
Abstract
Autonomic nerve fibres in the tumour microenvironment regulate cancer initiation and dissemination, but how nerves emerge in tumours is currently unknown. Here we show that neural progenitors from the central nervous system that express doublecortin (DCX+) infiltrate prostate tumours and metastases, in which they initiate neurogenesis. In mouse models of prostate cancer, oscillations of DCX+ neural progenitors in the subventricular zone—a neurogenic area of the central nervous system—are associated with disruption of the blood–brain barrier, and with the egress of DCX+ cells into the circulation. These cells then infiltrate and reside in the tumour, and can generate new adrenergic neurons. Selective genetic depletion of DCX+ cells inhibits the early phases of tumour development in our mouse models of prostate cancer, whereas transplantation of DCX+ neural progenitors promotes tumour growth and metastasis. In humans, the density of DCX+ neural progenitors is strongly associated with the aggressiveness and recurrence of prostate adenocarcinoma. These results reveal a unique crosstalk between the central nervous system and prostate tumours, and indicate neural targets for the treatment of cancer.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Change history
08 January 2020
An Amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Eichmann, A. & Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harb. Perspect. Med. 3, a006551 (2013).
Kumar, A. & Brockes, J. P. Nerve dependence in tissue, organ, and appendage regeneration. Trends Neurosci. 35, 691–699 (2012).
Ming, G. L. & Song, H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron 70, 687–702 (2011).
Tanaka, E. M. & Ferretti, P. Considering the evolution of regeneration in the central nervous system. Nat. Rev. Neurosci. 10, 713–723 (2009).
Magnon, C. et al. Autonomic nerve development contributes to prostate cancer progression. Science 341, 1236361 (2013).
Zhao, C. M. et al. Denervation suppresses gastric tumorigenesis. Sci. Transl. Med. 6, 250ra115 (2014).
Pundavela, J. et al. Nerve fibers infiltrate the tumor microenvironment and are associated with nerve growth factor production and lymph node invasion in breast cancer. Mol. Oncol. 9, 1626–1635 (2015).
Dobrenis, K., Gauthier, L. R., Barroca, V. & Magnon, C. Granulocyte colony-stimulating factor off-target effect on nerve outgrowth promotes prostate cancer development. Int. J. Cancer 136, 982–988 (2015).
Hayakawa, Y. et al. Nerve growth factor promotes gastric tumorigenesis through aberrant cholinergic signaling. Cancer Cell 31, 21–34 (2017).
Stopczynski, R. E. et al. Neuroplastic changes occur early in the development of pancreatic ductal adenocarcinoma. Cancer Res. 74, 1718–1727 (2014).
Peterson, S. C. et al. Basal cell carcinoma preferentially arises from stem cells within hair follicle and mechanosensory niches. Cell Stem Cell 16, 400–412 (2015).
Cohen, S., Levi-Montalcini, R. & Hamburger, V. A nerve growth-stimulating factor isolated from sarcomas 37 and 180. Proc. Natl Acad. Sci. USA 40, 1014–1018 (1954).
Ayala, G. E. et al. Cancer-related axonogenesis and neurogenesis in prostate cancer. Clin. Cancer Res. 14, 7593–7603 (2008).
Kobayashi, T., Kihara, K., Hyochi, N., Masuda, H. & Sato, K. Spontaneous regeneration of the seriously injured sympathetic pathway projecting to the prostate over a long period in the dog. BJU Int. 91, 868–872 (2003).
Francis, F. et al. Doublecortin is a developmentally regulated, microtubule-associated protein expressed in migrating and differentiating neurons. Neuron 23, 247–256 (1999).
Couillard-Despres, S. et al. Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 21, 1–14 (2005).
Brown, J. P. et al. Transient expression of doublecortin during adult neurogenesis. J. Comp. Neurol. 467, 1–10 (2003).
Zhang, J. & Jiao, J. Molecular biomarkers for embryonic and adult neural stem cell and neurogenesis. BioMed Res. Int. 2015, 727542 (2015).
Kaplan, M. P., Chin, S. S., Fliegner, K. H. & Liem, R. K. Alpha-internexin, a novel neuronal intermediate filament protein, precedes the low molecular weight neurofilament protein (NF-L) in the developing rat brain. J. Neurosci. 10, 2735–2748 (1990).
Schult, D. et al. Expression pattern of neuronal intermediate filament α-internexin in anterior pituitary gland and related tumors. Pituitary 18, 465–473 (2015).
Carden, M. J., Trojanowski, J. Q., Schlaepfer, W. W. & Lee, V. M. Two-stage expression of neurofilament polypeptides during rat neurogenesis with early establishment of adult phosphorylation patterns. J. Neurosci. 7, 3489–3504 (1987).
Walker, T. L., Yasuda, T., Adams, D. J. & Bartlett, P. F. The doublecortin-expressing population in the developing and adult brain contains multipotential precursors in addition to neuronal-lineage cells. J. Neurosci. 27, 3734–3742 (2007).
Lawson, D. A. et al. Basal epithelial stem cells are efficient targets for prostate cancer initiation. Proc. Natl Acad. Sci. USA 107, 2610–2615 (2010).
Ni, J. et al. Epithelial cell adhesion molecule (EpCAM) is associated with prostate cancer metastasis and chemo/radioresistance via the PI3K/Akt/mTOR signaling pathway. Int. J. Biochem. Cell Biol. 45, 2736–2748 (2013).
Pastrana, E., Cheng, L. C. & Doetsch, F. Simultaneous prospective purification of adult subventricular zone neural stem cells and their progeny. Proc. Natl Acad. Sci. USA 106, 6387–6392 (2009).
Chaker, Z., Codega, P. & Doetsch, F. A mosaic world: puzzles revealed by adult neural stem cell heterogeneity. Wiley Interdiscip. Rev. Dev. Biol. 5, 640–658 (2016).
Suh, H. et al. In vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2 + neural stem cells in the adult hippocampus. Cell Stem Cell 1, 515–528 (2007).
Sakakibara, S. et al. Mouse-Musashi-1, a neural RNA-binding protein highly enriched in the mammalian CNS stem cell. Dev. Biol. 176, 230–242 (1996).
Boutin, C. et al. NeuroD1 induces terminal neuronal differentiation in olfactory neurogenesis. Proc. Natl Acad. Sci. USA 107, 1201–1206 (2010).
Wang, S. et al. Prostate-specific deletion of the murine Pten tumor suppressor gene leads to metastatic prostate cancer. Cancer Cell 4, 209–221 (2003).
Guy, C. T., Cardiff, R. D. & Muller, W. J. Induction of mammary tumors by expression of polyomavirus middle T oncogene: a transgenic mouse model for metastatic disease. Mol. Cell. Biol. 12, 954–961 (1992).
Codega, P. et al. Prospective identification and purification of quiescent adult neural stem cells from their in vivo niche. Neuron 82, 545–559 (2014).
Liebig, C., Ayala, G., Wilks, J. A., Berger, D. H. & Albo, D. Perineural invasion in cancer: a review of the literature. Cancer 115, 3379–3391 (2009).
Sorrells, S. F. et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 555, 377–381 (2018).
Boldrini, M. et al. Human hippocampal neurogenesis persists throughout aging. Cell Stem Cell 22, 589–599.e585 (2018).
Kempermann, G. et al. Human adult neurogenesis: evidence and remaining questions. Cell Stem Cell 23, 25–30 (2018).
Ernst, A. & Frisén, J. Adult neurogenesis in humans–common and unique traits in mammals. PLoS Biol. 13, e1002045 (2015).
Ernst, A. et al. Neurogenesis in the striatum of the adult human brain. Cell 156, 1072–1083 (2014).
Ducy, P. et al. Leptin inhibits bone formation through a hypothalamic relay: a central control of bone mass. Cell 100, 197–207 (2000).
Takeda, S. et al. Leptin regulates bone formation via the sympathetic nervous system. Cell 111, 305–317 (2002).
Rhee, S. H., Pothoulakis, C. & Mayer, E. A. Principles and clinical implications of the brain–gut–enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 6, 306–314 (2009).
Magnon, C. Role of the autonomic nervous system in tumorigenesis and metastasis. Mol. Cell. Oncol. 2, e975643 (2015).
Engblom, C. et al. Osteoblasts remotely supply lung tumors with cancer-promoting SiglecFhigh neutrophils. Science 358, eaal5081 (2017).
Schagen, S. B. et al. Monitoring and optimising cognitive function in cancer patients: present knowledge and future directions. EJC Suppl. 12, 29–40 (2014).
Dietrich, J. & Kesari, S. Effect of cancer treatment on neural stem and progenitor cells. Cancer Treat. Res. 150, 81–95 (2009).
Guo, W., Patzlaff, N. E., Jobe, E. M. & Zhao, X. Isolation of multipotent neural stem or progenitor cells from both the dentate gyrus and subventricular zone of a single adult mouse. Nat. Protoc. 7, https://doi.org/10.1038/nprot.2012.123 (2012).
Hagihara, H., Toyama, K., Yamasaki, N. & Miyakawa, T. Dissection of hippocampal dentate gyrus from adult mouse. J. Vis. Exp. 33, e1543 (2009).
Acknowledgements
We thank the iRCM animal facility and the MIRcen core facility for stereotaxy; J. Baijer for assistance with cell sorting and analysis; A Bourdais for technical assistance with preliminary data; and P. Soyeux for human tissue sectioning. This work was supported by a CEA/DRF/IBFJ/iRCM starting grant, the ATIP-AVENIR group leader program (INSERM, Ligue Contre le Cancer and Plan Cancer) and ARC.
Author information
Authors and Affiliations
Contributions
P.M. and V.B. performed experiments, N.T. analysed the RNA sequencing, and performed statistical analysis, A.B. produced lentiviral vectors and performed stereotaxic surgery, V.F. and Y.A. provided the cohort of patients, P.-H.R. wrote the manuscript, and C.M. designed, performed and supervised all the experiments, prepared figures and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
A patent application, entitled ‘Methods for predicting outcome and treatment of patients suffering from prostate and breast cancer’, has been filed under the number EP18305460.0 on 13 April 2018, at the European Patent Office.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
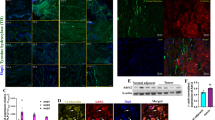
Extended Data Fig. 1 DCX+ cells express INA, a neural precursor marker, but not NF-H, a neuron-specific marker, in human prostate and Hi-MYC mouse prostate tumours, and are associated with tumour aggressiveness.
a, In human adenocarcinoma, DCX+ cells (green) and large peri-tumour nerve fibres (white arrow) express INA (red), a marker of developing neural cells and mature fibres. By contrast, mature nerve fibres express the neuron-specific cytoskeletal subunit of NF-H (light blue) but not DCX, a specific marker of neural progenitors in the central nervous system. DAPI, dark blue. Scale bar, 20 μm. b, Early time point analysis of recurrence-free survival of patients with high-risk prostate cancer. Early recurrence after radical prostatectomy is associated with high density of DCX+ cells (>20 DCX+ cells per field, n = 13). P = 0.0338, log-rank (Mantel–Cox). This is the same cohort of patients as shown in Fig. 1i, but here is focused on high-risk tumours. c, d, DCX+ cells are also present in mouse prostate tumours and are stained for PSA-NCAM (red) (c) and INA (red) (d) but do not express mature nerve markers such as NF-H (light blue), which suggests that they are neural progenitors. DCX, green; DAPI, dark blue. Scale bar, 20 μm. Three independent experiments.
Extended Data Fig. 2 Characterization and quantification of DCX–eYFP+-expressing cells in a Cre-lox mouse model of cancer.
a, Triple-transgenic DCX-creERT2;loxp-eYFP Hi-MYC mouse model of cancer mice is generated by crossing mice expressing MYC under the probasin promoter (AAR2/PBSN–MYC) with mice that express tamoxifen-inducible CreERT2 recombinase under the control of the DCX promoter/enhancer and bred with mice that have the eYFP gene inserted in the Gt(Rosa)26Sor locus with an upstream loxP-flanked STOP sequence (top). Timeline of tamoxifen injections (bottom). b. Schematics of axial, coronal and sagittal sections of the adult mouse brain showing the olfactory bulbs and olfactory tract (OT), the SVZ along the lateral ventricle, and the dentate gyrus. c. Images of DCX–eYFP+ neural precursors (yellow), in olfactory bulbs, from mice described in a, co-stained with anti-PSA-NCAM (green) and anti-INA (red) antibodies. DAPI, dark blue. Scale bar, 20 μm. Three independent experiments. d, FACS plots of eYFP+ cells isolated from Lin− cells from the brain (SVZ, olfactory bulbs, dentate gyrus and tumour tissues of DCX-creERT2;loxp-eYFP Hi-MYC mice (red) compared to control littermates (green and yellow). e, Quantification of Lin−eYFP+ cells in SVZ, olfactory bulbs and dentate gyrus (top) and prostate tumour (bottom). Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). Sample sizes are listed in Source Data. f, Lin−eYFP+ cells in Hi-MYC prostate tumours do not derive from tumour cells. Real-time quantitative PCR analyses of mRNA extracted from Lin−eYFP+ cells isolated from prostate tumour tissues, olfactory bulbs and the SVZ of DCX-creERT2;loxp-eYFP Hi-MYC mice. The Lin−eYFP+ cells do not express MYC oncogene or the cytokeratins (CK5 and CK18) present in the Lin+eYFP− fraction that comprises prostate epithelial cells. Data are mean + s.e.m.
Extended Data Fig. 3 Lin−eYFP+ cells in prostate tumours express markers of neural precursors from the central nervous system.
Representative FACS plots of SCA-1, PSA-NCAM, CD24 and EGFR expression in neural Lin−eYFP+ subpopulations from the brain at week eight after birth (SVZ, olfactory bulbs and dentate gyrus) and the prostate tumour at four months after birth. Note that two subpopulations of Lin−eYFP+ cells from prostate tumours express SCA-1 (prostate stromal cells are Lin−SCA-1+). Representative data from 48 independent experiments.
Extended Data Fig. 4 DCX–eYFP+progenitors in prostate tumours can proliferate and/or differentiate into neurons ex vivo.
a, Lin−eYFP+ cells do not display immune or endothelial phenotypes. Scatter plot representations (also known as ImmGen W-plots) of the mean-normalized expression values of the 200 most-highly expressed genes of each sample (SVZ, olfactory bulb or prostate, n = 2 per region) in each of the selected ImmGen cell populations. ImmGen populations are coloured on the basis of their main cell population families, listed in Supplementary Table 5 with correlation coefficients and P values. b, Differentiation of DCX–eYFP+ cells from prostate tumour tissues after one or eight days culture in neural medium supplemented with epidermal growth factor and basic fibroblast growth factor (EGF and bFGF, proliferation medium), or brain-derived neurotrophic factor and neurotrophin-3 (BDNF and NT-3, differentiation medium). Scale bar, 400 μm. Two independent experiments. c, Immunostaining of MAP2+ differentiated DCX–eYFP+ neural cells isolated from tumour (right) or olfactory bulbs (left). Scale bar, 20 μm. Three independent experiments. d, e, Quantification of proliferation of neural progenitors (d) and differentiated neural cells (e) with one or more neurites (red arrows (for example in b)) for eight days. Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). ***P < 0.001. Two independent experiments.
Extended Data Fig. 5 Genetic lineage tracing of Lin−eYFP+ progenitors during prostate development.
a, After tamoxifen-induced recombination of DCX-creERT2;loxp-eYFP Hi-MYC cancer mice at week 3 after birth, Lin−eYFP + neural progenitors are detected in tumour tissues during the early phases of cancer development, without infiltrating healthy tissues (n = 73), such as testis (n = 34), epididymis (n = 35) and cremaster (n = 29), that surround the tumour. Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). Twelve independent experiments. Sample sizes per group are listed in Source Data. b, FACS plots of Lin−eYFP+ cells from the prostate tumour of 6- or 16-week-old Hi-MYC mice. c, Oscillation of Lin−eYFP+ neural progenitors during prostate tumour development in the SVZ (left, n = 163) and olfactory bulbs (middle, n = 159), but not in the dentate gyrus (right, n = 59) of Hi-MYC cancer mice. Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). Forty-six independent experiments. Sample sizes per group are listed in Source Data. d, Further monitoring shows marked oscillations of Lin−eYFP+SCA-1−PSA-NCAM−CD24−EGFR− cells present in the olfactory bulbs, as observed in SVZ in Fig. 4b. Data are mean + s.e.m. Student;s t-test (one-sided, no adjustment). Forty-six independent experiments. Sample sizes are listed in Source Data. e–g, Tamoxifen is not cytotoxic to prostate or brain tissues, as shown by the fact that it does not affect the development of tumours in the Hi-MYC mouse model of cancer (e) and does not alter the cellularity (top) and viability (bottom) of cells in SVZ (f) or olfactory bulbs (g) over time. Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). Sample sizes are listed in Source Data. *P < 0.05, **P < 0.01, ***P < 0.001.
Extended Data Fig. 6 In Hi-MYC mouse model of cancer, Lin−eYFP+ progenitors egress the SVZ, migrate through the blood and infiltrate the tumour, where they do not express neural stem-cell markers.
a, b, After tamoxifen-induced recombination in the DCX-creERT2;loxp-eYFP Hi-MYC cancer mice at week 3 after birth, the number of Lin−eYFP + neural progenitors decreases in the SVZ at week 6 after birth (a) and—in turn—Lin−eYFP + cells emerge progressively in blood (b). Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). Sample sizes are listed in Source Data. Thirteen independent experiments. c, d, Representative FACS plots of Lin−eYFP+ cells isolated from the SVZ (c) or blood (d) of 6- or 16-week-old wild-type or Hi-MYC mice. ****P < 0.0001. e, Lin−eYFP+SCA-1−PSA-NCAM− cells in the SVZ—but not in prostate tumours—express neural stem-cell markers. Real-time quantitative PCR analyses of mRNA extracts obtained from purified subpopulations of the SVZ and prostate tumour tissues of DCX-creERT2;loxp-eYFP Hi-MYC mice. The Lin−eYFP+SCA-1−PSA-NCAM− cells from the SVZ express GFAP and GLAST, which are specific markers of neural stem cells. By contrast, Lin−eYFP+SCA-1−PSA-NCAM− cells in tumours do not express the stem-cell markers. Three independent experiments. Data are mean + s.e.m.
Extended Data Fig. 7 tdTomato-expressing neural precursors migrate from the SVZ towards olfactory bulbs.
a, Immunostaining of neural precursors, transduced by a tdTomato-expressing lentiviral vector administered by stereotaxic injection into the SVZ of tamoxifen-injected DCX-creERT2;loxp-EYFP Hi-MYC cancer mice, tdTomato+ cells leave the SVZ to reach the olfactory bulb area through the physiological rostral migratory stream, and differentiate into neurons in olfactory bulbs (top, white arrows, tdTomato+eYFP+ cells). Note that these tdTomato-expressing neural cells do not spread in the dentate gyrus (bottom). Scale bars, 20 μm. b, c, FACS plots showing tamoxifen-induced eYFP+ (green), lentiviral vector-transduced tdTomato+ (red, tdTomato+) and eYFP+tdTomato+ (yellow) neural precursors in the SVZ, olfactory bulbs and dentate gyrus of five-month-old Hi-MYC mice (b) and infiltration of these cells into prostate tumours (c). d, Lineage tracing of Lin−tdTomato+ cells that infiltrate prostate tumours but not healthy tissues surrounding the tumour (testis, epididymis and cremaster). Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). Sample sizes are listed in Source Data. ***P < 0.001. e, Summary of the five developmental stages of neural progenitors migrating (through the blood) from the central nervous system to the tumour during cancer development. tdTom+, tdTomato+.
Extended Data Fig. 8 Lin−tdTomato+ cells cannot reach the tumour after lentiviral injection in the dentate gyrus.
a, Frequencies of Lin−tdTomato+ cells in prostate tumours at 16 weeks of age, after tdTomato+ lentiviral vector injection by stereotaxy into the dentate gyrus (n = 6) at week 5 after birth, by comparison to an injection into the SVZ (n = 10 mice). Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). ***P < 0.001. b, Absolute numbers of Lin−tdTomato+ cells in the SVZ (n = 9) or dentate gyrus (n = 8) 16 weeks after the injection of the tdTomato+ lentiviral vector in the respective brain area. Data are mean + s.e.m. c. Respective mean fluorescence intensities (MFI) for tdTomato signal in SVZ (MFI/SVZ, red) and dentate gyrus (MFI/DG, grey) shown in b. d, Dentate gyrus vascularization (CD31+, pink) in one-year-old Hi-MYC mice does not exhibit any leakiness of albumin (orange) in the dentate gyrus. DCX–eYFP+ cells (green). DAPI, dark blue. Scale bar, 30 μm. e, The blood–brain barrier is breached in the SVZ of Hi-MYC cancer mice. Combined injection of fluorescein–albumin (65 kDa, green) and TRITC–dextran (4.4 kDa, red) reveals extravasation of both dyes in SVZ tissues (top) throughout sinusoids. In the dentate gyrus, the dyes do not spread outside of the capillaries (bottom). DAPI, dark blue. Scale bars, 20 μm. tdTom+, tdTomato+.
Extended Data Fig. 9 Lin− tdTomato+ cells can infiltrate Pten prostate tumours, PyMT breast tumours and metastases.
a, b, After injection of a tdTomato-expressing lentiviral vector by stereotaxy into the SVZ at week 5 after birth, Lin−tdTomato+ cells are found at week 16 after birth in Pten prostate tumour tissues (a) or PyMT breast tumour tissues (b), by contrast to wild-type prostate or mammary pad tissues. c, Frequencies of Lin−tdTomato+ cells in Pten mice (n = 5) or wild-type littermates (n = 8). Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). **P < 0.01. Lin−tdTomato+ cells are also found in metastases. d, e, Luciferase-expressing human PC3 cancer cells orthotopically xenografted in the mouse prostate disseminate to colon, liver, lung and lymph nodes between 12 to 14 weeks after graft (d) and metastatic tissues were carefully dissected before be stained with a combination of haematoxylin and eosin (e). Scale bar, 300 μm. f, Neural progenitors transduced by a tdTomato-expressing lentiviral vector administered by stereotaxy into the SVZ, travel to PC3 xenografts and developing metastases in colon, liver, lung and lymph nodes at weeks 12–14 after graft. g, Frequencies of Lin−tdTomato+ cells in tumour tissues, listed above, by comparison to healthy tissues. Data are mean + s.e.m. Student’s t-test (one-sided, no adjustment). *P < 0.05, **P < 0.01. Sample sizes per group are listed in Source Data. tdTom+, tdTomato+.
Extended Data Fig. 10 Genetic depletion of DCX+ cells.
a, b, Timeline for MYC-induced tumour development, injections of tamoxifen and diphtheria toxin treatment 48 h after the last injection of tamoxifen (a) and experimental protocols (b). c, Representative H & E-stained sections of prostates of 20-week-old Hi-MYC mice.
Supplementary information
Supplementary Information
This file contains Supplementary Tables 1 – 6.
Source data
Rights and permissions
About this article

Cite this article
Mauffrey, P., Tchitchek, N., Barroca, V. et al. Progenitors from the central nervous system drive neurogenesis in cancer. Nature 569, 672–678 (2019). https://doi.org/10.1038/s41586-019-1219-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-1219-y
This article is cited by
-
Stage II oesophageal carcinoma: peril in disguise associated with cellular reprogramming and oncogenesis regulated by pseudogenes
BMC Genomics (2024)
-
The nerve cells in gastrointestinal cancers: from molecular mechanisms to clinical intervention
Oncogene (2024)
-
How cancer hijacks the nervous system to grow and spread
Nature (2024)
-
Acid-sensing ion channels and downstream signalling in cancer cells: is there a mechanistic link?
Pflügers Archiv - European Journal of Physiology (2024)
-
The nerve growth factor-delivered signals in prostate cancer and its associated microenvironment: when the dialogue replaces the monologue
Cell & Bioscience (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
