
Abstract
Most sensory information destined for the neocortex is relayed through the thalamus, where considerable transformation occurs1,2. One means of transformation involves interactions between excitatory thalamocortical neurons that carry data to the cortex and inhibitory neurons of the thalamic reticular nucleus (TRN) that regulate the flow of those data3,4,5,6. Although the importance of the TRN has long been recognised7,8,9, understanding of its cell types, their organization and their functional properties has lagged behind that of the thalamocortical systems they control. Here we address this by investigating the somatosensory and visual circuits of the TRN in mice. In the somatosensory TRN we observed two groups of genetically defined neurons that are topographically segregated and physiologically distinct, and that connect reciprocally with independent thalamocortical nuclei through dynamically divergent synapses. Calbindin-expressing cells—located in the central core—connect with the ventral posterior nucleus, the primary somatosensory thalamocortical relay. By contrast, somatostatin-expressing cells—which reside along the surrounding edges of the TRN—synapse with the posterior medial thalamic nucleus, a higher-order structure that carries both top-down and bottom-up information10,11,12. The two TRN cell groups process their inputs in pathway-specific ways. Synapses from the ventral posterior nucleus to central TRN cells transmit rapid excitatory currents that depress deeply during repetitive activity, driving phasic spike output. Synapses from the posterior medial thalamic nucleus to edge TRN cells evoke slower, less depressing excitatory currents that drive more persistent spiking. Differences in the intrinsic physiology of TRN cell types, including state-dependent bursting, contribute to these output dynamics. The processing specializations of these two somatosensory TRN subcircuits therefore appear to be tuned to the signals they carry—a primary central subcircuit tuned to discrete sensory events, and a higher-order edge subcircuit tuned to temporally distributed signals integrated from multiple sources. The structure and function of visual TRN subcircuits closely resemble those of the somatosensory TRN. These results provide insights into how subnetworks of TRN neurons may differentially process distinct classes of thalamic information.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Data availability
All data are available from the corresponding author upon reasonable request.
Change history
27 August 2020
A Correction to this paper has been published: https://doi.org/10.1038/s41586-020-2680-3
References
Jones, E. G. The Thalamus (Cambridge Univ. Press, 2007).
Sherman, S. M. & Guillery, R. W. Functional organization of thalamocortical relays. J. Neurophysiol. 76, 1367–1395 (1996).
Hirsch, J. A., Wang, X., Sommer, F. T. & Martinez, L. M. How inhibitory circuits in the thalamus serve vision. Annu. Rev. Neurosci. 38, 309–329 (2015).
Crabtree, J. W. Functional diversity of thalamic reticular subnetworks. Front. Syst. Neurosci. 12, 41 (2018).
Halassa, M. M. & Acsády, L. Thalamic inhibition: diverse sources, diverse scales. Trends Neurosci. 39, 680–693 (2016).
Fogerson, P. M. & Huguenard, J. R. Tapping the brakes: cellular and synaptic mechanisms that regulate thalamic oscillations. Neuron 92, 687–704 (2016).
Guillery, R. W., Feig, S. L. & Lozsádi, D. A. Paying attention to the thalamic reticular nucleus. Trends Neurosci. 21, 28–32 (1998).
Pinault, D. The thalamic reticular nucleus: structure, function and concept. Brain Res. Rev. 46, 1–31 (2004).
Crick, F. Function of the thalamic reticular complex: the searchlight hypothesis. Proc. Natl Acad. Sci. USA 81, 4586–4590 (1984).
Diamond, M. E., von Heimendahl, M., Knutsen, P. M., Kleinfeld, D. & Ahissar, E. ‘Where’ and ‘what’ in the whisker sensorimotor system. Nat. Rev. Neurosci. 9, 601–612 (2008).
Mease, R. A., Sumser, A., Sakmann, B. & Groh, A. Cortical dependence of whisker responses in posterior medial thalamus in vivo. Cereb. Cortex 26, 3534–3543 (2016).
Sherman, S. M. Thalamus plays a central role in ongoing cortical functioning. Nat. Neurosci. 19, 533–541 (2016).
Fishell, G. & Rudy, B. Mechanisms of inhibition within the telencephalon: “where the wild things are”. Annu. Rev. Neurosci. 34, 535–567 (2011).
Hou, G., Smith, A. G. & Zhang, Z. W. Lack of intrinsic GABAergic connections in the thalamic reticular nucleus of the mouse. J. Neurosci. 36, 7246–7252 (2016).
Hu, H., Cavendish, J. Z. & Agmon, A. Not all that glitters is gold: off-target recombination in the somatostatin-IRES-Cre mouse line labels a subset of fast-spiking interneurons. Front. Neural Circuits 7, 195 (2013).
Lam, Y. W. & Sherman, S. M. Functional organization of the thalamic input to the thalamic reticular nucleus. J. Neurosci. 31, 6791–6799 (2011).
Lee, S. H., Govindaiah, G. & Cox, C. L. Heterogeneity of firing properties among rat thalamic reticular nucleus neurons. J. Physiol. (Lond.) 582, 195–208 (2007).
Diamond, M. E., Armstrong-James, M. & Ebner, F. F. Somatic sensory responses in the rostral sector of the posterior group (POm) and in the ventral posterior medial nucleus (VPM) of the rat thalamus. J. Comp. Neurol. 318, 462–476 (1992).
Roth, M. M. et al. Thalamic nuclei convey diverse contextual information to layer 1 of visual cortex. Nat. Neurosci. 19, 299–307 (2016).
Audette, N. J., Bernhard, S. M., Ray, A., Stewart, L. T. & Barth, A. L. Rapid plasticity of higher-order thalamocortical inputs during sensory learning. Neuron 103, 277–291.e4 (2019).
Zhang, W. & Bruno, R. M. High-order thalamic inputs to primary somatosensory cortex are stronger and longer lasting than cortical inputs. eLife 8, e44158 (2019).
Steriade, M., Domich, L. & Oakson, G. Reticularis thalami neurons revisited: activity changes during shifts in states of vigilance. J. Neurosci. 6, 68–81 (1986).
McCormick, D. A. & Bal, T. Sleep and arousal: thalamocortical mechanisms. Annu. Rev. Neurosci. 20, 185–215 (1997).
Marlinski, V. & Beloozerova, I. N. Burst firing of neurons in the thalamic reticular nucleus during locomotion. J. Neurophysiol. 112, 181–192 (2014).
Whitmire, C. J., Waiblinger, C., Schwarz, C. & Stanley, G. B. Information coding through adaptive gating of synchronized thalamic bursting. Cell Rep. 14, 795–807 (2016).
Llinás, R. R. & Steriade, M. Bursting of thalamic neurons and states of vigilance. J. Neurophysiol. 95, 3297–3308 (2006).
Clemente-Perez, A. et al. Distinct thalamic reticular cell types differentially modulate normal and pathological cortical rhythms. Cell Rep. 19, 2130–2142 (2017).
Destexhe, A., Contreras, D., Steriade, M., Sejnowski, T. J. & Huguenard, J. R. In vivo, in vitro, and computational analysis of dendritic calcium currents in thalamic reticular neurons. J. Neurosci. 16, 169–185 (1996).
Llinás, R. & Jahnsen, H. Electrophysiology of mammalian thalamic neurones in vitro. Nature 297, 406–408 (1982).
Lambert, R. C., Bessaïh, T., Crunelli, V. & Leresche, N. The many faces of T-type calcium channels. Pflugers Arch. 466, 415–423 (2014).
Kimura, A. & Imbe, H. Anatomically structured burst spiking of thalamic reticular nucleus cells: implications for distinct modulations of sensory processing in lemniscal and non-lemniscal thalamocortical loop circuitries. Eur. J. Neurosci. 41, 1276–1293 (2015).
Pinault, D., Bourassa, J. & Deschênes, M. The axonal arborization of single thalamic reticular neurons in the somatosensory thalamus of the rat. Eur. J. Neurosci. 7, 31–40 (1995).
McAlonan, K., Cavanaugh, J. & Wurtz, R. H. Guarding the gateway to cortex with attention in visual thalamus. Nature 456, 391–394 (2008).
Lee, S. C., Patrick, S. L., Richardson, K. A. & Connors, B. W. Two functionally distinct networks of gap junction-coupled inhibitory neurons in the thalamic reticular nucleus. J. Neurosci. 34, 13170–13182 (2014).
Crabtree, J. W. & Isaac, J. T. New intrathalamic pathways allowing modality-related and cross-modality switching in the dorsal thalamus. J. Neurosci. 22, 8754–8761 (2002).
Kimura, A. & Imbe, H. Robust subthreshold cross-modal modulation of auditory response by cutaneous electrical stimulation in first- and higher-order auditory thalamic nuclei. Neuroscience 372, 161–180 (2018).
Briggs, F. & Usrey, W. M. Emerging views of corticothalamic function. Curr. Opin. Neurobiol. 18, 403–407 (2008).
Crandall, S. R., Cruikshank, S. J. & Connors, B. W. A corticothalamic switch: controlling the thalamus with dynamic synapses. Neuron 86, 768–782 (2015).
Wimmer, R. D. et al. Thalamic control of sensory selection in divided attention. Nature 526, 705–709 (2015).
Nakajima, M., Schmitt, L. I. & Halassa, M. M. Prefrontal cortex regulates sensory filtering through a basal ganglia-to-thalamus pathway. Neuron 103, 445–458.e10 (2019).
Beierlein, M. Synaptic mechanisms underlying cholinergic control of thalamic reticular nucleus neurons. J. Physiol. (Lond.) 592, 4137–4145 (2014).
Conley, M., Kupersmith, A. C. & Diamond, I. T. The organization of projections from subdivisions of the auditory cortex and thalamus to the auditory sector of the thalamic reticular nucleus in galago. Eur. J. Neurosci. 3, 1089–1103 (1991).
Conley, M., Schmechel, D. E. & Diamond, I. T. Differential distribution of somatostatin-like immunoreactivity in the visual sector of the thalamic reticular nucleus in galago. Eur. J. Neurosci. 3, 237–242 (1991).
Graybiel, A. M. & Elde, R. P. Somatostatin-like immunoreactivity characterizes neurons of the nucleus reticularis thalami in the cat and monkey. J. Neurosci. 3, 1308–1321 (1983).
Daigle, T. L. et al. A suite of transgenic driver and reporter mouse lines with enhanced brain-cell-type targeting and functionality. Cell 174, 465–480. e22 (2018).
Agmon, A. & Connors, B. W. Thalamocortical responses of mouse somatosensory (barrel) cortex in vitro. Neuroscience 41, 365–379 (1991).
Jackman, S. L., Beneduce, B. M., Drew, I. R. & Regehr, W. G. Achieving high-frequency optical control of synaptic transmission. J. Neurosci. 34, 7704–7714 (2014).
Acknowledgements
We thank Z. Fu, G. Feng and their associates for interactions surrounding this project. We also thank S. Crandall, O. Ahmed, M. Zervas, D. Lipscombe, C. Chen, G. Manzano, S. Baskoylu, F. Pouille, B. Theyel and F. S. Susi for discussions. This work was supported by grants R01 NS100016, P20 GM103645, NSF 1738633, NSFGRFP 1058262 and NSF 1632738.
Author information
Authors and Affiliations
Contributions
R.I.M.-G., B.V., B.W.C. and S.J.C. designed the experiments. R.I.M.-G., B.V., J.B.Z., S.L.P., T.R.S. and S.J.C. conducted the experiments. R.I.M.-G., B.V., J.B.Z. and S.J.C. analysed the results. R.I.M.-G., B.V., B.W.C. and S.J.C. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Karel Svoboda and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Somatostatin cells form a shell around a calbindin-rich central core of the somatosensory TRN; nearly all neurons in the somatosensory TRN express parvalbumin.
a, Epifluorescence images showing serial sections of live tissue expressing AAV9-DIO-GFP (9 days of expression) through the TRN of two P23 SOM-Cre mice. Anterior-posterior (AP) positions of the sections relative to bregma are indicated. Notice how the SOM cells form a shell-like structure surrounding the central core of the somatosensory TRN (centred ~ −1.4 mm from bregma) (experiment replicated 12 times). b, Top, dot plots showing the locations of the genetically or immunohistochemically labelled TRN cell types for sections at different AP distances from bregma. Bottom, proportion of cells expressing PV, CB, SOM-Cre × tdT, and combinations of the markers. Quantification was restricted to the boxed regions shown in the top panels (cell counts: left, 947; middle, 1,075; right, 843; 3 sections from 3 mice). The proportion of CB cells was highest in the middle section of the somatosensory TRN (−1.4 mm), whereas the proportion of SOM-Cre × tdT cells was lowest in the middle section and higher in the more anterior and posterior sections (P < 0.001, χ2 test with Yates’ correction). Data are mean ± s.e.m. c, Left, confocal images of the somatosensory sector of TRN in a PV-Cre × tdT mouse (red) stained for NeuN (green). Right, quantification of expression. In PV-Cre × tdT mice, 98.09% of TRN neurons were double positive for tdT and NeuN, 0.73% were positive only for tdT, and 1.18% were positive only for NeuN (1,218 neurons, 2 sections, 1 mouse). d, Same as c but tissue was stained immunohistochemically for parvalbumin. In PV-Cre × tdT mice, 98.66% of TRN neurons were double-positive for tdT and PV, 0.34% were positive only for tdT and 0.34% were positive only for PV (1,177 neurons, 2 sections, 1 mouse). e, DIC-IR (left) and tdT fluorescence (right) images of TRN in a live slice from a PV-Cre × tdT mouse. Asterisk indicates a soma that did not express tdT. During such live imaging, tdT-negative somata were relatively rare in TRN (>80% of somata visualized in DIC-IR expressed tdT; n = 5 slices, 3 mice; right panel). f, All tdT-negative cells targeted for recording (n = 8/8) had physiological properties of glia. Left, TRN glial cells had hyperpolarized resting potentials (typically less than −85 mV) and no action potentials even during large depolarizations.
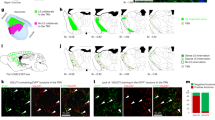
Extended Data Fig. 2 Cells expressing Sst and Calb1 mRNA are topographically segregated in the somatosensory TRN.
a, Left, confocal image overlay of expression of Sst and Calb1 mRNA (encoding SOM and CB) through the TRN of a PV-Cre × tdT mouse. In the somatosensory TRN (boxed region), Sst expressing cells are preferentially localized in the medial and lateral edge zones, whereas Calb1 cells are generally in the central zone. Insets show mRNA expression patterns of cells in the lateral edge (top row), central (middle row), and medial edge (bottom row) zones. Although most cells predominantly expressed either Sst (red) or Calb1 (green) mRNA, some (about 10%) expressed both (arrow). b, Proportion of cells expressing various combinations of markers for each zone (for b–e, 593 cells, 6 sections, 6 mice; average of 98.9 ± 8.5 cells per section). 99.1 ± 0.2% of cells expressed PV. Most cells in the medial and lateral edge zones expressed only Sst and Pvalb, but not Calb1 (about 64%). By contrast, most central cells expressed only Calb1 and Pvalb, but not Sst (about 76%). c, Normalized Sst (left) or Calb1 (right) mRNA expression as a function of medial–lateral position in the the TRN (0 = most medial, 1 = most lateral). Cells expressing Sst but not Calb1 were concentrated near the edges (red) (mean Sst expression for edge vs central: 0.325 ± 0.020 versus 0.052 ± 0.006, P < 0.0001, unpaired two-tailed t-test). Cells in the centre generally had high levels of Calb1 but not Sst (mean Calb1 expression for central vs edge: 0.326 ± 0.012 versus 0.086 ± 0.012, P < 0.0001, unpaired two-tailed t-test). Around 10% of somatosensory TRN cells expressed both Sst and Calb1 (yellow) and clustered in the lateral half of the TRN, with their focus being near the boundary between the central and lateral zones (average normalized medial–lateral position 0.71 ± 0.02). Compared with the cells expressing Sst or Calb1 ‘alone’, the dual Sst/Calb1 cells had lower expression of both Sst (Sst/Calb1 cells: 0.25 ± 0.02 vs Sst only: 0.43 ± 0.02, P < 0.0001, unpaired two-tailed t-test) and Calb1 mRNA (Sst/Calb1 cells: 0.23 ± 0.15 vs Calb1 only: 0.39 ± 0.01, P < 0.0001, unpaired two-tailed t-test). Together this suggests they may represent a transitional population. About 9% of somatosensory TRN cells expressed PV only (grey) and were dispersed fairly evenly across the sector (mean medial-lateral position: 0.38 ± 0.03). d, Relative expression of Sst versus Calb1 (within-cell) as a function of medial-lateral position in the TRN (a value of 1.0 on the y-axis index indicates Sst expression only, a value of 0 indicates Calb1 only, and 0.5 indicates equal expression values for Sst and Calb1). Most cells (80%) had very high (>0.9) or very low (<0.1) relative expression scores, indicating most cells have strong biases towards a single marker (10:1 ratios or greater). Consistent with Fig. 1 and a–c, Sst-biased cells tended to cluster towards the TRN edges, whereas Calb1-biased were located centrally (edge versus central preference scores: 0.76 ± 0.03 versus 0.14 ± 0.01, P < 0.0001, paired two-tailed t-test). A moving average (11-point) is superimposed to illustrate the spatial trends in expression bias. Interestingly, even the dual-expressing Sst/Calb1 cells (yellow) tended to have a topographical preference according to relative Sst vs Calb1 expression: dual-expressing edge cells tended to have higher Sst:Calb1 preference scores than dual-expressing central cells (dual edge versus central cells: 0.58 ± 0.04 versus 0.47 ± 0.04, P = 0.06, unpaired two-tailed t-test). e, Calb1 expression as a function of Sst expression for cells in the medial, central, lateral, and combined zones. P values shown in the panels are the results of two-tailed paired (within-cell) t-tests comparing Calb1 and Sst expression. Data are mean ± s.e.m.
Extended Data Fig. 3 In the somatosensory TRN of postnatal SOM-Cre mice, both Cre expression and viral Cre-dependent protein expression are restricted mainly to SOM cells along the edges.
a, Epifluorescence images of virally induced, SOM-Cre-dependent GFP expression patterns in TRN (live 300-μm-thick slices from 7 mice). Virus carrying a Cre-dependent GFP gene (AAV9-DIO-GFP) was injected throughout the TRNs of SOM-Cre × tdTomato or SOM-Cre × ICR mice. This drove GFP expression (green) in SOM cells that expressed Cre at or after the time of the virus injection (age at injection P17.1 ± 1.18 days; age at imaging 26.8 ± 1.19). Mouse 1: images of GFP alone and overlay of GFP and genetically labelled tdTomato cells. Mice 2–7: GFP alone. Within the somatosensory TRN, expression of SOM-Cre × GFP was restricted to the edges of the nucleus and excluded from the central zone after early development. Experiment replicated 12 times. b, Same as in a except using PV-Cre mice (age at virus injection P14; age at imaging P23). Nearly all of the TRN cells expressed both genetically labelled PV-Cre × tdTomato and virally induced PV-Cre × GFP. In addition to demonstrating ubiquitous postnatal PV-Cre expression across the TRN, these results show that the virus has no obvious tropisms for different topographic regions of the TRN. Experiment replicated 4 times. c, Confocal images of Pvalb, Sst, and Cre mRNA expression through the TRN of a P27 SOM-Cre heterozygous mouse (18 μm thick section). The overall patterns of Sst and Cre mRNA labelling were nearly identical (experiment replicated in 5 sections from 4 mice). d, Sst and Cre mRNA were generally expressed in the same neurons (total counts: 167 cells, 5 sections, 4 mice; measured in the somatosensory sector of the TRN). Of the Cre-expressing cells, 88.3 ± 0.1% expressed Sst mRNA. Of the Sst-expressing cells, 96.1 ± 0.01% expressed Cre mRNA. e, AAV9-DIO-GFP injections into the TRNs of postnatal SOM-Cre mice led to GFP expression patterns that were very similar to Sst mRNA patterns. Left, DIC and epifluorescence images from a live 300-μm-thick section expressing AAV9-DIO-GFP (9 days after virus injection) in a SOM-Cre heterozygous mouse. This experiment was replicated 12 times (see a, Fig. 1g and Extended Data Fig. 1a for additional examples). f, Same as e but virus was injected into a wild-type mouse (ICR strain; no Cre expression) (tested in n = 2 mice). Matching display settings were used for the slices in c and d. The lack of fluorescent signal indicates that the AAV-DIO-GFP virus used here drives GFP expression only in Cre-expressing cells. Data are mean ± s.e.m.
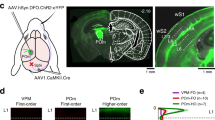
Extended Data Fig. 4 VP afferents target the CB-dense centre of the somatosensory TRN, whereas POM afferents target the SOM-dense medial and lateral edges.
a, Four representative examples showing the POM injection sites and anterograde projections to TRN in live 300-μm slices. Red: SOM-Cre × tdT. Green: ChR2–eYFP. Boundaries of the thalamic nuclei (TRN outlined by dashed lines) were determined from SOM-Cre × tdT fluorescence and by bright-field images (not shown). Injection sites were located in POM nuclei, seen here as the large, bright eYFP spots medial to TRN. The smaller bands of eYFP along the medial and lateral edges of the somatosensory TRN are axons and terminals from the labelled POM cells. In addition to the TRN edges, POM also targeted layer 5a (L5a) and L1 of barrel cortex (but not L4; data not shown). Top left image shows the injection site for Fig. 2c, d. Experiment replicated in 18 mice. b, Left, schematic showing recording configuration (also applies to the experiments of Fig. 2c–f). Right, additional representative examples of synaptic responses of cells across the medial–lateral axis of somatosensory TRN evoked by POM stimulation. TRN cells near the edges of the somatosensory TRN received strong input from the POM whereas cells located in the centre of the TRN did not. Experiments replicated in 5 slices, 5 mice. c, Similar to a, but for VP afferents. VP afferents clearly targeted the central zone of TRN. VP also projected to L4 of the barrel cortex but not to L5a or L1 (not shown). Top left image shows the injection site for Fig. 2g, h. Experiments were replicated in 25 mice. d, Similar to b, but for VP. Left, schematic showing recording configuration (also applies to the experiments of Fig. 2g–j). Right, cells in the centre of the somatosensory TRN received stronger input from the VP than did cells located near the edges of the TRN. The experiment was replicated in 6 slices, 4 mice.
Extended Data Fig. 5 Comparison of EPSC kinetics and short-term synaptic depression based on topography or reporter labelling.
a, Schematic showing approximate locations of TRN somata targeted for each subcircuit. EPSCs in medial and lateral edge cells were examined separately here, using the dataset from Fig. 3a–c. V-clamp at −84 mV. b, Kinetics of the EPSCs. Left, optically evoked EPSCs of example cells from each group (stimulus 1 of c). EPSCs evoked by POM inputs to medial and lateral edge cells were broader, with greater fractional areas in the later portions of the responses than EPSCs evoked by VP inputs to central cells. Right, kinetic differences between responses are shown for two methods of quantification. The first is the ratio of the EPSC area during the last 90 ms of the response (10–100 ms from stimulus onset) to the full EPSC area (0–100 ms from stimulus onset; left graph). This ratio, which describes the fractional area of the late portion of the response, varied among cells groups (ANOVA, P < 0.0001). Central values were significantly lower than medial and lateral edge values (two-tailed Bonferroni’s t-tests, P < 0.002 and P < 0.002, respectively), whereas the two edge groups were not different from one another (two-tailed Bonferroni’s t-test, P = 0.99). As a second method of quantifying response kinetics we calculated the ratio of the full EPSC area (0–100 ms from stimulus onset) to the peak EPSC amplitude (right graph, as in Fig. 3b). This ratio also varied as a function of cell group (ANOVA, P < 0.0005). Again, central values were significantly lower than medial and lateral edge values (two-tailed Bonferroni’s t-tests, P < 0.002 and P < 0.03, respectively), whereas the two edge groups were not different from one another (two-tailed Bonferroni’s t-test, P = 0.99). The half-widths of EPSCs were not significantly different between any of the groups (all P > 0.05 for two-tailed Bonferroni’s t-tests; data not shown). For b, c, central cells: 48 cells from 14 mice; medial cells: 13 cells from 11 mice, lateral cells: 8 cells from 7 mice. c, Left, representative traces illustrating the short-term synaptic depression of optically evoked EPSCs across groups (10 Hz trains of LED stimuli: arrows). Right, average EPSC areas measured during the first 10 ms of the responses. Depression of VP-evoked responses in central cells was stronger than depression of POM-evoked responses in medial or lateral cells (two-tailed Bonferroni’s t-tests; central versus lateral and central versus medial, both P < 0.0001), whereas depression in the two edge cell groups did not differ significantly (two-tailed Bonferroni’s t-test, P = 0.99). d, Left, schematic showing experimental design. Recordings were made in slices from SOM-Cre × tdT (Ai14) mice. ChR2–eYFP was virally expressed in VP cells. We optically stimulated the VP cells/axons and recorded evoked synaptic responses from closely spaced tdT-positive and tdT-negative neurons in the central zone of the TRN. We ensured that both cells had similar eYFP terminal labelling surrounding them; usually the cells were separated by less than 50 μm. The goal was to test whether there were response differences between central TRN cells that were SOM-Cre × tdT-positive versus SOM-Cre × tdT-negative. Neurons were tdT-positive if they expressed Cre recombinase at any stage of their lives; in SOM-Cre mice, TRN central zone neurons seemed to express Cre only early in development (see text, Fig. 1b, e, Extended Data Figs. 1a, 2, 3). Middle and right, average optically evoked synaptic responses (1 ms LED pulses, 10 Hz trains) for tdT-positive and tdT-negative cell groups for stimuli delivered at 2× (middle) and 16× (right) the threshold for evoking a clear EPSC. Threshold intensity was not significantly different between tdT-positive and tdT-negative cells. The average LED power used at 2× threshold was 2.13 mW. Response magnitudes, short-term EPSC depression, and kinetics (inset traces) did not differ between tdT-positive and tdT-negative cells at 2× threshold (n = 19 pairs of cells) or at 16× threshold (n = 12 pairs of cells) (ANOVA, P = 0.98 and P = 0.61, respectively; all P > 0.05 for cell-type comparisons on individual stimuli in the trains, two-tailed Bonferroni’s t- tests). Thus, VP synaptic inputs seem to be no different for central TRN cells that express SOM-Cre during early development than for those that are SOM-Cre negative continuously. Data are mean ± s.e.m.
Extended Data Fig. 6 Differences in short-term depression between the two somatosensory thalamoreticular pathways do not depend on axon terminal stimulation, stimulus strength, or the temporal components of the responses measured.
Some studies have shown that synaptic release probability and short-term plasticity can be abnormal when optogenetic light stimuli are directed at opsin-expressing presynaptic terminals47. We addressed this issue in the experiments described in a–c. a, VP synapses onto central TRN cells undergo greater short-term depression than POM synapses onto edge cells when optically stimulating the thalamocortical (TC) terminals within TRN. Left, schematic showing recording configuration when optically stimulating over the terminals of the ChR2-expressing TC cells. For the experiments of a–c, GABAA receptors and T-type calcium channels were blocked with picrotoxin (50 μM) and TTA-P2 (1 μM), respectively. Right, average short-term synaptic depression. Light intensities were chosen to evoke around 1.5–3 nA peak responses. EPSC areas were measured over the first 10 ms of each response. There was greater short-term depression for the VP → central cell pathway (n = 20 cells, 5 mice) than the POM → edge cell pathway (n = 17 cells, 8 mice) (P < 0.0001, ANOVA stimulus 2–10; all P < 0.005 from stimulus 5–10, two-tailed Bonferroni’s t-test). b, VP inputs to central cells remain more depressing than POM inputs to edge cells when stimulating within the TC nuclei. Left, schematic showing recording configuration when optically stimulating over cell bodies and proximal axons of the ChR2-expressing TC cells. Same cells as a. Right, average short-term synaptic depression was greater in central than edge cells (P < 0.0001, ANOVA stimulus 2–10; all P < 0.05 from stimulus 5, 7–10, two-tailed Bonferroni’s t-test). This pattern of short-term depression in central cells (VP inputs) and edge cells (POM inputs) did not differ from the previously observed pattern in the same group of cells when stimulating over the TC terminals (a) or in other experiments of the study (d, e, Fig. 3, Extended Data Fig. 5). c, Optical control indicating that light scattering and spread to the TC terminals did not trigger the responses observed when stimulating within TC nuclei (that is, the responses in b). Left, schematic showing recording configuration. The LED was directed to ventrally located ‘control spots’, away from the ChR2-expressing TC pathway but equally distant from the TC terminals as the effective stimulus spots within the TC nuclei. Therefore, if synaptic responses were observed, they would be triggered by light spreading to the ChR2-expressing TC terminals within the TRN. The same cells were recorded as in a, b. Right, EPSC areas when stimulating over the TC nuclei versus the control spots using matching LED parameters (intensities and spot sizes). No clear responses were evoked when stimulating over the control spots, suggesting that light spread to the terminals did not drive the responses when stimulating within the TC nuclei (all P < 0.0001; two-tailed paired t-tests). d, Schematic showing recording configuration for e, f when optically stimulating VP inputs to central cells and POM inputs to edge cells. ChR2-expressing VP and POM inputs were photostimulated at their terminals (1 ms LED pulse, 10 Hz stimuli) and EPSCs were recorded in central and edge TRN cells, respectively. e, Differences in short-term depression of central and edge cells does not change with varying input strength. Maximum response amplitudes tended to be larger for central than for edge cells. Here we compared short-term depression of synaptic responses in central and edge cells, varying stimulus intensities for the central cells so that their responses were either weaker (top) or stronger (bottom) than those of edge cells (mean central EPSCs: 1.3 ± 0.2 nA and 5.5 ± 0.8 nA, respectively; mean edge EPSCs 2.1 ± 0.3 nA). There were no major differences in patterns of short-term synaptic depression for the weaker versus stronger responses. In both cases, there was more depression for the central cells than the edge cells (P < 0.0001, ANOVA stimulus 2–10; all P < 0.05 from stimulus 5–10, two-tailed Bonferoni’s t-test; central weak responses: n = 45 cells, 11 mice; central strong responses: n = 32 cells, 9 mice; edge cells: n = 21 cells, 14 mice). Central responses were acquired at approximately 2× the response threshold for the top row (average LED power 1.92 mW) and approximately 16× response threshold for the bottom row (average LED power 12.5 mW). f, Short-term synaptic depression is similar whether EPSCs are measured only during the initial fast components of the responses (the first 10 ms) or across longer durations (100 ms). Top, normalized EPSC area dynamics for the initial 10 ms of the responses (same plot shown on Fig. 3c). The EPSC amplitudes were similar for central and edge cells to stimulus 1 (mean central EPSCs: 2.0 ± 0.2 nA; mean edge EPSCs 2.1 ± 0.3 nA, unpaired two-tailed t-test, P = 0.93). There was greater short-term depression for the central cells than for the edge cells (P < 0.0001, ANOVA stimulus 2–10; all P < 0.005 from stimulus 5-10, two-tailed Bonferroni’s t-test; central cells: 48 cells, 14 mice; edge: 21 cells, 14 mice). Bottom, normalized area dynamics for the full responses (0–100 ms). Again, there was greater short-term depression for the central cells than for the edge cells (P < 0.0001, ANOVA stimulus 2–10; P < 0.05 for stimulus 8–10, two-tailed Bonferroni’s t-test). Thus, the overall short-term synaptic depression patterns are similar for early and later components of the EPSCs. Data are mean ± s.e.m.
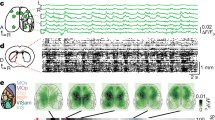
Extended Data Fig. 7 Intrinsic cellular properties vary according to medial–lateral position in the TRN.
a–c, Inset, voltage responses to hyperpolarizing current steps for representative central and edge cells. Throughout this figure, central cells are coloured black and edge cells are red. Note the higher input resistance for the edge cell (316 vs 152 MΩ). Plots show passive membrane properties for central and edge cells as a function of soma position (far medial edge = position 0, far lateral edge = position 1; steady-state Vm ≈ −84 mV). Open circles indicate individual cell measurements. Filled circles are averages for the cell groups: medial edge, central and lateral edge. d, Central cells have larger somas than edge cells. ROIs outlining complete somas were hand-drawn with the aid of ROIManager (ImageJ) using the PV-IHC tissue from Fig. 1 (n = 575 cells; 3 sections; 3 mice). The mean soma area for central and edge were 110.1 μm2 vs 95.6 μm2, respectively (unpaired two-tailed t-test, P < 0.0001). Cell areas seem to change gradually with location. e, Representative examples of edge cell and central cell action potentials (APs). f–h, AP properties as a function of soma location. Half-width (f), AP maximum rate-of-rise (g), AP maximum rate-of-repolarization (h). Edge cells have wider APs due to slower repolarization slopes, suggesting differences in potassium conductances. i, Representative AP afterpotentials of each cell type. Edge cells generally have more hyperpolarized fast-afterhyperpolarizations (fast-AHPs), afterdepolarizations (ADPs), and slow-afterhyperpolarizations (slow-AHPs). j–l, Fast-AHP (j), ADP (k) and slow-AHP (l) as a function of somatic location (APs evoked from a steady state Vm ≈ −74 mV; values relative to AP threshold). m, Representative examples of repetitive tonic firing evoked by 75 pA current steps (from −74 mV baseline potentials) for central and edge cells. n, Spike frequency vs current intensity (FI) plots. Closed circles show the frequency for the first 2 spikes in each train. Open circles show the frequency for the last 2 spikes. The initial firing frequency is similar between edge and central cells at multiple current steps. However, edge cells fire at higher frequencies than central cells by the end of the current pulses, perhaps because edge cells have higher input resistances (for n, o, central cells n = 29, edge cells n = 40). o, Central cells have more spike frequency adaptation than edge cells (unpaired two-tailed t-test P < 0.0001). p–r, Same as m–o, except responses were evoked from steady-state potentials of −84 mV to elicit intrinsic bursts. Central cells showed more pronounced bursts than edge cells. On average, central cells required stronger currents to evoke spiking than edge cells when stimulated from −84 mV steady-state potentials (q) (see Supplementary Information 1 for more threshold information). However, the initial spike frequencies for central cells were much higher than those for edge cells due to their bursty nature (q). The frequencies by the ends of the trains were nearly identical for central and edge cells (q). Central cells had stronger spike frequency adaptation than edge cells (r), largely due to stronger initial bursting in central cells (unpaired two-tailed t-test, P < 0.0001; for q, r: central cells n = 31, edge cells n = 42). For the physiological measurements in this figure, central and edge cell recordings were interleaved within slices (drawn from at least 8 mice per measured property). Data are mean ± s.e.m. See Supplementary Information 1 for additional intrinsic physiological properties and statistical comparisons.
Extended Data Fig. 8 Inhibitory output from the TRN: SOM-expressing TRN edge cells project to the POM, whereas CB cells project to the VPM.
a, Fluorescence (left) and DIC (middle) images from a live 300-μm slice following ChR2–eYFP expression in CB-Cre-positive TRN cells. Fluorescent axons of these TRN CB cells target the VPM but not the POM. White and red circles show the locations of recorded VPM and POM neurons, respectively. Right, strong IPSCs were evoked in the VPM neurons (black) whereas almost no IPSCs were evoked in the POM neurons (red) by optical activation of CB-Cre TRN cells (each trace is the response for one cell, averaged over 10 sweeps; Vhold −34 mV). Arrows indicate onsets of LED flashes (1 ms duration). Glutamate receptors were blocked by applying APV (50 μM) and DNQX (20 μM). The experiment was replicated in 4 slices from 3 CB-Cre mice. b, Same as a except ChR2–eYFP expression was in SOM-Cre TRN edge cells. Notice both the strong axonal targeting and IPSCs in the POM, rather than in the VPM. The experiment was replicated in 6 slices from 5 SOM-Cre mice. c, Left, group plot comparing fluorescence projection intensities to the VPM and the POM from the two TRN cell types. Fluorescence was normalized to the maximum level across the thalamic relay nuclei, with zero values set to the minimum level in the slice (n = 6 slices from 5 SOM-Cre mice; n = 4 slices from 3 CB-Cre mice; dashed lines represent a second slice for one mouse). Projections of CB-Cre TRN cells were stronger in the VPM than in the POM, whereas projections of SOM-Cre TRN cells were stronger in the POM than in the VPM (all P < 0.0001, two-tailed Bonferroni’s t-tests). Right, strengths of IPSCs recorded in the VPM and the POM reflected their anatomical inputs from the TRN. IPSCs evoked by photostimulation of CB-Cre TRN central cells (black) were large in the VPM (n = 15, 3 mice) and very weak in the POM (n = 16, 3 mice) (two-tailed Bonferroni’s t- test, P < 0.0001). Conversely, IPSCs evoked by stimulation of SOM-Cre edge cells (red) were strong in the POM (n = 12 cells, 4 mice) and undetectable in the VPM (n = 15 cells, 4 mice) (two-tailed Bonferroni’s t- test, P < 0.0001). Open circles show response areas for individual cells. Closed circles show averages per mouse. Comparisons were made using matched LED power. Data are mean ± s.e.m.
Extended Data Fig. 9 Visual TRN and somatosensory TRN have similar centre and edge subcircuits.
a, Representative images showing dLGN and LP injection sites and anterograde projections to the TRN. dLGN projects to the central zone whereas LP projects to the edges of the visual sector of the TRN. This experiment was replicated in 5 mice with dLGN viral injections, and in 13 mice with LP viral injections. b, Two representative examples (left and middle panels) of synaptic responses from cells across the medial-lateral axis of visual TRN evoked by photostimulation of dLGN. Cells in the centre of visual TRN received strong input from dLGN, whereas inputs to cells located near the edges of TRN were very weak or undetectable (average of 10 sweeps, −84 mV). Right, relationship between soma location in TRN and synaptic response amplitude (n = 19 cells, 6 slice preparations, each preparation represented by a different symbol shape; means for the 3 zones are represented by filled symbols and connected by lines), showing that cells in the centre of visual TRN respond strongly to dLGN input and those in the medial and lateral edges have minimal dLGN inputs (as in Fig. 2, and Extended Data Fig. 4). c, Same as b except characterizing synaptic inputs to TRN from LP. Cells along the medial edge (and in some cases along the lateral edge) of visual TRN received strong input from LP. By contrast, cells located in the centre of visual TRN generally had weak or no responses to LP input (average of 10 sweeps, −84 mV; n = 49 cells,12 preps). d, Dynamic properties of visual thalamoreticular responses. To study short-term synaptic depression, photostimulation intensities were chosen to evoke peak responses of around 2.5 nA (average LED powers: 20.8 mW for LP, 6.6 mW for dLGN). EPSC areas were measured over the first 10 ms of each response. Short-term depression was more pronounced for the central cells than the edge cells (P < 0.0001, ANOVA, stimulus 2–10). Depression in the two edge cell groups did not differ (two-tailed Bonferroni’s t-test, all P > 0.99). Depression of dLGN-evoked responses in central cells was stronger than depression of LP-evoked responses in medial or lateral cells (two-tailed Bonferroni’s t-test; central vs medial, central vs lateral, both P < 0.0001; central group: 7 cells, 5 mice; medial group: 17 cells, 11 mice; lateral group: 7 cells, 7 mice). Analogous to subcircuits of the somatosensory TRN, dLGN-evoked EPSCs of central cells in the visual TRN were less prolonged than LP-evoked EPSCs of edge cells (not shown). For example, the ratio of the EPSC areas to the peak EPSC amplitudes (as in Fig. 3b) varied as a function of cell group (ANOVA, P < 0.009; central cells ratio = 2.74 ± 0.24; medial cells ratio 5.27 ± 0.49; lateral cells ratio 5.14 ± 0.60, same cells as for short-term depression analysis, above). The two edge groups did not differ (two-tailed Bonferroni’s t-test, P = 0.99). Central cell ratios were significantly lower than medial and lateral cell ratios (two-tailed Bonferroni’s t-test, P < 0.009, <0.05, respectively). e, Input resistances of medial edge (n = 16 cells, 14 mice), central (n = 14 cells, 13 mice), and lateral edge cells (n = 15 cells, 13 mice) varied as a function of cell group (P < 0.03, ANOVA). The two edge groups were not different from one another (two-tailed Bonferoni’s t-test, P = 0.99). Central cell input resistance was significantly different from medial, although not from lateral (two-tailed Bonferroni’s t-tests, P < 0.04 and P = 0.074, respectively). f, Central cells of visual TRN had more robust intrinsic bursting than those along the edges. Left, representative stimulus-offset bursts. Right, average number of spikes during bursts was 5.8 ± 0.76 for central cells (n = 19 cells, 15 mice), 0.8 ± 0.30 for medial cells (n = 19 cells, 14 mice), and 1.1 ± 0.50 for lateral cells (n = 14 cells, 12 mice) (P < 0.0001, ANOVA). The two edge groups were not different from one another (two-tailed Bonferoni’s t-test, P = 0.99), but the edge cells discharged far fewer spikes per burst than the central cells (two-tailed Bonferoni’s t-test, P < 0.0001 for both medial vs central cells and lateral vs central cells). For the experiments represented in this figure, the boundaries of the TRN were determined using either tdTomato expression driven by SOM-Cre or PV-Cre, IHC for PV, or brightfield images. Data are mean ± s.e.m.
Extended Data Fig. 10 Organizational and operational scheme for somatosensory circuits of the TRN.
a, Schematic illustrating the organization of the somatosensory sector of the TRN. Excitatory inputs from the primary VP thalamic nucleus innervate the CB-dense central zone of TRN (green cells), whereas higher-order POM inputs innervate the SOM-dense edges (red cells). The inhibitory projections of TRN cells to the VP and POM seem to reciprocate the patterns of their inputs (coloured axons in the schematic). The spatial organization of the neurochemically distinct TRN cell types, and the stark segregation of their primary and higher order connections, suggest a core-shell organization for somatosensory circuits of the TRN. b, Distinct responses of central and edge TRN cells to their thalamic inputs. The spiking responses of central and edge cells are determined by the dynamic synaptic properties of their respective thalamic inputs combined with the intrinsic physiological properties of the TRN cells themselves. The spiking of central cells evoked by VP input is phasic, with strong initial bursts that deeply depress with repetitive stimulation; this is consistent with a core circuit that processes transient sensory signals. By contrast, the spiking responses of edge cells to POM input are more sustained, with initially weaker but more persistent activation; this shell-like circuit is suited to processing temporally distributed higher-order signals.
Supplementary information
Supplementary Information
Supplementary Information Table 1 | Intrinsic physiological properties of central and edge TRN cells. Passive membrane properties, action potential (AP) kinetics, and repetitive firing properties are shown for central and edge neurons of somatosensory TRN (page 1) and visual TRN (page 2). Properties were measured in current-clamp mode, with steady-state Vm set to either -84 mV (“burst mode”, left) or -74 mV (“tonic mode”, right). “Offset bursts” are intrinsic bursts triggered by the offset of hyperpolarizing current pulses, as illustrated in Fig. 3d and Extended Data Fig. 9f. The numbers of cells are shown in parenthesis. T-tests were unpaired and two-tailed. Data are expressed as mean ± SEM.
Rights and permissions
About this article

Cite this article
Martinez-Garcia, R.I., Voelcker, B., Zaltsman, J.B. et al. Two dynamically distinct circuits drive inhibition in the sensory thalamus. Nature 583, 813–818 (2020). https://doi.org/10.1038/s41586-020-2512-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2512-5
This article is cited by
-
Pallidal GABA B receptors: involvement in cortex beta dynamics and thalamic reticular nucleus activity
The Journal of Physiological Sciences (2023)
-
Thalamocortical loops as temporal demodulators across senses
Communications Biology (2023)
-
The endocannabinoid N-arachidonoyl dopamine is critical for hyperalgesia induced by chronic sleep disruption
Nature Communications (2023)
-
Thalamic subnetworks as units of function
Nature Neuroscience (2022)
-
Untangling the cortico-thalamo-cortical loop: cellular pieces of a knotty circuit puzzle
Nature Reviews Neuroscience (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
