, T. A., O’Doherty, J., Camerer, C. F., Schultz, W. & Rangel, A. Dissociating the role of the orbitofrontal cortex and the striatum in the computation of goal values and prediction errors. J. Neurosci. 28, 5623–5630 (2008).
Padoa-Schioppa, C. Neuronal origins of choice variability in economic decisions. Neuron 80, 1322–1336 (2013).
Conen, K. E. & Padoa-Schioppa, C. Neuronal variability in orbitofrontal cortex during economic decisions. J. Neurophysiol. 114, 1367–1381 (2015).
Rich, E. L. & Wallis, J. D. Decoding subjective decisions from orbitofrontal cortex. Nat. Neurosci. 19, 973–980 (2016).
Roesch, M. R. & Olson, C. R. Neuronal activity related to anticipated reward in frontal cortex: does it represent value or reflect motivation? Ann. NY Acad. Sci. 1121, 431–446 (2007).
Schultz, W. Neuronal reward and decision signals: from theories to data. Physiol. Rev. 95, 853–951 (2015).
Bartra, O., McGuire, J. T. & Kable, J. W. The valuation system: a coordinate-based meta-analysis of BOLD fMRI experiments examining neural correlates of subjective value. Neuroimage 76, 412–427 (2013).
Stalnaker, T. A., Cooch, N. K. & Schoenbaum, G. What the orbitofrontal cortex does not do. Nat. Neurosci. 18, 620–627 (2015).
Cohen, M. R. & Newsome, W. T. What electrical microstimulation has revealed about the neural basis of cognition. Curr. Opin. Neurobiol. 14, 169–177 (2004).
Clark, K. L., Armstrong, K. M. & Moore, T. Probing neural circuitry and function with electrical microstimulation. Proc. R. Soc. Lond. B 278, 1121–1130 (2011).
Salzman, C. D., Britten, K. H. & Newsome, W. T. Cortical microstimulation influences perceptual judgements of motion direction. Nature 346, 174–177 (1990).
Murasugi, C. M., Salzman, C. D. & Newsome, W. T. Microstimulation in visual area MT: effects of varying pulse amplitude and frequency. J. Neurosci. 13, 1719–1729 (1993).
Kuwabara, M., Kang, N., Holy, T. E. & Padoa-Schioppa, C. Neural mechanisms of economic choices in mice. eLife 9, e49669 (2020).
Wolff, S. B. & Ölveczky, B. P. The promise and perils of causal circuit manipulations. Curr. Opin. Neurobiol. 49, 84–94 (2018).
Padoa-Schioppa, C. Range-adapting representation of economic value in the orbitofrontal cortex. J. Neurosci. 29, 14004–14014 (2009).
Kobayashi, S., Pinto de Carvalho, O. & Schultz, W. Adaptation of reward sensitivity in orbitofrontal neurons. J. Neurosci. 30, 534–544 (2010).
Rustichini, A., Conen, K. E., Cai, X. & Padoa-Schioppa, C. Optimal coding and neuronal adaptation in economic decisions. Nat. Commun. 8, 1208 (2017).
Histed, M. H., Bonin, V. & Reid, R. C. Direct activation of sparse, distributed populations of cortical neurons by electrical microstimulation. Neuron 63, 508–522 (2009).
Ballesta, S. & Padoa-Schioppa, C. Economic decisions through circuit inhibition. Curr. Biol. 29, 3814–3824 (2019).
Conen, K. E. & Padoa-Schioppa, C. Partial adaptation to the value range in the macaque orbitofrontal cortex. J. Neurosci. 39, 3498–3513 (2019).
Cisek, P. Making decisions through a distributed consensus. Curr. Opin. Neurobiol. 22, 927–936 (2012).
Padoa-Schioppa, C. & Conen, K. E. Orbitofrontal cortex: a neural circuit for economic decisions. Neuron 96, 736–754 (2017).
Rustichini, A. & Padoa-Schioppa, C. A neuro-computational model of economic decisions. J. Neurophysiol. 114, 1382–1398 (2015).
Wong, K. F. & Wang, X. J. A recurrent network mechanism of time integration in perceptual decisions. J. Neurosci. 26, 1314–1328 (2006).
Merrill, D. R., Bikson, M. & Jefferys, J. G. Electrical stimulation of excitable tissue: design of efficacious and safe protocols. J. Neurosci. Methods 141, 171–198 (2005).
Kim, S. et al. Behavioral assessment of sensitivity to intracortical microstimulation of primate somatosensory cortex. Proc. Natl Acad. Sci. USA 112, 15202–15207 (2015).
Lee, S. W., Eddington, D. K. & Fried, S. I. Responses to pulsatile subretinal electric stimulation: effects of amplitude and duration. J. Neurophysiol. 109, 1954–1968 (2013).
Hussin, A. T., Boychuk, J. A., Brown, A. R., Pittman, Q. J. & Teskey, G. C. Intracortical microstimulation (ICMS) activates motor cortex layer 5 pyramidal neurons mainly transsynaptically. Brain Stimul. 8, 742–750 (2015).
Tolias, A. S. et al. Mapping cortical activity elicited with electrical microstimulation using FMRI in the macaque. Neuron 48, 901–911 (2005).
Stoney, S. D. Jr, Thompson, W. D. & Asanuma, H. Excitation of pyramidal tract cells by intracortical microstimulation: effective extent of stimulating current. J. Neurophysiol. 31, 659–669 (1968).
Arsiero, M., Lüscher, H. R., Lundstrom, B. N. & Giugliano, M. The impact of input fluctuations on the frequency-current relationships of layer 5 pyramidal neurons in the rat medial prefrontal cortex. J. Neurosci. 27, 3274–3284 (2007).
La Camera, G., Giugliano, M., Senn, W. & Fusi, S. The response of cortical neurons to in vivo-like input current: theory and experiment. I. Noisy inputs with stationary statistics. Biol. Cybern. 99, 279–301 (2008).
Ethier, C., Brizzi, L., Darling, W. G. & Capaday, C. Linear summation of cat motor cortex outputs. J. Neurosci. 26, 5574–5581 (2006).
Griffin, D. M., Hudson, H. M., Belhaj-Saïf, A. & Cheney, P. D. Hijacking cortical motor output with repetitive microstimulation. J. Neurosci. 31, 13088–13096 (2011).
Van Acker, G. M. III et al. Effective intracortical microstimulation parameters applied to primary motor cortex for evoking forelimb movements to stable spatial end points. J. Neurophysiol. 110, 1180–1189 (2013).
Acknowledgements
We thank H. Schoknecht for help with animal training and J. Assad, E. Bromberg-Martin, E. Fehr, D. Freedman, I. Monosov and L. Snyder for comments on the manuscript. This research was supported by the National Institutes of Health (grants number R01-DA032758 and R01-MH104494 to C.P.-S. and grant number F31-MH107111 to K.E.C.) and by the McDonnell Center for Systems Neuroscience (pre-doctoral fellowship to W.S.).
Author information
Authors and Affiliations
Contributions
S.B. and W.S. collected and analysed the data; K.E.C. designed experiment 2; and C.P.-S. supervised the project and wrote the manuscript. All of the authors edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Tianming Yang and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Experiment 2, control for choice frequency.
We noticed that across sessions the difference in value range (ΔVA–ΔVB) was correlated with the fraction of trials in which the animal chose juice A (% A choice) and with the relative value (ρ). In principle, these correlations could represent confounding factors. Indeed, 50 μA stimulation could partly disrupt the valuation process. As a result, the animal might respond by defaulting to the juice type most frequently chosen in that session, or to the preferred juice type. If so, the range-dependent bias would be akin to the order bias (experiment 1), in the sense that it would result from functional disruption as opposed to facilitation. To address this concern, we identified a subset of sessions for which choices between the two juices were split almost evenly. In this subset of sessions, the difference in value range and the fraction of A choices were not correlated. We reasoned that if the range-dependent bias observed for the whole dataset was driven by a default to the most frequently chosen option, the bias should disappear when the analysis was restricted to this subset of sessions. However, this was not the case. In fact, the range-dependent bias measured for the selected subset was larger than that measured for the entire population. We concluded that range-dependent biases did not reflect simple heuristics. a, Correlation between the difference in value range and the fraction of A choices. Each data point represents one session. Considering the entire dataset (black data points, N = 96 sessions), the two measures were significantly correlated (r ≥ 0.71, P < 10−15, Pearson and Spearman correlation tests). We defined a small ellipse centred on coordinates [0, 50] (axes = [9, 14]). The ellipse identified a subset of data (pink data points, N = 31 sessions) for which the difference in value range and the fraction of A choices were not correlated (P ≥ 0.69, Pearson and Spearman correlation tests). b, Correlation between the difference in value range and the relative value. Considering the entire dataset, the two measures were significantly correlated (r ≥ 0.33, P ≤ 0.001, Pearson and Spearman correlation tests). However, when the analysis was restricted to the subset of sessions identified in a (pink data points), the correlation changed sign. c, Range-dependent bias, same data as in Fig. 4c, d. Considering the entire dataset, the change in relative value was significantly correlated with the difference in value range (r ≥ 0.34, P ≤ 0.0007, Pearson and Spearman correlation tests). The correlation did not dissipate when the analysis was restricted to the subset of sessions identified in a (pink data points; r ≥ 0.45, P ≤ 0.01, Pearson and Spearman correlation tests). In this figure, data from the two animals are combined. Grey and pink lines in the three panels were obtained from Deming regressions.
Extended Data Fig. 2 Experiment 2, results obtained in paired sessions.
In N = 33 instances, we ran two back-to-back sessions offering the same two juices and leaving the electrode in place, but changing the quantity ranges such that ΔVA–ΔVB would differ. a, Example of paired sessions. b, Population analysis. Each pair of sessions in the scatter plot is connected by a line, of which we computed the slope. Data points filled in green correspond to sessions in a. Data from the two monkeys are pooled. Across the population, slopes were typically >0 (P = 0.007, two-tailed Wilcoxon signed-rank test). Hence, range-dependent biases were not dictated by the juice pair or by the location of the electrode within OFC.
Extended Data Fig. 3 Experiment 2, analysis of response times (RTs).
a, Example session 1. Each data point represents one trial type and the two lines were obtained from linear regressions. Under normal conditions (stimOFF, black), RTs decreased as a function of the chosen value (x axis). Electrical stimulation (stimON, red) generally reduced RTs. Linear fits reveal that lower RTs were due to a lower intercept, as opposed to a steeper (that is, more negative) slope. b, c, Population analysis, monkey D (N = 35). For each session, we regressed RTs onto the chosen value, separately for stimOFF and stimON trials. We then compared the intercepts and the slopes at the population level. The picture emerging from a was confirmed for the population. In b (intercept), each data point represents one session. The population is significantly displaced below the identity line (P = 0.018, two-tailed Wilcoxon test). In c (slope), it can be noticed that the slope under stimulation was shallower (less negative), probably due to a floor effect. Filled data points correspond to the session shown in a. d, Example session 2. Same format as in a. e, f, Population analysis, monkey G (N = 61). Same format as in b, c. Electrical stimulation significantly lowered the intercept but did not significantly alter the slope. Filled data points correspond to the session shown in d. In b, c, e, f, values indicated in the insert refer to the difference between the stimON measure and the stimOFF measure, averaged across the population. All P values are from two-tailed Wilcoxon tests, and t-tests provided very similar results.
Extended Data Fig. 4 Experiment 1, range-dependent choice biases.
a–c, Results obtained when electric current was delivered at 25 μA, 50 μA and ≥ 100 μA. In each panel, x and y axes represent the difference in value range (in uB) and the difference in relative value, respectively. Each data point represents one session. Sessions from the two animals and with different stimulation times (offer 1 or offer 2) were pooled. Grey lines were obtained from linear regressions. Each panel indicates the P values obtained from Pearson and Spearman correlation tests. In essence, the choice bias imposed by the stimulation (δρ) was correlated with the difference in value ranges (ΔVA–ΔVB) at low current (25 μA; weakly) and intermediate current (50 μA), but not at high current (≥100 μA).
Extended Data Fig. 5 Stimulation in experiment 2 did not systematically alter the sigmoid steepness.
For this analysis, the two groups of trials (stimOFF, stimON) were examined separately (see Methods). The two axes represent the sigmoid steepness in the two conditions. Sessions from the two animals were pooled (N = 95, 2 outliers removed), and each data point represents one session. The grey ellipse represents the 90% confidence interval. The P value is from a Wilcoxon test and similar results were obtained with a t-test.
Extended Data Fig. 6 Experiment 1, interpretation of the order bias.
a, Decelerating response function. The black line represents an ideal response function, which relates the number of spikes emitted by a cell in a given time window (y axis) to the synaptic current entering the cell (x axis). In the condition highlighted in yellow, IO is the synaptic current due to the offer on the monitor, r is the corresponding response, IS is the synaptic current due to the stimulation, and δr is the corresponding increase in the number of spikes. The condition highlighted in blue is similar, except that IO is larger (IO,blue > IO,yellow). Because the response function is decelerating, δr in the blue condition is smaller (δrblue < δryellow). In experiment 1, only one good was presented at the time. Neurons associated with that good were naturally more active (higher IO) than neurons associated with the other good. Thus deceleration in the response function induced a bias favouring the good not offered during the stimulation (order bias). For given IO,yellow and IO,blue, the difference δryellow – δrblue increases with IS. Hence, higher stimulation currents induced larger order biases. b, Concurrent presence of order bias and range-dependent bias. The cartoon illustrates an ideal session in experiment 1. We assume that under normal conditions there is no order bias (stimOFF, continuous lines). Thus the two sigmoids for AB trials and BA trials coincide. We also assume that stimulation is delivered during offer 1, and that ΔVA–ΔVB > 0. The order bias separates the two sigmoids such that under stimulation the sigmoid for AB trials is on the left of that for BA trials (stimON, dashed lines). The range-dependent bias imposes a shift on the total sigmoid, including both AB and BA trials (not shown), which moves to the right compared to normal conditions. The two choice biases are complementary and independent.
Supplementary information
Rights and permissions
About this article

Cite this article
Ballesta, S., Shi, W., Conen, K.E. et al. Values encoded in orbitofrontal cortex are causally related to economic choices. Nature 588, 450–453 (2020). https://doi.org/10.1038/s41586-020-2880-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2880-x
This article is cited by
-
A neural mechanism for conserved value computations integrating information and rewards
Nature Neuroscience (2024)
-
Curiosity: primate neural circuits for novelty and information seeking
Nature Reviews Neuroscience (2024)
-
Task state representations in vmPFC mediate relevant and irrelevant value signals and their behavioral influence
Nature Communications (2023)
-
Value dynamics affect choice preparation during decision-making
Nature Neuroscience (2023)
-
Orbitofrontal cortex control of striatum leads economic decision-making
Nature Neuroscience (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
