
Abstract
Persistently depolarizing sodium (Na+) leak currents enhance electrical excitability1,2. The ion channel responsible for the major background Na+ conductance in neurons is the Na+ leak channel, non-selective (NALCN)3,4. NALCN-mediated currents regulate neuronal excitability linked to respiration, locomotion and circadian rhythm4,5,6,7,8,9,10. NALCN activity is under tight regulation11,12,13,14 and mutations in NALCN cause severe neurological disorders and early death15,16. NALCN is an orphan channel in humans, and fundamental aspects of channel assembly, gating, ion selectivity and pharmacology remain obscure. Here we investigate this essential leak channel and determined the structure of NALCN in complex with a distinct auxiliary subunit, family with sequence similarity 155 member A (FAM155A). FAM155A forms an extracellular dome that shields the ion-selectivity filter from neurotoxin attack. The pharmacology of NALCN is further delineated by a walled-off central cavity with occluded lateral pore fenestrations. Unusual voltage-sensor domains with asymmetric linkages to the pore suggest mechanisms by which NALCN activity is modulated. We found a tightly closed pore gate in NALCN where the majority of missense patient mutations cause gain-of-function phenotypes that cluster around the S6 gate and distinctive π-bulges. Our findings provide a framework to further study the physiology of NALCN and a foundation for discovery of treatments for NALCN channelopathies and other electrical disorders.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
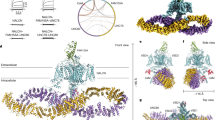
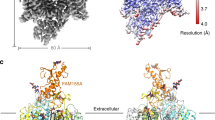
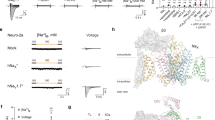
Data availability
The NALCN–FAM155A coordinates are deposited at the PDB under accession number 6XIW. Cryo-EM data are deposited in the Electron Microscopy Data Bank under accession number EMD-22203. All other data are included in the paper and the supplementary information files. Source data are provided with this paper.
References
Hodgkin, A. L. & Katz, B. The effect of sodium ions on the electrical activity of giant axon of the squid. J. Physiol. 108, 37–77 (1949).
Khaliq, Z. M. & Bean, B. P. Pacemaking in dopaminergic ventral tegmental area neurons: depolarizing drive from background and voltage-dependent sodium conductances. J. Neurosci. 30, 7401–7413 (2010).
Lee, J. H., Cribbs, L. L. & Perez-Reyes, E. Cloning of a novel four repeat protein related to voltage-gated sodium and calcium channels. FEBS Lett. 445, 231–236 (1999).
Lu, B. et al. The neuronal channel NALCN contributes resting sodium permeability and is required for normal respiratory rhythm. Cell 129, 371–383 (2007).
Lutas, A., Lahmann, C., Soumillon, M. & Yellen, G. The leak channel NALCN controls tonic firing and glycolytic sensitivity of substantia nigra pars reticulata neurons. eLife 5, e15271 (2016).
Shi, Y. et al. NALCN is a “leak” sodium channel that regulates excitability of brainstem chemosensory neurons and breathing. J. Neurosci. 36, 8174–8187 (2016).
Jospin, M. et al. UNC-80 and the NCA ion channels contribute to endocytosis defects in synaptojanin mutants. Curr. Biol. 17, 1595–1600 (2007).
Xie, L. et al. NLF-1 delivers a sodium leak channel to regulate neuronal excitability and modulate rhythmic locomotion. Neuron 77, 1069–1082 (2013).
Lear, B. C. et al. The ion channel narrow abdomen is critical for neural output of the Drosophila circadian pacemaker. Neuron 48, 965–976 (2005).
Flourakis, M. et al. A conserved bicycle model for circadian clock control of membrane excitability. Cell 162, 836–848 (2015).
Lu, B. et al. Peptide neurotransmitters activate a cation channel complex of NALCN and UNC-80. Nature 457, 741–744 (2009).
Swayne, L. A. et al. The NALCN ion channel is activated by M3 muscarinic receptors in a pancreatic β-cell line. EMBO Rep. 10, 873–880 (2009).
Lu, B. et al. Extracellular calcium controls background current and neuronal excitability via an UNC79–UNC80–NALCN cation channel complex. Neuron 68, 488–499 (2010).
Philippart, F. & Khaliq, Z. M. Gi/o protein-coupled receptors in dopamine neurons inhibit the sodium leak channel NALCN. eLife 7, e40984 (2018).
Bend, E. G. et al. NALCN channelopathies: distinguishing gain-of-function and loss-of-function mutations. Neurology 87, 1131–1139 (2016).
Bramswig, N. C. et al. Genetic variants in components of the NALCN–UNC80–UNC79 ion channel complex cause a broad clinical phenotype (NALCN channelopathies). Hum. Genet. 137, 753–768 (2018).
Stephens, R. F., Guan, W., Zhorov, B. S. & Spafford, J. D. Selectivity filters and cysteine-rich extracellular loops in voltage-gated sodium, calcium, and NALCN channels. Front. Physiol. 6, 153 (2015).
Chua, H. C., Wulf, M., Weidling, C., Rasmussen, L. P. & Pless, S. A. The NALCN channel complex is voltage sensitive and directly modulated by extracellular calcium. Sci. Adv. 6, eaaz3154 (2020).
Bouasse, M., Impheng, H., Servant, Z., Lory, P. & Monteil, A. Functional expression of CLIFAHDD and IHPRF pathogenic variants of the NALCN channel in neuronal cells reveals both gain- and loss-of-function properties. Sci. Rep. 9, 11791 (2019).
Wie, J. et al. Intellectual disability-associated UNC80 mutations reveal inter-subunit interaction and dendritic function of the NALCN channel complex. Nat. Commun. 11, 3351 (2020).
Shen, H., Liu, D., Wu, K., Lei, J. & Yan, N. Structures of human Nav1.7 channel in complex with auxiliary subunits and animal toxins. Science 363, 1303–1308 (2019).
Jiang, D. et al. Structure of the cardiac sodium channel. Cell 180, 122–134 (2020).
Zhao, Y. et al. Molecular basis for ligand modulation of a mammalian voltage-gated Ca2+ channel Cell 177, 1495–1506 (2019).
Zhao, Y. et al. Cryo-EM structures of apo and antagonist-bound human Cav3.1. Nature 576, 492–497 (2019).
Xu, H. et al. Structural basis of Nav1.7 inhibition by a gating-modifier spider toxin. Cell 176, 702–715 (2019).
Ghezzi, A., Liebeskind, B. J., Thompson, A., Atkinson, N. S. & Zakon, H. H. Ancient association between cation leak channels and Mid1 proteins is conserved in fungi and animals. Front. Mol. Neurosci. 7, 15 (2014).
Hanamura, K. et al. Extracellular phosphorylation of a receptor tyrosine kinase controls synaptic localization of NMDA receptors and regulates pathological pain. PLoS Biol. 15, e2002457 (2017).
Moore, K. L. The biology and enzymology of protein tyrosine O-sulfation. J. Biol. Chem. 278, 24243–24246 (2003).
Heinemann, S. H., Terlau, H., Stühmer, W., Imoto, K. & Numa, S. Calcium channel characteristics conferred on the sodium channel by single mutations. Nature 356, 441–443 (1992).
Yang, J., Ellinor, P. T., Sather, W. A., Zhang, J. F. & Tsien, R. W. Molecular determinants of Ca2+ selectivity and ion permeation in L-type Ca2+ channels. Nature 366, 158–161 (1993).
Tang, L. et al. Structural basis for Ca2+ selectivity of a voltage-gated calcium channel. Nature 505, 56–61 (2014).
Clairfeuille, T. et al. Structural basis of α-scorpion toxin action on Nav channels. Science 363, eaav8573 (2019).
Tao, X., Lee, A., Limapichat, W., Dougherty, D. A. & MacKinnon, R. A gating charge transfer center in voltage sensors. Science 328, 67–73 (2010).
Ahuja, S. et al. Structural basis of Nav1.7 inhibition by an isoform-selective small-molecule antagonist. Science 350, aac5464 (2015).
Shen, H. et al. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 362, eaau2596 (2018).
Lacroix, J. J. et al. Intermediate state trapping of a voltage sensor. J. Gen. Physiol. 140, 635–652 (2012).
Singh, A. K., McGoldrick, L. L. & Sobolevsky, A. I. Structure and gating mechanism of the transient receptor potential channel TRPV3. Nat. Struct. Mol. Biol. 25, 805–813 (2018).
Hughes, T. E. T. et al. Structural insights on TRPV5 gating by endogenous modulators. Nat. Commun. 9, 4198 (2018).
Miller, A. N. & Long, S. B. Crystal structure of the human two-pore domain potassium channel K2P1. Science 335, 432–436 (2012).
Kasap, M., Aamodt, E. J., Sagrera, C. E. & Dwyer, D. S. Novel pharmacological modulation of dystonic phenotypes caused by a gain-of-function mutation in the Na+ leak-current channel. Behav. Pharmacol. 31, 465–476 (2020).
Lee, C. H. & MacKinnon, R. Voltage sensor movements during hyperpolarization in the HCN channel. Cell 179, 1582–1589 (2019).
Bianchi, D. et al. On the mechanisms underlying the depolarization block in the spiking dynamics of CA1 pyramidal neurons. J. Comput. Neurosci. 33, 207–225 (2012).
Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. J. Struct. Biol. 152, 36–51 (2005).
Scheres, S. H. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 180, 519–530 (2012).
Grant, T., Rohou, A. & Grigorieff, N. cisTEM, user-friendly software for single-particle image processing. eLife 7, e35383 (2018).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Rohou, A. & Grigorieff, N. CTFFIND4: Fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Tegunov, D. & Cramer, P. Real-time cryo-electron microscopy data preprocessing with Warp. Nat. Methods 16, 1146–1152 (2019).
Matthies, D. et al. Single-particle cryo-EM structure of a voltage-activated potassium channel in lipid nanodiscs. eLife 7, e37558 (2018).
Rosenthal, P. B. & Henderson, R. Optimal determination of particle orientation, absolute hand, and contrast loss in single-particle electron cryomicroscopy. J. Mol. Biol. 333, 721–745 (2003).
Cardone, G., Heymann, J. B. & Steven, A. C. One number does not fit all: mapping local variations in resolution in cryo-EM reconstructions. J. Struct. Biol. 184, 226–236 (2013).
Kelley, L. A., Mezulis, S., Yates, C. M., Wass, M. N. & Sternberg, M. J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 10, 845–858 (2015).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D 74, 531–544 (2018).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Goddard, T. D. et al. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 27, 14–25 (2018).
Croll, T. I. ISOLDE: a physically realistic environment for model building into low-resolution electron-density maps. Acta Crystallogr. D 74, 519–530 (2018).
Afonine, P. V. et al. New tools for the analysis and validation of cryo-EM maps and atomic models. Acta Crystallogr. D 74, 814–840 (2018).
Williams, C. J. et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 27, 293–315 (2018).
Pettersen, E. F. et al. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Holm, L. Benchmarking fold detection by DaliLite v.5. Bioinformatics 35, 5326–5327 (2019).
Sievers, F. et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7, 539 (2011).
Waterhouse, A. M., Procter, J. B., Martin, D. M., Clamp, M. & Barton, G. J. Jalview Version 2—a multiple sequence alignment editor and analysis workbench. Bioinformatics 25, 1189–1191 (2009).
Robert, X. & Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 42, W320–W324 (2014).
Boone, A. N., Senatore, A., Chemin, J., Monteil, A. & Spafford, J. D. Gd3+ and calcium sensitive, sodium leak currents are features of weak membrane-glass seals in patch clamp recordings. PLoS ONE 9, e98808 (2014).
Pravda, L. et al. MOLEonline: a web-based tool for analyzing channels, tunnels and pores (2018 update). Nucleic Acids Res. 46 (W1), W368–W373 (2018).
Ashkenazy, H. et al. ConSurf 2016: an improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 44 (W1), W344–W350 (2016).
Acknowledgements
We thank members of the Pless laboratory and Genentech colleagues in the Research Materials, BioMolecular Resources and Structural Biology departments for their support of this project; and appreciate the encouragement of A. Rohou, C. Koth, J. Kiefer, S. Hymowitz, V. Dixit and A. Chan. Members of the Pless group acknowledge the Carlsberg Foundation (CF16-0504), the Independent Research Fund Denmark (7025-00097A, 9124-00002B), the Novo Nordisk Foundation (NNF19OC0056438) and the Lundbeck Foundation (R252-2017-1671) for financial support. Reagents are available under a material transfer agreement with Genentech or the appropriate party.
Author information
Authors and Affiliations
Contributions
M.K., C.L.N. and T.C. established protein purification and reconstitution methods. Z.R.L. generated key protein expression reagents. M.K., C.L.N. and C.P.A. optimized cryo-EM sample preparation and data collection. M.K. determined the structure, with guidance from C.C. H.C.C. established methods to record function of the NALCN complex. H.C.C., C.W., O.Ø.B. and A.O.A. performed molecular biology, electrophysiology, biochemistry and pharmacology experiments. M.K., H.C.C., C.L.N., C.W., T.C., O. Ø.B., A.O.A, C.C., S.A.P. and J.P. analysed the data. M.K., H.C.C., C.C., S.A.P. and J.P. wrote the manuscript with input from all authors. C.C., S.A.P. and J.P. supervised the project and are co-senior authors.
Corresponding authors
Ethics declarations
Competing interests
M.K., C.L.N., T.C., Z.R.L., C.P.A., C.C. and J.P. are employees of Genentech/Roche; the other authors declare no competing interests.
Additional information
Peer review information Nature thanks J. David Spafford and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 NALCN function in heterologous expressions systems.
a, b, Representative current traces from Xenopus laevis oocytes (a) and HEK293T cells (b) expressing NALCN alone or with FAM155A, UNC80 and UNC79. Holding potential 0 mV, hyperpolarizing steps from +80 mV to −100 mV in 20 mV increments; oocyte experiments were performed in ND96 recording buffer, HEK293T experiments were performed under symmetrical Na+ conditions. See source data for full experimental details. c, Western blot of total lysate and surface fraction proteins extracted from HEK293T cells expressing the indicated constructs, representative of three independent experiments. d, Representative current traces from oocytes expressing NALCN, FAM155A, UNC80 and UNC79 with NaVβ1 (n = 14), NaVβ2 (n = 10) or CaVα2δ-1 (n = 11). Protocol as above.
Extended Data Fig. 2 NALCN biochemistry and cryo-electron microscopy processing.
a, NALCN purification scheme. b, SDS–PAGE of select samples from the STREP and FLAG affinity purification steps (representative of three or more independent experiments). c, Size-exclusion chromatography of NALCN-FAM155A in lipid nanodiscs (MSP1E3D1), representative of two independent experiments. d, Example cryo-EM micrograph image of NALCN-FAM155A-MSP1E3D1 complex from 15,080 collected micrographs (see f). e, Representative 2D-class averages after 2 rounds of 2D classification from 200 classes and approximately 720,000 particles. f, Data collection and processing workflow. g, Heat map representation of the distribution of assigned particle orientations. h, FSC between two half datasets yields a global resolution estimate of approximately 2.8 Å resolution.
Extended Data Fig. 3 NALCN cryo-EM map.
a, Isosurface rendering of the 3D map with surface coloring according to the local resolution estimated by windowed FSCs. b, Example 3D map overlay for transmembrane regions across DI. c, Example 3D map overlay on representative lipids modelled within the transmembrane region.
Extended Data Fig. 4 Comparison of NALCN with NaV and CaV channel structures.
a, Helical S4-S5 linkers are highlighted by colour in NALCN, NaV1.7 (PDB: 6J8G), CaV1.1 (PDB: 6JPA), and CaV3.1 (PDB: 6KZP); intracellular view (left) and side views (right) are shown. b, Side view of NALCN, CaV1.1, NavPaS (PDB: 6A90) and NaV1.7 highlighting the DIII-DIV linker, CTD, and the β1a-subunit (of CaV1.1).
Extended Data Fig. 5 NALCN structure-based sequence alignment.
Selectivity filter with inner ring (IR, red box) and outer ring (OR, purple box) residues indicated.
Extended Data Fig. 6 Extracellular pore loops, FAM155A sequence alignment (partial), and auxiliary subunit comparisons.
a, Close-in view of the NALCN ECL loops from the DI, DIII and DIV PM, highlighting disulfide bonds (in stick representation), compared to the analogous ECLs from NaV1.7 (PDB: 6J8G), CaV1.1 (PDB: 6JPA), and CaV3.1 (PDB: 6KZP). b, Multi-sequence alignment of selected FAM155A-like proteins; for clarity, spanning only the parts of the cysteine-rich domain (CRD). N-terminal region (171-175) not shown. c, FAM155A-CRD and the MuSK Fzd-like CRD (PDB: 3HKL). d, Superposition of the NaV1.7-β1-β2 complex (PDB: 6J8G) onto NALCN. e, Superposition of the CaV1.1- α2δ-1-β1a-ɣ complex (PDB: 6JPA) onto NALCN.
Extended Data Fig. 7 NALCN pore volume and selectivity filter comparisons.
a, Pore volume rendering65 of NALCN (similar to Fig. 3a), NaV1.7 (PDB: 6J8G), CaV1.1 (PDB: 6JPA), and CaV3.1 (PDB: 6KZP). b, Comparison of selectivity filters of NALCN, NaV1.7 (PDB: 6J8G), CaV1.1 (PDB: 6JPA), and CaV3.1 (PDB: 6KZP). c, NaV1.7-tetrodotoxin (TTX; PDB: 6J8I) and NaV1.7-saxitoxin (STX; PDB: 6J8G) complexes shown alone and superimposed onto NALCN (where NaV1.7 has been omitted for clarity in the superposition).
Extended Data Fig. 8 S6 sequence alignment and NALCN π-bulge.
a, Multi-sequence alignment from select ion channels. Residues that support the S6 π-bulge in NALCN are outlined with a purple box and dashed line indicates the observed hydrogen bond to the backbone carbonyl of the i-4 residue. Residues with side-chains that fill the NALCN pore fenestrations are outlined with a blue box, and residues with side-chains lining the closed S6-activation gate in NALCN are outlined with a green box. b, Side-view of S6 with polar side-chains supporting the π-bulge shown as stick with putative hydrogen bonds (dashed line) to the i-4 backbone carbonyls (*). c, Close-in view of the S6 π-bulges in NALCN. d, TRPV3 channel S6 in apo and ligand bound states (PDB: 6DVW and 6DVZ, respectively), with location of the π-bulge indicated.
Extended Data Fig. 9 NALCN voltage-sensor domain comparisons and features.
a–c, Comparison of the VSDs from KVchim (KV1.2/2.1; PDB: 3LNM), NALCN and NaV1.7 (PDB: 6J8G) with the S2 residue of the HCS shown in green sticks and the S4 gating charges sown in blue sticks. d, S3-S4 of NALCN VSD1-4 with the 310-helix regions indicated. e, Basic residues clustered around the intracellular vestibules of NALCN VSDs are shown as blue sticks; the S2 aromatic residue of the HCS is shown in green sticks.
Extended Data Fig. 10 Characterization of select CLIFAHDD and IHPRF1 mutations.
a, Position of CLIFAHDD (blue) and IHPRF1 (red) mutations on NALCN. b, Exemplar current traces from Xenopus laevis oocytes expressing WT or indicated mutants recorded in ND96 solution (dashed line indicates zero current level). c, Imax–V plots (normalized to averaged outward current at +80 mV of WT complex) from Xenopus laevis oocytes expressing WT (white) or indicated CLIFAHDD/IHPRF1 mutants (blue and red, respectively) recorded in ND96 solution. d, Left panel: western blot of total lysate and surface fraction proteins extracted from HEK293T cells expressing the indicated constructs. NALCN was co-expressed with UNC80, UNC79 and FAM155A (all tagged with C-terminal eGFP-2xFLAG). Right panel: surface expression of NALCN (relative to Na+/K+ATPase expression levels). Data shown as average ± s.d. (n = 3); ns = not statistically different in two-sided paired Student’s t-test.
Extended Data Fig. 11 Conservation analysis and potential protein interaction sites.
a, Conservation analysis66 of NALCN highlighting the FAM155A docking site (FAM155A in grey surface rendering). Sequences used for analysis: Homo sapiens, Mus musculus, Rattus norvegicus, Xenopus tropicalis, Ictalurus punctatus, Danio rerio, Gallus gallus, Columba livia, Drosophila melanogaster, Aedes albopictus, Caenorhabditis elegans, Anisakis simplex. b, Superposition of NALCN, NaV1.7 (PDB: 6J8G), CaV1.1 (PDB: 6JPA), CaV3.1 (PDB: 6KZP) and HCN1 (PDB: 5U6P) based on the S5 segment, with only the S5 and S4-S5 linkers shown for clarity. c, Conservation analysis (as in panel a) shown for the NALCN CTD-region.
Supplementary information
Rights and permissions
About this article

Cite this article
Kschonsak, M., Chua, H.C., Noland, C.L. et al. Structure of the human sodium leak channel NALCN. Nature 587, 313–318 (2020). https://doi.org/10.1038/s41586-020-2570-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2570-8
This article is cited by
-
Extending and outlining the genotypic and phenotypic spectrum of novel mutations of NALCN gene in IHPRF1 syndrome: identifying recurrent urinary tract infection
Neurological Sciences (2023)
-
Network-Based Data Analysis Reveals Ion Channel-Related Gene Features in COVID-19: A Bioinformatic Approach
Biochemical Genetics (2023)
-
Architecture of the human NALCN channelosome
Cell Discovery (2022)
-
Structure and mechanism of NALCN-FAM155A-UNC79-UNC80 channel complex
Nature Communications (2022)
-
Structural architecture of the human NALCN channelosome
Nature (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
