
Abstract
An outbreak of coronavirus disease 2019 (COVID-19)1,2,3, caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)4, has spread globally. Countermeasures are needed to treat and prevent further dissemination of the virus. Here we report the isolation of two specific human monoclonal antibodies (termed CA1 and CB6) from a patient convalescing from COVID-19. CA1 and CB6 demonstrated potent SARS-CoV-2-specific neutralization activity in vitro. In addition, CB6 inhibited infection with SARS-CoV-2 in rhesus monkeys in both prophylactic and treatment settings. We also performed structural studies, which revealed that CB6 recognizes an epitope that overlaps with angiotensin-converting enzyme 2 (ACE2)-binding sites in the SARS-CoV-2 receptor-binding domain, and thereby interferes with virus–receptor interactions by both steric hindrance and direct competition for interface residues. Our results suggest that CB6 deserves further study as a candidate for translation to the clinic.
Similar content being viewed by others

Main
Emerging and recurrent pathogens are global challenges for public health5. Cases of a typical pneumonia—officially designated as COVID-19 by the World Health Organization—have been reported since the end of 20191,2,3. The underlying pathogen has been confirmed to be a novel coronavirus, which—on the basis of phylogenetic evidence—is closely related to a bat coronavirus6. The International Committee on Taxonomy of Viruses has recently named the virus SARS-CoV-24, although some virologists have argued that HCoV-19 would be more appropriate7. As of 1 April 2020, there were more than 880,000 confirmed cases of COVID-19 and more than 44,000 COVID-19-related deaths globally (https://covid19.who.int/). No vaccines or drugs have yet been approved for the treatment of COVID-19. However, multiple clinical trials have been launched to evaluate the efficacy and safety of inactivated convalescent plasma in the treatment of COVID-19 (www.ClinicalTrials.gov, identifiers NCT04292340, NCT04327349 and NCT04321421). Preliminary data have suggested a potential benefit from treatment with plasma from convalescent individuals (http://www.nhc.gov.cn/xcs/fkdt/202002/2a1d9d8569c549e7a5d5737751c74480.shtml), which indicates that neutralizing antibodies from convalescent patients could inhibit virus infection and have potential for clinical use. However, the practical limitations of collecting plasma from convalescent individuals, and a lack of suitable risk assessment, make large-scale plasma transfusion programmes a challenge.
SARS-CoV-2 is the third highly pathogenic coronavirus that infects humans to have emerged; it uses the same receptor—angiotensin-converting enzyme 2 (ACE2)—for cell entry as severe acute respiratory syndrome coronavirus (SARS-CoV)6,8. As with other coronaviruses, the SARS-CoV-2 spike (S) protein can be further separated into S1 and S2 subdomains, with S1 binding to the host receptor and S2 mediating membrane fusion9. The receptor-binding domain (RBD) of the SARS-CoV-2 S protein at the S1 C-terminal domain has recently been mapped, and the crystal structure of the SARS-CoV-2 RBD in complex with human ACE2 has been solved10,11,12,13. Previous studies have revealed that a large number of antibodies show neutralization activity by targeting the RBD of either SARS-CoV or Middle East respiratory syndrome coronavirus (MERS-CoV), presumably by disrupting the virus–receptor engagement14,15,16. Thus, we attempted to isolate neutralizing monoclonal antibodies (mAbs) against SARS-CoV-2 from patients convalescing from COVID-19, using a similar strategy to that previously reported17,18.
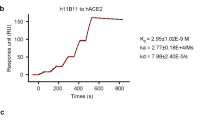
We used a recombinant RBD of the SARS-CoV-2 S protein as the bait to sort specific memory B cells from peripheral blood mononuclear cells of a patient convalescing from COVID-19. The sequences of the variable regions of IgG antibodies in the sorted cells were obtained through 5′ rapid amplification of cDNA ends (RACE) from individual B cells. We identified two mAbs—named CA1 and CB6 (Extended Data Table 1)—as being able to block the binding of soluble SARS-CoV-2 RBD to the ACE2 receptor transiently expressed on HEK293T cells (Fig. 1a). Binding analysis of CA1 and CB6 by fluorescence-activated cell sorting (FACS) revealed that both mAbs can specifically bind to HEK293T cells transfected with SARS-CoV-2 S protein, but not to cells transfected with SARS-CoV S or MERS-CoV S protein (Fig. 1b). We assessed the binding kinetics of CA1 and CB6 using a surface plasmon resonance (SPR) assay. The measured equilibrium constants (KD) of CA1 and CB6 with the SARS-CoV-2 RBD were 4.68 ± 1.64 nM and 2.49 ± 1.65 nM, respectively (Fig. 1c, Extended Data Table 2). Because CA1 and CB6 have distinctive complementarity-determining-region (CDR) sequences (Extended Data Table 1), we examined the potential competitive binding of CA1 and CB6 to the SARS-CoV-2 RBD in an Octet-based binding assay. Recombinant SARS-CoV-2 RBD was first biotinylated and immobilized on a streptavidin biosensor, and then saturated with CB6 or CA1. The addition of CA1 to CB6-saturated probe or CB6 to CA1-saturated probe showed no complementary binding (Extended Data Fig. 1), which indicates that CA1 and CB6 bind to overlapping epitopes on the SARS-CoV-2 RBD.

a, CA1 and CB6 can block SARS-CoV-2 RBD binding to ACE2, as shown in a FACS-based assay. ACE2–GFP fusion protein was transiently expressed on the surface of HEK293T cells, and stained with SARS-CoV-2 RBD protein pre-incubated with isotype-control IgG, CA1 or CB6. Experiments were performed twice, and one set of representative data is displayed. b, Left, HEK293T cells that express SARS-CoV S protein were permeabilized, and stained with isotype-control IgG, CA1, CB6 or the positive-control mAb 80R26. Cells expressing MERS-CoV S (middle) or SARS-CoV-2 S (right) protein were stained with isotype-control IgG, CA1, CB6 or the control mAb 4C214. Experiments were performed twice, and one set of representative data is displayed. hFc, human Fc. c, The binding kinetics of CA1 (left) and CB6 (right) to recombinant SARS-CoV-2 RBD were obtained using the BIAcore 8K system in single-cycle mode. mAbs were captured on the chip, and serial dilutions of RBD were then injected over the chip surface. The KD is labelled accordingly. Values are mean ± s.d. of three independent results.
Next, we investigated the neutralization abilities of CA1 and CB6 against SARS-CoV-2 infection using pseudoviruses that express S antigen of SARS-CoV-2, and then investigated the same abilities using live virus in vitro. Both CA1 and CB6 inhibit pseudovirus transduction into Huh7, Calu-3 and HEK293T cells (Fig. 2a–c). Notably, CB6 had stronger neutralizing activity than CA1 in terms of its 50% neutralization dose (ND50) in all three tested cell lines. Consistently, CB6 also exhibited stronger neutralizing activity than CA1 against live SARS-CoV-2 infection of Vero E6 cells, with observed ND50 values of 0.036 ± 0.007 μg ml−1 for CB6 and 0.38 μg ml−1 for CA1 (Fig. 2d, e). In summary, both mAbs exhibit substantial neutralizing activities against SARS-CoV-2 infection in vitro, although CB6 exhibited greater neutralizing activity than CA1.

a–c, SARS-CoV-2 pseudovirus was incubated with threefold serially diluted CA1 or CB6. The mixtures were then added to Huh7 (a), Calu-3 (b) or HEK293T (c) cells. After a 24-h incubation, neutralization potencies of the mAbs were evaluated in a luciferase assay system. ND50 values were calculated by fitting the luciferase activity from serially diluted antibody to a sigmoidal dose–response curve. The experiments were performed twice and similar results were obtained. One set of representative data from one experiment is shown; data are average values of two replicates. d, Mixtures of live SARS-CoV-2 virus and serially diluted CA1 or CB6 were added to Vero E6 cells. After a 30-h incubation, ND50 was calculated by fitting the viral RNA copies (determined by quantitative PCR with reverse transcription (RT–qPCR)) from serially diluted antibody to a sigmoidal dose–response curve. The experiments for CA1 were performed twice, and similar results were obtained. For CB6, the values shown are the mean ± s.d. of three independent experiments. For both CA1 and CB6, one set of representative data from one experiment is shown; data are average values of two replicates. e, Mixtures of SARS-CoV-2 and serially diluted CA1 or CB6 were added to Vero E6 cells. After a 72-h incubation, ND50 values were calculated by fitting the proportion of cytopathic effect with serially diluted antibody to a sigmoidal dose–response curve. The experiments were independently performed twice and similar results were obtained. One set of representative data is shown.
We further tested CB6 in vivo in a rhesus macaque model of SARS-CoV-2 infection, in both prophylactic and treatment settings. Given the potential risk of an antibody-dependent enhancement effect (as observed in SARS-CoV infection)19,20, we introduced two leucine-to-alanine substitutions at residues 234 and 235 (known as the LALA mutation) of the Fc portion of CB6 (hereafter, CB6(LALA)) to lower the risk of Fc-mediated acute lung injury. In the treatment group, three macaques (6–8 years old) were challenged with 1 × 105 50% tissue culture infectious dose (TCID50) of SARS-CoV-2 via intratracheal incubation. Fifty mg per kg body weight CB6(LALA) was administered at 1 and 3 days post-infection (dpi) intravenously. Macaques in the control group (n = 3) were given an equal volume of phosphate-buffered saline (PBS) at both 1 and 3 dpi. The viral titres from throat swabs were evaluated daily until 7 dpi. In the control group, the viral loads reached peak levels (approximately 106.5 RNA copies per ml) at 4 dpi and then declined naturally (Fig. 3a). By contrast, CB6(LALA) treatment reduced virus titres immediately after administration. Specifically, by 4 dpi, CB6(LALA) had reduced the viral titre by approximately 3 log compared to the control group (Fig. 3a). Additionally, in the prophylactic group of three macaques, a single dose of CB6(LALA) (50 mg kg−1 body weight) before challenge with SARS-CoV-2 protected the macaques from SARS-CoV-2 infection. Only minimal levels of virus were detected in the throat swabs in this group and the peak viral load was no more than 103 RNA copies per ml (Fig. 3a), indicating a strong prophylactic protection effect of CB6(LALA) antibody against infection with SARS-CoV-2.

a, Nine rhesus macaques (6 males and 3 females) were divided into pre-exposure (prophylactic), post-exposure (treatment) and control groups with three macaques (2 males and 1 female) in each group. Before infection, the macaques in the pre-exposure group were infused with 50 mg kg−1 body weight CB6(LALA) intravenously. One day later, all macaques were inoculated with 1 × 105 TCID50 SARS-CoV-2 via intratracheal intubation. The post-exposure group were also infused with 50 mg kg−1 CB6(LALA) on days 1 and 3 after exposure, and the three macaques in the control group were given PBS as a control. Viral RNA loads in throat swabs, determined by RT–qPCR, were monitored for 7 days. Data are average values from 3 macaques for the first 5 days, from 2 macaques at 6 dpi, and from 1 macaque at 7 dpi. To evaluate the viral loads for each macaque at the indicated time point, RT–qPCR were performed with two replicates. b–d, Histopathology and immunohistochemical examination of lung tissues from pre-exposure, post-exposure and control macaques. One macaque from each group was euthanized and necropsied at 5 dpi. Samples for histological examination were stored in formalin for 7 days, embedded in paraffin, sectioned and stained before examination by light microscopy. Haematoxylin and eosin (H&E) sections exhibited the interstitial pneumonia and inflammatory-factor infiltration in tissues. Masson’s trichrome showed lung tissue fibrosis. Scale bar, 200 μm.
In addition to reductions in viral titre, we further investigated whether CB6(LALA) also inhibited the pathological lung damage in the macaques challenged with SARS-CoV-2. One macaque from each group was euthanized and necropsied at 5 dpi. The control macaque displayed interstitial pneumonia characterized by thickened alveolar septa and proliferation and fibrosis of fibroblasts, with intensive infiltration of monocytes and lymphocytes. In some alveolar cavities, cellulose exudation was observed, with the formation of hyaline membrane and pulmonary haemorrhage. There was also clear thrombosis in the pulmonary capillary lumen, with accumulation of necrotic and exfoliated epithelial cells of the bronchioles (Fig. 3b). By contrast, macaques treated prophylactically or after infection displayed limited pathological lung damage. The necropsied macaques from the treatment and prophylactic groups had an overall intact alveolar structure, reduced oedema and no formation of hyaline membrane, with less fibrosis and less leukocyte infiltration compared to the control macaques (Fig. 3c, d). In addition, no serious small bronchi and pulmonary capillary lesions were observed (Fig. 3c, d). Thus, CB6(LALA) inhibited SARS-CoV-2 viral titre and reduced infection-related lung damage, in both prophylactic and treatment settings.
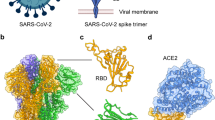
To further investigate the blocking and neutralizing mechanisms of CB6, we prepared a protein complex of recombinant CB6 Fab and SARS-CoV-2 RBD for crystal screening (Extended Data Fig. 2). The 3D structure of the CB6 Fab–SARS-CoV-2 RBD complex was determined by X-ray crystallography at an overall resolution of 2.9 Å (Extended Data Table 3). Overall, CB6 binds to SARS-CoV-2 RBD with a buried surface of 1,088 Å2 (Fig. 4a). Specifically, the CB6 VH segment dominates the interaction with the SARS-CoV-2 RBD by all three CDRs, forming concentrated polar contacts (Fig. 4b, Extended Data Table 4) and hydrophobic interactions (Fig. 4c, Extended Data Table 4), whereas VL has limited contacts with the LCDR1 and LCDR3 loops (Fig. 4d, Extended Data Table 4).

a, The structure of CB6 bound to SARS-CoV-2 RBD. The SARS-CoV-2 RBD is coloured cyan (core subdomain) and yellow (external subdomain). The variable fragment of CB6 is shown with HCDR1, HCDR2 and HCDR3 loops from the VH domain (purple) (which are coloured blue, marine and purple, respectively), and the LCDR1, LCDR2 and LCDR3 loops from the VL domain (pink) (which are coloured hot pink). b, c, Both hydrophilic interactions (b) and hydrophobic interactions (c) between the CB6 heavy chain and SARS-CoV-2 RBD are displayed. d, The binding details between the CB6 light chain and SARS-CoV-2 RBD are presented. The hydrogen bonds are shown as dashed black lines. e, Superimposition of CB6–SARS-CoV-2 RBD complex and ACE2–SARS-CoV-2 RBD complex (PDB code 6LZG) reveals steric competition between CB6 and ACE2 for RBD binding. The CB6–SARS-CoV-2 RBD structure is superimposed on ACE2–SARS-CoV-2 RBD to demonstrate steric hindrance. ACE2 is shown as a cartoon (grey). f, Competitive binding surfaces of CB6 with ACE2 on SARS-CoV-2 RBD. The SARS-CoV-2 RBD binding surface to ACE2 and CB6 is shown. The residues bound by both CB6 and ACE2 are coloured magenta. The residues in contact with ACE2 alone are coloured grey, and the residues in contact with CB6 alone are coloured blue. The amino acids on the SARS-CoV-2 RBD interface that contact CB6 or ACE2 are labelled.
To analyse the mechanisms by which CB6 blocks the interaction between SARS-CoV-2 RBD and ACE2, we superimposed the structure of the CB6 Fab–SARS-CoV-2 RBD complex with that of ACE2–SARS-CoV-2 RBD (Protein Data Bank (PDB) code 6LZG) (Fig. 4e). The superimposition of the RBD in its CB6-bound form with its ACE2-bound form resulted in Cα root mean squared deviation of 0.282 Å (for 169 atoms), indicating that the binding of CB6 did not induce substantial conformational changes in the SARS-CoV-2 RBD. The binding of CB6 induced steric hindrance to the binding of ACE2, which is mediated by both the VH and VL of CB6 (Fig. 4e). The whole of the CB6 light chain—as well as most of the heavy chain—has structure clashes with the receptor. There are also substantial overlapping binding areas between CB6 and ACE2 on the RBD (Fig. 4f). In summary, the blocking mechanisms of CB6 depends on both its VH and its VL, which provide steric hindrance and direct competition for interface residues to abrogate the binding of ACE2 to the SARS-CoV-2 RBD.
Previous studies focusing on SARS-CoV infection in vitro and in mouse models19,20 indicate the potential risk of antibody-dependent enhancement hindering the ability of antibody-based therapies to control inflammation in the lung and other organs. On the contrary, antibody-dependent enhancement may lead to acute respiratory injury, acute respiratory distress syndrome and other observed inflammation-based sequelae. Given the close phylogenetic relationship between SARS-CoV and SARS-CoV-2, the risk of antibody-dependent enhancement needs to be mitigated. To solve this problem, we introduced the LALA mutations to the Fc portion of CB6 molecule to eliminate the antibody-dependent cellular cytotoxicity effect. Subsequently, CB6(LALA) showed a protective effect in a rhesus monkey model of SARS-CoV-2 infection, and did not exacerbate tissue damage induced by the infection. To our knowledge, this represents the first report to evaluate the function of neutralizing mAbs against SARS-CoV-2 using nonhuman primates, and can be used to inform the future design of dosing regimens in clinical trials.
Currently, no drugs or vaccines have been approved as a treatment for COVID-19, highlighting the unmet medical need to develop therapeutic agents. Neutralizing mAbs are promising candidates to combat emerging viruses. Using Ebola virus as an example, ZMapp—which comprises three chimeric mAbs—was used to successfully treat two individuals with Ebola virus disease in 201421; MAb114 has also shown notable treatment benefits, reducing the mortality rate of this disease from about 67% to 34% for all patients, and to 9.9% for patients with low viral loads22. Recently, MAb114 has been designated a breakthrough therapy for the treatment of Ebola virus disease. The neutralizing activity in vitro, and protection efficacy in vivo, of CB6 against SARS-CoV-2 is good and is likely to be comparable to that of MAb114 against Ebola virus23,24.
In addition to post-exposure treatments, neutralizing mAbs can also be used in a prophylactic setting. The first mAb approved by the US Food and Drug Administration against infectious disease is palivizumab, which is indicated for the prophylaxis of pre-term infants and children at a high risk of infection with respiratory syncytia virus. The current pandemic of COVID-19 poses great risks to front-line healthcare workers, elderly patients and patients with pre-existing conditions. In the current situation (when vaccines are not available), preventative treatments are greatly needed. Our in vivo protection data from pre-exposure settings in macaques indicate that CB6 is a promising candidate for development as a prophylaxis for COVID-19.
Our structural analysis has revealed the molecular basis for the neutralizing activity of CB6. Notably, the interacting epitopes on the SARS-CoV-2 RBD for CB6 largely overlap with the binding epitopes of ACE2, which indicates that this mAb has a strong resistance potential to SARS-CoV-2 mutation variants. In the 157 viral genomes for SARS-CoV-2 that are currently deposited in the National Center for Biotechnology Information (NCBI) databank (as of 1 April 2020), two substitutions in the RBD region have been observed (G476S and V483A) (Extended Data Fig. 3). Although G476S is located within the binding interface with CB6, this residue has limited contribution to the antibody–antigen interaction and is thus unlikely to influence the binding of CB6.
During the revision of this Article, CR3022—a neutralizing antibody against SARS-CoV that was previously isolated from a patient convalescing from SARS—was found to be cross-reactive with the SARS-CoV-2 RBD. Crystal-structure analysis revealed that CR3022 targets a highly conserved epitope, distal from the receptor-binding site and does not neutralize infection with SARS-CoV-225. Superimposition of the SARS-CoV-2 RBD in complex with either CB6 or CR3022 (PDB code 6W41) indicates that it is unlikely that CB6 will have an effect on the binding of CR3022 (and vice versa) (Extended Data Fig. 4). Nevertheless, the reported molecular basis for the binding of CR3022 with SARS-CoV-2 RBD deepened our understanding of antibody recognition of SARS-CoV-2.
In conclusion, our data indicate that CB6—a neutralizing mAb isolated from a patient convalescing from COVID-19—could be a potential therapeutic agent for the treatment of COVID-19 and that it deserves further translational development.
Methods
Study design
This study was designed to isolate SARS-CoV-2-specific mAbs from the blood of patients recovering from infections with SARS-CoV-2; characterize the properties of mAbs, including blocking function, binding affinity, neutralization against SARS-CoV-2 and neutralizing mechanism; and then to test the best-performing mAb for therapeutic efficacy in rhesus macaques.
Peripheral blood mononuclear cells were isolated, following the manufacturer’s instructions (GE Healthcare), before being subjected to cell sorting as previously reported17. In brief, peripheral blood mononuclear cells were incubated with His-tagged SARS-CoV-2 RBD at 100 nM before staining with anti-CD3, anti-CD16, anti-CD235a, anti-CD19, anti-CD38, anti-CD27 and anti-His. Antigen-specific memory B cells were identified by the following markers: CD3−, CD16−, CD235a−, CD38−, CD19+, CD27+, IgG+ and His+, and sorted into 96-well PCR plates with single cell per well. Flow cytometric analysis and cell sorting were performed on a BD FACSAria ΙΙΙ flow cytometer (BD Biosciences) and the data were analysed using FlowJo. The genes that encode the immunoglobulin VH and VL chains were amplified by 5′ RACE and nested PCR. The variable regions of these genes were then linked with the coding sequences for human IgG1 constant region to generate full-length mAbs, which were then expressed and purified under good laboratory practice conditions.
Eleven mAbs were generated with this method. Through blocking assays for SARS-CoV-2 RBD and ACE2 by FACS, two mAbs were analysed for several properties: (a) binding specificity for the S protein of MERS-CoV, SARS-CoV and SARS-CoV-2; (b) the competition assay for mAbs binding using Octet; and (c) the ability of the mAbs to neutralize the SARS-CoV-2 in Vero E6 cells. The results suggested that these two mAbs specifically bound and potently neutralized SARS-CoV-2 in vitro. Using a rhesus macaque model of SARS-CoV-2 infection, we tested the protective and curative efficacy of CB6. We equally divided the nine macaques into pre-exposure, post-exposure and negative-control groups. Viral loads from throat swabs were detected in the 3 groups over a consecutive period of 7 days. The macaques were euthanized and necropsied on 5 dpi to evaluate the therapeutic effect of CB6. Then, we investigated the neutralizing mechanism of the mAb through structural analysis of CB6–SARS-CoV-2 RBD. The high-resolution structure of this complex clarified that the binding surface of CB6 on the SARS-CoV-2 RBD overlapped with that of ACE2, indicating a substantial stereospecific hindrance to ACE2 contacting RBD.
Cells and viruses
HEK293T (ATCC, CRL-3216) cells, Huh7 (Institute of Basic Medical Sciences CAMS, 3111C0001CCC000679) cells, Calu-3 (ATCC, HTB-55) cells and Vero E6 (ATCC, CRL-1586) cells were cultured at 37 °C in Dulbecco’s Modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS). The cells have not been identified or tested for mycoplasma contamination. The SARS-CoV-2 strain BetaCoV/Wuhan/IVDC-HB-envF13/2020 (accession identifier: EPI_ISL_408511) was isolated by the National Institute for Virus Disease Control and Prevention (Chinese Center for Disease Control and Prevention). Vero E6 cells were applied to the reproduction of SARS-CoV-2 stocks.
Gene construction
The recombinant proteins SARS-CoV-2 RBD and ACE2 were used in FACS, surface plasmon resonance (SPR), biolayer interferometry (BLI) and crystal screening assays. The coding sequences of SARS-CoV-2 RBD tagged with a C-terminal 6× His tag was cloned into the pCAGGS expression vector using the EcoRI and XhoI restriction sites. The coding sequences of ACE2 (residues 19–615, accession number BAJ21180) was cloned into the baculovirus transfer vector pFastbac1 (Invitrogen) containing an N-terminal gp67 signal peptide and a C-terminal 6× His tag. The full-length coding region of the SARS-CoV S (accession number NC_004718), MERS-CoV S (accession number JX869050) or SARS-CoV-2 S protein with a C-terminal Flag tag was cloned into the pCAGGS vector using the EcoRI and SmaI restriction sites (pCAGGS-SARS-CoV-S-Flag, pCAGGS-MERS-CoV-S-Flag and pCAGGS-SARS-CoV-2-S-Flag, respectively). The pEGFP-N1-ACE2 plasmid was constructed by cloning the coding region of ACE2 into pEGFP-N1 using restriction enzymes XhoI and SmaI.
Protein expression and purification
The SARS-CoV-2 RBD recombinant protein was expressed in HEK293T cells. The pCAGGS plasmid containing the SARS-CoV-2 RBD coding sequences was transiently transfected into cells. After 3 days, the supernatant was collected and soluble protein was purified by Ni affinity chromatography using a HisTrap HP 5-ml column (GE Healthcare). The sample was further purified via gel filtration chromatography with a Superdex 200 column (GE Healthcare) in a buffer composed of 20 mM Tris-HCl (pH 8.0) and 150 mM NaCl. The Bac-to-Bac baculovirus expression system (Invitrogen) was used to express the ACE2 for Octet analysis. The constructed pFastbac1 vectors were transformed into DH10Bac competent cells to generate recombinant bacmids. Transfection of bacmids and virus amplification were conducted in Sf9 cells, and Hi5 cells were used for protein expression. The soluble ACE2 protein was purified by the same process as described for the purification of SARS-CoV-2 RBD.
FACS assay
The activity of mAbs to block the binding between the SARS-CoV-2 RBD and ACE2 was assessed by FACS. HEK293T cells were transiently transfected with the pEGFP-N1-ACE2 expression plasmid for 24 h. The SARS-CoV-2 RBD protein at a concentration of 200 ng/ml was mixed with the mAbs or isotype IgG at a molar ratio of 1:10 and incubated at 4 °C for 1 h. Then mixtures were added to 5 × 105 HEK293T cells expressing ACE2 and incubated at 4 °C for another hour. After washing with PBS three times, the cells were stained with anti-His APC-conjugated antibody for another 30 min and analysed using flow cytometry (BD FACSCalibur).
To test the binding specificity, pCAGGS-SARS-CoV-S-Flag, pCAGGS-MERS-CoV-S-Flag and pCAGGS-SARS-CoV-2-S-Flag plasmids were transfected into HEK293T cells using PEI (Alfa) according to the manufacturer’s instructions. Four hundred thousand cells were collected 24 h after transfection, and suspended in permeabilization solution (BD) on ice for 20 min. After washing with PBS twice, the cells were incubated with 20 μg/ml CA1, CB6, 80R, 4C2 or isotype IgG at room temperature for 1 h, followed by washing with PBS twice and further incubation with anti-human IgG APC-conjugated antibody (with the exception of 4C2, which used anti-mouse IgG APC-conjugated antibody as the second antibody) and anti-Flag FITC-conjugated antibodies. After washing, the cells were analysed using a BD FACSCalibur. All of these data were analysed using FlowJo.
SPR
SPR measurements were done at room temperature using a BIAcore 8K system with Protein A biosensor chips (GE Healthcare). For all measurements, a buffer consisting of 150 mM NaCl, 10 mM HEPES, pH 7.4 and 0.005% (v/v) Tween-20 was used as running buffer, and all proteins were exchanged into this buffer in advance through gel filtration. The blank channel of the chip served as the negative control. Two mAbs were captured on the chip at 6,000 response units. Gradient concentrations of SARS-CoV-2 RBD (from 50 nM to 3.125 nM with twofold dilution) then flowed over the chip surface. After each cycle, the sensor was regenerated with Gly-HCl (pH 1.7). The affinity was calculated using a 1:1 (Langmuir) binding fit model with BIAevaluation software.
BLI
The competitive binding of CA1, CB6 and ACE2 were measured by BLI using the Octet RED96 system (FortéBio). All experiments were performed at 25 °C in a buffer containing 20 mM Tris-HCl and 150 mM NaCl, pH 7.0. Streptavidin biosensors were pre-equilibrated in the buffer for at least 10 min before use in experiments. Biotinylated SARS-CoV-2 RBD was loaded onto streptavidin biosensors for 300 s. For determining competitive characteristics, sensors were immersed with the first protein (or buffer as a control) for 240 s, and then the second protein for another 240 s. We analysed the data using FortéBio Data Analysis.
Neutralization assay
SARS-CoV-2 pseudovirus preparation was performed as previously described27. In brief, the plasmids of 15 μg pCAGGS-SARS-CoV-2-S and 15 μg pNL4-3.luc.RE were cotransfected into HEK293T cells. Forty-eight h later, the supernatant was sequentially collected, centrifuged and filtered through a 0.22 μm sterilized membrane. The TCID50 was determined by the transduction of pseudovirus into Huh7 cells. For the neutralization assay, 104 cells per well in 100 μl were seeded in the 96-well plates 16 h before infection. Fifty μl supernatant containing 103 TCID50 pseudovirus was incubated with equal volume of threefold serially diluted antibodies for 1 h at 37 °C. Both mAbs were tested in the concentrations ranging from 5.08 ng/ml to 100 μg/ml in the context of Huh7 cells and Calu-3 cells; for HEK293T cells, 15.24 ng/ml to 300 μg/ml mAb concentrations were used. The mixtures of pseudoviruses and mAbs were then added to Huh7 cells, with two replicates. After a 4-h incubation, the medium was replaced with DMEM containing 10% fetal bovine serum, and the samples were incubated for an additional 24 h at 37 °C. Cell lysates were transferred (10 μl per well) into luminometer plates (Microfluor 96-well plates), and luciferase substrate (40 μl per well)—included in luciferase assay system—was added. Luciferase activity was measured using a GloMax 96 Microplate luminometer (Promega). The 50% neutralization dose was calculated using GraphPad Prism 6.0.
An infectious SARS-CoV-2 neutralization assay was performed. Vero E6 cells were seeded at 8 × 104 per well in a 24-well culture plates at 37 °C for 24 h before being used. To reach a multiplicity of infection of 0.005, 1 ml of diluted SARS-CoV-2 virus and twofold serially diluted CA1 (from 5.0 μg/ml to 0.15 μg/ml) or CB6 (from 1.0 μg/ml to 0.015 μg/ml) mAbs were mixed in the medium containing 2% FBS. Meanwhile, a control group (without antibody) was set up, and the culture supernatant containing virus was collected 30 h post infection. One hundred and forty μl of the supernatant at each well was taken and RNA extraction was carried out according to the instructions of QIAamp viral RNA mini kit. RT–qPCR detection was carried out with one-step TB Green PrimeScript PLUS RT–PCR Kit (Perfect Real Time), and the primer was RBD-qF1: 5′-CAATGGTTTAACAGGCACAGG-3′, RBD-qR1: 5′-CTCAAGTGTCTGTGGATCACG-3′. The total volume of the reaction system was 20 μl. The reaction conditions were: reverse transcription at 42 °C for 5 min, pre-denaturation at 95 °C for 10 s, 40 cycles including denaturation at 95 °C for 10 s, and annealing and extension at 60 °C for 30 s. Another neutralization assay was based on cytopathic effect (CPE). Serial twofold dilutions of 50 μl of antibody CA1 (from 97.6 ng/ml to 200 μg/ml) or CB6 (from 48.8 ng/ml to 100 μg/ml) were prepared in a 96-well tissue culture plate in MEM medium. An equal volume of SARS-CoV-2 virus containing 100 TCID50 was added, and the antibody–virus mixture was incubated at 37 °C for 1 h. The antibody–virus mixture was then transferred into a 96-well microtitre plate containing an equal volume of confluent Vero E6 cells with 8 repeats and incubated at 37 °C for 3 days. Cells infected with 100 TCID50 of SARS-CoV-2 and without the virus were applied as positive and uninfected controls, respectively. The CPE in each well was observed daily and recorded on day 3 after infection. A virus back-titration was performed to assess the correct virus titre used in each experiment. The ND50 was calculated using GraphPad Prism 6.0. All experiments were followed the standard operating procedures of the approved Biosafety Level 3 facility.
Human samples
The human samples were obtained according to procedures approved by the Chinese Academy of Sciences, and complied with all relevant ethical regulations regarding human research. The blood was taken from a patient convalescing from COVID-19 after they had signed the informed consent form.
Macaque experiments
All macaque experiments were carried out according to the procedures approved by the Chinese Academy of Sciences and complied with all relevant ethical regulations regarding animal research.
Nine rhesus macaques (6–8 years old, 3 females and 6 males) were inoculated with 1 × 105 TCID50 SARS-CoV-2 via intratracheal routes. The rhesus macaques were divided into three groups: a control group (one female and two males), a pre-exposure group (one female and two males) and a post-exposure group (one female and two males). The rhesus macaques in the control group were injected with PBS. The pre-exposure group were given 50 mg/kg CB6 intravenously one day before viral challenge, and the post-exposure group were injected with the same amount of antibody on days 1 and 3 after exposure. In addition to routine clinical observation, oropharyngeal swabs were collected for seven days. Swabs were placed into 1 ml of DMEM after collection. Viral RNA was extracted using the QIAamp Viral RNA Mini Kit (Qiagen), according to the manufacturer’s instructions, and was eluted in 50 μl of elution buffer and used as the template for RT–qPCR. The pairs of primers were used targeting the S gene: RBD-qF1: 5′-CAATGGTTTAACAGGCACAGG-3′; RBD-qR1: 5′-CTCAAGTGTCTGTGGATCACG-3′. Two μl RNA was used to verify the RNA quantity by HiScript II One Step qRT–PCR SYBR Green Kit (Vazyme Biotech) according to the manufacturer’s instructions. The amplification was performed as followed: 50 °C for 3 min, 95 °C for 30 s followed by 40 cycles consisting of 95 °C for 10 s, 60 °C for 30 s, and a default melting curve step in an ABI StepOne machine.
Histopathology and immunohistochemistry
Macaque necropsies were performed according to a standard protocol. Samples for histological examination were stored in 10% neutral-buffered formalin for 7 days, embedded in paraffin, sectioned and stained with haematoxylin and eosin or Masson’s trichrome before examination by light microscopy.
Crystal screening and structure determination
The SARS-CoV-2 RBD protein and CB6 Fab fragment were mixed at a molar ration of 1:1. The mixture was incubated on ice for 1 h and further purified by Superdex-200 column (GE Healthcare). Five mg/ml and 10 mg/ml of stable complex formed by SARS-CoV-2 RBD and CB6 Fab proteins, respectively, were used for crystal screening by vapour-diffusion sitting-drop method at 18 °C. Diffracting crystals were obtained in a concentration of 5 mg/ml at the mother liquid containing 24% w/v PEG 1500, 20% v/v glycerol.
For data collection, the crystals were cryo-protected by briefly soaking in reservoir solution supplemented with 20% (v/v) glycerol before flash-cooling in liquid nitrogen. Diffraction data were collected at Shanghai Synchrotron Radiation Facility BL17U1 (wavelength, 0.97919 Å) at 100 K. The dataset was processed with XDS software (from W. Kabsch, http://xds.mpimf-heidelberg.mpg.de/). The structure of the SARS-CoV-2 RBD–CB6 complex was determined by the molecular replacement method using Phaser28 with the previously reported SARS-CoV-2 RBD structure (PDB code 6LZG) and Fab structure (PDB code 4TSA). The atomic models were completed with Coot29 and refined with phenix.refine in Phenix30, The Ramachandran plot determined by MolProbity indicates that 96.3% in the most-favoured region, 3.53% in the allowed region and 0.16% in the disallowed region31. Data collection, processing, and refinement statistics are summarized in Extended Data Table 3. All structural figures were generated using Pymol software (http://www.pymol.org).
Sequence alignments
The GenBank accession number of the sequences used for analysing the conservation of epitopes among SARS-CoV-2 are the following: MT263459, MT262993, MT262907, MT262915, MT262909, MT262916, MT262911, MT262908, MT262913, MT262912, MT262914, MT262906, MT262899, MT262898, MT262897, MT262904, MT262905, MT262896, MT262903, MT262902, MT262901, MT262900, MT262910, NC_045512, MT263396, MT263445, MT263421, MT258381, MT263391, MT263446, MT259269, MT263395, MT263468, MT263381, MT263074, MT258379, MT259271, MT263458, MT263429, MT258380, MT263382, MT259228, MT259227, MT259231, MT259226, MT259254, MT258377, MT259248, MT258383, MT263435, MT263398, MT258382, MT259277, MT263392, MT263418, MT259236, MT263406, MT263417, MT263436, MT263443, MT259256, MT263452, MT259264, MT263420, MT259281, MT263450, MT263402, MT263433, MT263413, MT263404, MT259260, MT263437, MT263438, MT263403, MT263399, MT263464, MT259261, MT263439, MT259237, MT263410, MT259251, MT263444, MT263465, MT263447, MT263454, MT263463, MT263431, MT263424, MT263432, MT263416, MT259282, MT263419, MT263467, MT263440, MT263411, MT256917, MT256918, MT256924, MT263405, MT263408, MT263386, MT263469, MT263400, MT263425, MT263387, MT259285, MT263430, MT263449, MT263412, MT263414, MT263462, MT263434, MT263457, MT263461, MT263448, MT263423, MT263422, MT263415, MT259249, MT263442, MT259245, MT263427, MT263384, MT263441, MT263383, MT259243, MT263426, MT263388, MT263428, MT263455, MT263456, MT263453, MT263451, MT259235, MT263460, MT263390, MT263394, MT263466, MT259250, MT259280, MT259258, MT259239, MT259240, MT263407, MT263385, MT263397, MT263409, MT263393, MT263401, MT259279, MT259284, MT263389, MT259270, MT259259, MT259242 and MT259238. The sequence logos were generated using the web server of Weblogo (http://weblogo.berkeley.edu/logo.cgi).
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
Further information and requests for resources and reagents should be directed to J.Y. The accession number for the atomic coordinates and diffraction data reported in this study is PDB code 7C01. The sequences of CA1 and CB6 mAbs have been deposited in GenBank with the accession codes MT470194–MT470197. Source data are provided with this paper.
References
Tan, W. et al. Notes from the field: a novel coronavirus genome identified in a cluster of pneumonia cases – Wuhan, China 2019–2020. China CDC Weekly 2, 61–62 (2020).
The 2019-nCoV Outbreak Joint Field Epidemiology Investigation Team & Li, Q. Notes from the field: an outbreak of NCIP (2019-nCoV) infection in China – Wuhan, Hubei province, 2019–2020. China CDC Weekly 2, 79–80 (2020).
Wang, C., Horby, P. W., Hayden, F. G. & Gao, G. F. A novel coronavirus outbreak of global health concern. Lancet 395, 470–473 (2020).
Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 5, 536–544 (2020).
Gao, G. F. From “A”IV to “Z”IKV: attacks from emerging and re-emerging pathogens. Cell 172, 1157–1159 (2018).
Zhou, P. et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270–273 (2020).
Jiang, S. et al. A distinct name is needed for the new coronavirus. Lancet 395, 949 (2020).
Hoffmann, M. et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280.e8 (2020).
Lu, G., Wang, Q. & Gao, G. F. Bat-to-human: spike features determining ‘host jump’ of coronaviruses SARS-CoV, MERS-CoV, and beyond. Trends Microbiol. 23, 468–478 (2015).
Wang, Q. et al. Structural and functional basis of SARS-CoV-2 entry by using human ACE2. Cell 181, 894–904.e9 (2020).
Shang, J. et al. Structural basis of receptor recognition by SARS-CoV-2. Nature 581, 221–224 (2020).
Lan, J. et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 581, 215–220 (2020).
Yan, R. et al. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 367, 1444–1448 (2020).
Li, Y. et al. A humanized neutralizing antibody against MERS-CoV targeting the receptor-binding domain of the spike protein. Cell Res. 25, 1237–1249 (2015).
Du, L. et al. The spike protein of SARS-CoV—a target for vaccine and therapeutic development. Nat. Rev. Microbiol. 7, 226–236 (2009).
Wang, Q., Wong, G., Lu, G., Yan, J. & Gao, G. F. MERS-CoV spike protein: targets for vaccines and therapeutics. Antiviral Res. 133, 165–177 (2016).
Wang, Q. et al. Molecular determinants of human neutralizing antibodies isolated from a patient infected with Zika virus. Sci. Transl. Med. 8, 369ra179 (2016).
Wang, Q. et al. Neutralization mechanism of human monoclonal antibodies against Rift Valley fever virus. Nat. Microbiol. 4, 1231–1241 (2019).
Channappanavar, R. et al. Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe 19, 181–193 (2016).
Liu, L. et al. Anti-spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection. JCI Insight 4, e123158 (2019).
Lyon, G. M. et al. Clinical care of two patients with Ebola virus disease in the United States. N. Engl. J. Med. 371, 2402–2409 (2014).
Mulangu, S. et al. A randomized, controlled trial of Ebola virus disease therapeutics. N. Engl. J. Med. 381, 2293–2303 (2019).
Gaudinski, M. R. et al. Safety, tolerability, pharmacokinetics, and immunogenicity of the therapeutic monoclonal antibody mAb114 targeting Ebola virus glycoprotein (VRC 608): an open-label phase 1 study. Lancet 393, 889–898 (2019).
Corti, D. et al. Protective monotherapy against lethal Ebola virus infection by a potently neutralizing antibody. Science 351, 1339–1342 (2016).
Yuan, M. et al. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 368, 630–633 (2020).
Sui, J. et al. Potent neutralization of severe acute respiratory syndrome (SARS) coronavirus by a human mAb to S1 protein that blocks receptor association. Proc. Natl Acad. Sci. USA 101, 2536–2541 (2004).
Du, L., Zhang, X., Liu, J. & Jiang, S. Protocol for recombinant RBD-based SARS vaccines: protein preparation, animal vaccination and neutralization detection. J. Vis. Exp. 51, 2444 (2011).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D 66, 213–221 (2010).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D 66, 12–21 (2010).
Acknowledgements
We thank S. Yao for his help in revising the manuscript; the staff of beamline BL17U1 at the Shanghai Synchrotron Radiation Facility for assistance during data collection; Z. Fan for her technical help in performing BLI and SPR; all colleagues of the National Biosafety Laboratory for their support during this study; Y. Yao and Y. Peng for supporting all the experiments in nonhuman primates; and F. Wang, Y. Hu, Y. Jia, S. Ren, L. Wu and S. Tan for their help in performing the experiments. This work was supported by the Strategic Priority Research Program of CAS (XDB29040201), the National Major Science and Technology Major Project (2018ZX09711003), the National Key Research and Development Program of China (2020YFC0841400) and the National Natural Science Foundation of China (81922044 and 81973228). Q.W. is supported by the Youth Innovation Promotion Association CAS (2018119). G.F.G and J.Y are supported by the foundation of the NSFC Innovative Research Group (81621091).
Author information
Authors and Affiliations
Contributions
Q.W., G.F.G. and J.Y. initiated and coordinated the project. R.S., F.-S.W., Q.W., G.F.G., Z.Y. and J.Y. designed the experiments. Q.W. performed the cell sorting with the help of Z.C., J.S. and Z.T. R.S., X.D., T.S. and C.H. sequenced the antibodies. R.S. and X.D. conducted the SPR and Octet analysis. W.H. and L.W. prepared the pseudotype SARS-CoV-2 virus. T.S. evaluated the neutralizing potency using pseudovirus. Y.Z. and P.L. evaluated the neutralizing potency using authentic virus, with the help from B.Z., W.J.L. and G.W. C.S. conducted the animal experiments. Y.Z. and Z.Y. performed histopathology and immunohistochemistry assays with the assistance from G.G. and X.H. X.D., X.B. and C.H. grew the complex crystals. J.Q. collected the diffraction data and determined the complex structure with the help from L.W. R.S., H.F., F.-S.W., Q.W., G.F.G. and J.Y. analysed the data and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
R.S., Q.W., G.F.G. and J.Y. are listed as inventors on pending patent applications for CA1 and CB6. The pending patent of CB6 has been licensed. The other authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Analysis of direct competitive binding characteristics of CA1, CB6 and ACE2.
a, b, Competition between CA1 and CB6 (a) or either mAb and ACE2 (b) for binding to the SARS-CoV-2 RBD. Octet sensors immobilized with the SARS-CoV-2 RBD were first saturated with one mAb or the kinetic buffer, and then exposed to the other mAb or ACE2 protein. The experiments were independently performed twice and similar results were obtained. The binding profiles of one experiment are shown.
Extended Data Fig. 2 Preparation of the CB6 Fab–SARS-CoV-2 RBD protein complex.
a, Gel filtration profiles of CB6 Fab (red), SARS-CoV-2 RBD (blue) and the complex of the two (purple) were analysed by size-exclusion chromatography as indicated. The CB6 Fab forms a stable complex with the SARS-CoV-2 RBD. The experiments were independently performed twice and similar results were obtained. b, The SDS–PAGE analyses are shown in reducing (+DTT) conditions. The CB6 Fab was depolymerized into VH and VL fragments (25 kDa). The SARS-CoV-2 RBD exhibited a diffuse band, owing due to glycosylation (38 kDa). For gel source data, see Supplementary Fig. 1.
Extended Data Fig. 3 Sequence alignment of RBD among strains of SARS-CoV-2.
We obtained 157 full-length sequences of the SARS-CoV-2 RBD by searching the NCBI databank. The sequence logos were generated using the Weblogo web server (http://weblogo.berkeley.edu/logo.cgi). Four strains isolated from human patients with COVID-19 in the USA carry G476S, and eight strains carry V483A; further details are provided in Methods.
Extended Data Fig. 4 Comparison of the binding epitopes of CB6 and CR3022.
Superimposition of the CB6–SARS-CoV-2 RBD and CR3022–SARS-CoV-2 RBD (PDB code 6W41) complexes reveals no epitope overlapping between CB6 and CR3022. The SARS-CoV-2 RBD is shown as a cartoon (yellow). The CB6 heavy chain and light chain are coloured grey and pink, respectively. The CR3022 heavy chain and light chain are coloured light blue and cyan, respectively.
Supplementary information
Supplementary Figure 1
| Raw gel-picture of CB6/RBD complex preparation The SDS-PAGE analyses are shown in reducing (+DTT) conditions. CB6-Fab is depolymerized into VH and VL fragments (25 kD). SARS-CoV-2 RBD exhibited a diffuse band due to glycosylation (38 kD). The right panel is not related to this work.
Rights and permissions
About this article

Cite this article
Shi, R., Shan, C., Duan, X. et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 584, 120–124 (2020). https://doi.org/10.1038/s41586-020-2381-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2381-y
This article is cited by
-
Single-domain antibodies against SARS-CoV-2 RBD from a two-stage phage screening of universal and focused synthetic libraries
BMC Infectious Diseases (2024)
-
SARS-CoV-2 immunity in animal models
Cellular & Molecular Immunology (2024)
-
Potent and broadly neutralizing antibodies against sarbecoviruses induced by sequential COVID-19 vaccination
Cell Discovery (2024)
-
An mRNA vaccine encoding the SARS-CoV-2 receptor-binding domain protects mice from various Omicron variants
npj Vaccines (2024)
-
A human antibody derived from original SARS-CoV-2 infection effectively neutralizes omicron
Advanced Biotechnology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
