
Abstract
A hominin third metatarsal discovered in 2007 in Callao Cave (Northern Luzon, the Philippines) and dated to 67 thousand years ago provided the earliest direct evidence of a human presence in the Philippines. Analysis of this foot bone suggested that it belonged to the genus Homo, but to which species was unclear. Here we report the discovery of twelve additional hominin elements that represent at least three individuals that were found in the same stratigraphic layer of Callao Cave as the previously discovered metatarsal. These specimens display a combination of primitive and derived morphological features that is different from the combination of features found in other species in the genus Homo (including Homo floresiensis and Homo sapiens) and warrants their attribution to a new species, which we name Homo luzonensis. The presence of another and previously unknown hominin species east of the Wallace Line during the Late Pleistocene epoch underscores the importance of island Southeast Asia in the evolution of the genus Homo.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

Data availability
The data that support the findings of this study are available from the corresponding authors upon reasonable request. Homo luzonensis has been deposited in the ZooBank database (http://zoobank.org/) with Life Science Identifier urn:lsid:zoobank.org:act:4F743862-662F-4E6B-9812-8A05533C1347. The description of the species has been deposited with Life Science identifier urn:lsid:zoobank.org:pub:0E4607F1-1374-4842-B32B-7CE2250807DF.
References
Mijares, A. S. et al. New evidence for a 67,000-year-old human presence at Callao Cave, Luzon, Philippines. J. Hum. Evol. 59, 123–132 (2010).
Détroit, F., Corny, J., Dizon, E. Z. & Mijares, A. S. “Small size” in the Philippine human fossil record: is it meaningful for a better understanding of the evolutionary history of the Negritos? Hum. Biol. 85, 45–66 (2013).
Grün, R., Eggins, S., Kinsley, L., Moseley, H. & Sambridge, M. Laser ablation U-series analysis of fossil bones and teeth. Palaeogeogr. Palaeoclimatol. Palaeoecol. 416, 150–167 (2014).
Kaifu, Y. et al. Taxonomic affinities and evolutionary history of the Early Pleistocene hominids of Java: dentognathic evidence. Am. J. Phys. Anthropol. 128, 709–726 (2005).
Zanolli, C. Additional evidence for morpho-dimensional tooth crown variation in a New Indonesian H. erectus sample from the Sangiran Dome (central Java). PLoS ONE 8, e67233 (2013).
Kaifu, Y. et al. Descriptions of the dental remains of Homo floresiensis. Anthropol. Sci. 123, 129–145 (2015).
Kaifu, Y. et al. Unique dental morphology of Homo floresiensis and its evolutionary implications. PLoS ONE 10, e0141614 (2015).
Grine, F. E. & Franzen, J. L. Fossil hominid teeth from the Sangiran dome (Java, Indonesia). Courier Forschungsinstitut Senckenberg (CFS) 171, 75–103 (1994).
Xing, S., Martinón-Torres, M. & Bermúdez de Castro, J. M. The fossil teeth of the Peking Man. Sci. Rep. 8, 2066 (2018).
Sawyer, S. et al. Nuclear and mitochondrial DNA sequences from two Denisovan individuals. Proc. Natl Acad. Sci. USA 112, 15696–15700 (2015).
Gómez-Robles, A., Martinón-Torres, M., Bermúdez de Castro, J. M., Prado-Simón, L. & Arsuaga, J. L. A geometric morphometric analysis of hominin upper premolars. Shape variation and morphological integration. J. Hum. Evol. 61, 688–702 (2011).
Xing, S. et al. Middle Pleistocene hominin teeth from Longtan Cave, Hexian, China. PLoS ONE 9, e114265 (2014).
Wood, B. A. & Engleman, C. A. Analysis of the dental morphology of Plio-Pleistocene hominids. V. Maxillary postcanine tooth morphology. J. Anat. 161, 1–35 (1988).
Monsarrat, P. et al. Interrelationships in the variability of root canal anatomy among the permanent teeth: a full-mouth approach by cone-beam CT. PLoS ONE 11, e0165329 (2016).
Scott, G. R., Turner, C. G. II, Townsend, G. C. & Martinón-Torres, M. The Anthropology of Modern Human Teeth: Dental Morphology and Its Variation in Recent and Fossil Homo sapiens (Cambridge Univ. Press, Cambridge, 2018).
Irish, J. D., Bailey, S. E., Guatelli-Steinberg, D., Delezene, L. K. & Berger, L. R. Ancient teeth, phenetic affinities, and African hominins: another look at where Homo naledi fits in. J. Hum. Evol. 122, 108–123 (2018).
van den Bergh, G. D. et al. Homo floresiensis-like fossils from the early Middle Pleistocene of Flores. Nature 534, 245–248 (2016).
Bush, M. E., Lovejoy, C. O., Johanson, D. C. & Coppens, Y. Hominid carpal, metacarpal, and phalangeal bones recovered from the Hadar formation: 1974–1977 collections. Am. J. Phys. Anthropol. 57, 651–677 (1982).
Susman, R. L., Stern, J. T. Jr & Jungers, W. L. Arboreality and bipedality in the Hadar hominids. Folia Primatol. 43, 113–156 (1984).
Ward, C. V., Kimbel, W. H., Harmon, E. H. & Johanson, D. C. New postcranial fossils of Australopithecus afarensis from Hadar, Ethiopia (1990–2007). J. Hum. Evol. 63, 1–51 (2012).
Napier, J. Fossil hand bones from Olduvai Gorge. Nature 196, 409–411 (1962).
Moyà-Solà, S., Köhler, M., Alba, D. M. & Almécija, S. Taxonomic attribution of the Olduvai hominid 7 manual remains and the functional interpretation of hand morphology in robust australopithecines. Folia Primatol. 79, 215–250 (2008).
Larson, S. G. et al. Descriptions of the upper limb skeleton of Homo floresiensis. J. Hum. Evol. 57, 555–570 (2009).
Trinkaus, E. The Shanidar Neandertals (Academic, London, 1983).
Walker, A. & Leakey, R. E. The Nariokotome Homo erectus skeleton (Harvard Univ. Press, Cambridge, 1993).
Lorenzo, C., Arsuaga, J. L. & Carretero, J. M. Hand and foot remains from the Gran Dolina early Pleistocene site (Sierra de Atapuerca, Spain). J. Hum. Evol. 37, 501–522 (1999).
Kivell, T. L. et al. The hand of Homo naledi. Nat. Commun. 6, 8431 (2015).
Mittra, E. S., Smith, H. F., Lemelin, P. & Jungers, W. L. Comparative morphometrics of the primate apical tuft. Am. J. Phys. Anthropol. 134, 449–459 (2007).
Latimer, B. M., Lovejoy, C. O., Johanson, D. C. & Coppens, Y. Hominid tarsal, metatarsal, and phalangeal bones recovered from the Hadar formation: 1974–1977 collections. Am. J. Phys. Anthropol. 57, 701–719 (1982).
Griffin, N. L. & Richmond, B. G. Joint orientation and function in great ape and human proximal pedal phalanges. Am. J. Phys. Anthropol. 141, 116–123 (2010).
Trinkaus, E. & Patel, B. A. An Early Pleistocene human pedal phalanx from Swartkrans, SKX 16699, and the antiquity of the human lateral forefoot. C. R. Palevol 15, 978–987 (2016).
Jungers, W. L. et al. Descriptions of the lower limb skeleton of Homo floresiensis. J. Hum. Evol. 57, 538–554 (2009).
Jungers, W. L. et al. The foot of Homo floresiensis. Nature 459, 81–84 (2009).
Harcourt-Smith, W. E. H. et al. The foot of Homo naledi. Nat. Commun. 6, 8432 (2015).
DeSilva, J. M. et al. The lower limb and mechanics of walking in Australopithecus sediba. Science 340, 1232999 (2013).
Stern, J. T. J. Jr & Susman, R. L. The locomotor anatomy of Australopithecus afarensis. Am. J. Phys. Anthropol. 60, 279–317 (1983).
Latimer, B., Ohman, J. C. & Lovejoy, C. O. Talocrural joint in African hominoids: implications for Australopithecus afarensis. Am. J. Phys. Anthropol. 74, 155–175 (1987).
Westaway, K. E. et al. An early modern human presence in Sumatra 73,000–63,000 years ago. Nature 548, 322–325 (2017).
Sutikna, T. et al. Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia. Nature 532, 366–369 (2016).
Li, Z.-Y. et al. Late Pleistocene archaic human crania from Xuchang, China. Science 355, 969–972 (2017).
Bellwood, P. The search for ancient DNA heads east. Science 361, 31–32 (2018).
Ingicco, T. et al. Earliest known hominin activity in the Philippines by 709 thousand years ago. Nature 557, 233–237 (2018).
Brown, P. et al. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431, 1055–1061 (2004).
Jungers, W. L. in A Companion to Paleoanthropology (ed. Begun, D. R.) 582–598 (Blackwell, Hoboken, 2013).
Argue, D., Groves, C. P., Lee, M. S. Y. & Jungers, W. L. The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters. J. Hum. Evol. 107, 107–133 (2017).
Voris, H. K. Maps of Pleistocene sea levels in Southeast Asia: shorelines, river systems and time durations. J. Biogeogr. 27, 1153–1167 (2000).
Heaney, L. R., Balete, D. S. & Rickart, E. A. The Mammals of Luzon Island (Johns Hopkins Univ. Press, Baltimore, 2016).
Heaney, L. R. A synopsis of the mammalian fauna of the Philippine Islands. Fieldiana Zool. 88, 1–61 (1998).
van der Geer, A. A. E., Lomolino, M. V. & Lyras, G. A. ‘Island Life’ before man: biogeography of palaeo-insular mammals. J. Biogeogr. 44, 995–1006 (2017).
van den Hoek Ostende, L. W. Cladistics and insular evolution, an unfortunate marriage? Another tangle in the Deinogalerix analysis of Borrani et al. (2017). Cladistics 34, 708–713 (2018).
Spoor, C. F., Zonneveld, F. W. & Macho, G. A. Linear measurements of cortical bone and dental enamel by computed tomography: applications and problems. Am. J. Phys. Anthropol. 91, 469–484 (1993).
Duval, M. & Martín-Francés, L. Quantifying the impact of µCT-scanning of human fossil teeth on ESR age results. Am. J. Phys. Anthropol. 163, 205–212 (2017).
Grün, R., Athreya, S., Raj, R. & Patnaik, R. ESR response in tooth enamel to high-resolution CT scanning. Archaeol. Anthropol. Sci. 4, 25–28 (2012).
Immel, A. et al. Effect of X-ray irradiation on ancient DNA in sub-fossil bones — guidelines for safe X-ray imaging. Sci. Rep. 6, 32969 (2016).
Wood, B. Koobi Fora Research Project. Volume 4, Hominid Cranial Remains (Clarendon, Oxford, 1991).
Duncan, A. S., Kappelman, J. & Shapiro, L. J. Metatarsophalangeal joint function and positional behavior in Australopithecus afarensis. Am. J. Phys. Anthropol. 93, 67–81 (1994).
R Core Team. R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, 2017).
Claude, J. Morphometrics with R (Springer, 2008).
Dray, S. & Dufour, A.-B. The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22, 1–20 (2007).
Adams, D. C., Collyer, M. L. & Kaliontzopoulou, A. Geomorph: software for geometric morphometric analyses. R package version 3.0.6 https://cran.r-project.org/package=geomorph (2018).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer-Verlag, 2009).
Kassambara, A. ggpubr: ‘ggplot2’ based publication ready plots. R package version 0.1.8 https://cran.r-project.org/web/packages/ggpubr/ (2017).
Bonhomme, V., Picq, S., Gaucherel, C. & Claude, J. Momocs: outline analysis using R. J. Stat. Softw. 56, 1–24 (2014).
Schlager, S. in Statistical Shape and Deformation Analysis (eds Zheng, G. et al.) 217–256 (Academic, 2017).
Adler, D. et al. rgl: 3D visualization using OpenGL. R package version 0.99.16 https://cran.r-project.org/web/packages/rgl/ (2018).
Dryden, I. L. shapes. R package version 1.2.4 https://cran.r-project.org/web/packages/shapes/ (2017).
Oksanen, J. et al. vegan: community ecology package. R package version 2.5-4 https://cran.r-project.org/web/packages/vegan/ (2018).
Darroch, J. N. & Mosimann, J. E. Canonical and principal components of shape. Biometrika 72, 241–252 (1985).
Jungers, W. L., Falsetti, A. B. & Wall, C. E. Shape, relative size, and size-adjustments in morphometrics. Am. J. Phys. Anthropol. 38, 137–161 (1995).
Musgrave, J. H. in Human Evolution Vol. 11 (ed. Day, M. H.) 59–85 (Taylor & Francis, London, 1973).
Corny, J. & Détroit, F. Anatomic identification of isolated modern human molars: testing procrustes aligned outlines as a standardization procedure for elliptic Fourier analysis. Am. J. Phys. Anthropol. 153, 314–322 (2014).
Corny, J. et al. Dental phenotypic shape variation supports a multiple dispersal model for anatomically modern humans in Southeast Asia. J. Hum. Evol. 112, 41–56 (2017).
Zanolli, C. et al. The Middle Pleistocene (MIS 12) human dental remains from Fontana Ranuccio (Latium) and Visogliano (Friuli-Venezia Giulia), Italy. A comparative high resolution endostructural assessment. PLoS ONE 13, e0189773 (2018).
Zanolli, C. et al. Inner tooth morphology of Homo erectus from Zhoukoudian. New evidence from an old collection housed at Uppsala University, Sweden. J. Hum. Evol. 116, 1–13 (2018).
Susman, R. L. Comparative and functional morphology of hominoid fingers. Am. J. Phys. Anthropol. 50, 215–236 (1979).
Case, D. T. & Heilman, J. New siding techniques for the manual phalanges: a blind test. Int. J. Osteoarchaeol. 16, 338–346 (2006).
Christensen, A. M. Techniques for siding manual phalanges. Forensic Sci. Int. 193, 84–87 (2009).
Mitteroecker, P. & Bookstein, F. Linear discrimination, ordination, and the visualization of selection gradients in modern morphometrics. Evol. Biol. 38, 100–114 (2011).
Acknowledgements
F.D., J.C., G.D. and C.Z. thank L. Puymerail. We thank T. Sutikna and W. Saptomo (Pusat Penelitian Arkeologi Nasional); X.-J. Wu and W. Liu (IVPP); J. de Vos (Naturalis); N. Giang Hai, N. Kim Thuy, N. Lan Cuong and N. Thi Mai Huong (Institute of Archaeology); N. Viet (Center for Southeast Asian Prehistory); A. Kijngam and R. Thosarat (Thai Fine Arts Department); the National Research Council of Thailand; G. Garcia-Pack, I. Tattersall and E. Delson (American Museum of Natural History); the Field Museum of Natural History; the Ethiopian Authority for Research and Conservation of the Cultural Heritage; the National Museums of Kenya and Government of Kenya; the Ditsong National Museum of Natural History, the Evolutionary Studies Institute, University of the Witwatersrand; the Centre de Valorisation des Collections scientifiques de l’Université de Poitiers; S. Bahuchet, A. Fort, M. Friess, A. Froment, D. Grimaud-Hervé, L. Huet, V. Laborde and P. Mennecier (MNHN); the Nespos Society (https://www.nespos.org); MorphoSource, Duke University (https://www.morphosource.org/); the Senckenberg Research Institute Frankfurt; the ESRF Paleontological Database (paleo.esrf.eu); P. Fernandez, C. Higham, W. Jungers, Y. Kaifu, R. Kono, H. Matsumura, M. Oxenham, N. Tayles and M. Tocheri for granting access to or sharing information and data on recent and fossil hominins; M. Stoneking and Q. Fu (Max Planck Institute for Evolutionary Anthropology); AST-RX and Imagerie 2D/3D MH-SU platforms (MNHN); G. Champion, A. Giral, A. Gossez, A. Van Heteren, J. Surault and J. Tanedo for discussions or technical assistance; the National Museum of the Philippines and its Directorial staff; colleagues and students from the Archaeological Studies Program (University of the Philippines); the 2011 and 2015 IMQP Erasmus Mundus students; and the Cagayan Provincial Government and the Protected Area Management Board-Peñablanca for authorizing fieldwork at Callao Cave. Funding was provided by the Wenner-Gren Foundation, the Leakey Foundation Research Grant and the University of the Philippines Enhanced Creative Work and Research Grant (ECWRG 2015-1-016). R.G.’s work was partly funded by the Australian Research Council Grant DP110101415. The research of P.J.P. was supported by Australian Research Council grants FT100100527 and DP140100384. The research of G.D. was supported by the Agence Nationale pour la Recherche (ANR ARCHOR-12-CULT-0006), the Omo Group Research Expedition (J.-R. Boisserie), the MEAE, the Université de Poitiers and the Conseil départemental de la Vienne (ACI 2013-2014). The research of J.C. was supported by SYNTHESYS (NL-TAF-1130) and ‘IUSS Ferrara 1391’ grants. Aspects of the research of F.D. were supported by the Labex BCDiv ‘Biological and Cultural Diversities’, Erasmus Mundus IMQP and IDQP, MQPI (MEAE) and PaléoFED.
Reviewer information
Nature thanks Aida Gómez-Robles, William L. Jungers, Yousuke Kaifu and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
F.D., J.C., G.D. and C.Z. analysed the hominin fossils, A.S.M. is the project leader, E.D. and E.R. are senior members of the excavation team, R.G. conducted the dating, F.D. and P.J.P. wrote the manuscript, F.D., J.C., G.D., C.Z. and P.J.P. edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Inventory of the fossil elements attributed to H. luzonensis and detailed views of the dental remains.
a, The hominin fossils recovered from Callao Cave. R, right; L, left; P, premolar; M, molar. b, Three-dimensional rendering of the postcanine maxillary teeth CCH6-b to CCH6-e (M2–P3): occlusal (top row) and buccal (bottom row) aspects. Enamel is shown in dark blue, dentine and cement in light brown and pulp cavity in dark grey. In all views, mesial is to the right, distal to the left. c, CCH6-a, right M3: occlusal, buccal, lingual, mesial and distal aspects (from top to bottom and left to right). Occlusal view: mesial is to the right, distal to the left. d, CCH6-a to CCH6-e, right M3–P3: photograph of occlusal aspect. Mesial is to the right, distal to the left. The numbers indicate the locations of the detailed views of the inter-proximal contact facets (IPCFs): P3 (CCH6-e), mesial IPCF 1: note the small size of this IPCF, indicating that the canine was probably not large; 2–5: note the perfect match between corresponding pairs of mesial (top row) and distal (bottom row, mirrored images) IPCFs, from the P3 (CCH6-e) to the M3 (CCH6-a). e, CCH6-c, right M1: distal aspect, showing the partially fused lingual and disto-buccal roots. Lingual is to the left, buccal to the right. f, CCH8, left P3 or P4, photograph of the original fossil (occlusal view) and three-dimensional rendering: occlusal (top row), buccal (middle row) and mesio-buccal (bottom row) aspects. Enamel is shown in dark blue, dentine and cement in light brown and pulp cavity in dark grey. g, CCH9, right M3: occlusal, buccal, lingual, mesial and distal aspects (from top to bottom and left to right). Occlusal view: mesial is to the right, distal to the left; captures of the three-dimensional surface model. Scale bars, 10 mm (IPCF views in d (1–5) are not to scale); the mirrored image is indicated by an asterisk.
Extended Data Fig. 2 Dental metrics.
a, b, d–f, Bivariate scatter plots for mesio-distal (MD) versus bucco-lingual (BL) diameters of P3 (a), P4 (b), M1 (d), M2 (e) and M3 (f). c, Key for a, b, d–g, j, k. Sample sizes for a, b, d, e, f, respectively: Australopithecus, n = 23, 23, 21, 26, 26; Paranthropus, n = 22, 20, 26, 23, 20; African and European early Homo, n = 23, 19, 37, 21, 17; Asian early Homo, n = 16, 17, 17, 13, 11; H. neanderthalensis, n = 23, 26, 27, 30, 20; H. sapiens, n = 57, 61, 86, 70, 33; H. floresiensis, n = 1, 1, 1, 1, 0; H. luzonensis, n = 2∗, 2∗, 1, 1, 2 (∗CCH8 is a P3 or a P4). A detailed list of specimens can be found in Supplementary Table 4. g–i, bgPCA of the log-shape ratios of bucco-lingual and mesio-distal diameters of four postcanine maxillary teeth (P3, P4, M1 and M2), CCH6 was treated as a supplementary individual and was plotted a posteriori. g, Scatter plot of specimens for bgPC1 versus bgPC2, with convex hulls for all groups, except H. floresiensis and H. luzonensis. Sample sizes: Australopithecus, n = 6; Paranthropus, n = 5; African and European early Homo, n = 13; Asian early Homo, n = 5; H. neanderthalensis, n = 12; H. sapiens, n = 47; H. floresiensis, n = 1; H. luzonensis, n = 1. A detailed list of specimens can be found in Supplementary Table 4. h, Variable scores for bgPC1 versus bgPC2 (correlation circle; log-shape ratios of variables). i, Bar plot of eigenvalues (%) of bgPC1–bgPC5. j, k, Bivariate scatter plot of the summed square root of computed occlusal surface areas of premolars versus molars (j) and bgPCA of the log-shape ratios of bucco-lingual and mesio-distal diameters of four postcanine maxillary teeth (k) similar to the analyses presented in Fig. 3b and in g, respectively, but with ‘Negritos’ treated as a separate group (same sample sizes as in g, except for: H. sapiens, n = 38 and H. sapiens ‘Negritos’, n = 9).
Extended Data Fig. 3 Elliptic Fourier analysis of M1 crown contour.
CCH6-c compared to the holotype of H. floresiensis (LB1) and large samples of archaeological and recent H. sapiens individuals. a, PCA of shape data for all specimens, scatter plot of individual scores for PC1 versus PC2 (see Methods; elliptic Fourier descriptors applied to Procrustes-aligned outlines, ten harmonics included). LGM, Last Glacial Maximum. Sample sizes: H. luzonensis, n = 1; H. floresiensis, n = 2; pre-LGM, n = 2; pre-Neolithic post-LGM, n = 12; Neolithic/post-Neolithic, n = 232; recent ‘Negritos’, n = 19. A detailed list of specimens can be found in Supplementary Table 5. b, Bar plot of eigenvalues (%) of PC1–PC6. c, Extreme shape variations along PC1 and PC2. The scores of H. luzonensis M1 along PC1 and PC2 reflects a crown outline shape that is mesio-distally compressed, but not as much as that of H. floresiensis (two versions of the LB1 right M1). d, Right M1 of the holotype of H. floresiensis LB1 showing the two different versions of the crown outline (see Methods): the original contour (V1; in blue) published in a previous study7, and the contour (V2; in red) drawn by J.C. differ in the compensation of the mesial IPCF. These two versions differ minimally in the results of the elliptic Fourier analysis (see d and h). e, PCA of Fourier descriptors for the means of 16 groups of H. sapiens (sample sizes in brackets, see details in Supplementary Table 5), H. luzonensis CCH6-c and H. floresiensis LB1 (V1 and V2 treated as 2 groups): scatter plot of mean scores for PC1 versus PC2 with a superimposed minimum spanning tree indicating distances between groups. f, Extreme shape variations along PC1 and PC2: H. floresiensis differs from H. luzonensis in having a M1 crown contour shape that is more compressed mesio-distally, with a more developed protocone.
Extended Data Fig. 4 Procrustes analyses of the premolar and molar EDJ.
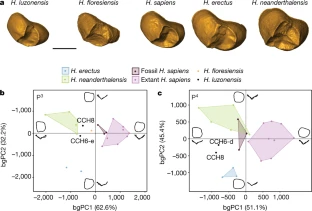
a, Landmarks placed on the main dentine horns (in red) and semilandmarks (in white) positioned along the marginal ridges of the premolar EDJ. b, Landmarks placed on the main dentine horns (in red) and the semilandmarks (in white) positioned along the marginal ridges and on the oblique crest of the molar EDJ. c, Key for d–i. d–g, PCAs of the three-dimensional landmarks Procrustes-registered shape coordinates of the P3s (d), P4s (e), M1s (f) and M2s (g). h, i, bgPCAs of the three-dimensional landmarks Procrustes-registered shape coordinates of the M1s (h) and M2s (i). Sample sizes for d, e, f and h, and g and i, respectively: H. erectus, n = 2, 3, 5, and 3; H. neanderthalensis, n = 5, 6, 5, and 6; fossil H. sapiens, n = 3, 3, 4, and 3; extant H. sapiens, n = 8, 9, 7, and 9; H. floresiensis, n = 1, 0, 0, and 0; H. luzonensis, n = 2, 2, 1, and 1. A detailed list of specimens can be found in Supplementary Table 6.
Extended Data Fig. 5 CCH2, intermediate manual phalanx of H. luzonensis.
a, Photograph of the original specimen CCH2 in palmar view. b, Three-dimensional rendering of CCH2. From left to right: palmar, lateral, dorsal, medial, distal (top), proximal (bottom), disto-lateral and proximo-lateral aspects. c, Comparison of CCH2 with Pliocene (A.L.333-88), Lower Pleistocene (OH7 (FLK NN-F) and KNM-WT 15000BO), Upper Pleistocene (LB1/48) and recent (PAPO-74-11) intermediate manual phalanges in palmar (top) and side (bottom) views. All specimens are from rays 2–5 of unknown side, except for OH7 (third ray, probably from the right hand of a juvenile individual). Note the unique proximally accentuated beak located on the dorsum of the proximal surface of CCH2. d–f, Relative robusticity of the intermediate manual phalanx CCH2. Box-and-whisker plots depicting the ratio of the interarticular length (IA) and the maximum medio-lateral width of the base (MLbase) (d), the head (MLend) (e) and the midshaft (MLmid) (f). Box, 25–75th percentiles; centre line, median; whiskers, non-outlier range; dots, outliers; dotted line, value for CCH2. AE-eH, African and European early Homo; Aus, Australopithecus; Hf, H. floresiensis; Hl, H. luzonensis; Hna, H. naledi; Hn, H. neanderthalensis; Hs, H. sapiens, Par/eH, Paranthropus/early Homo. n indicates sample size. A detailed list of specimens can be found in Supplementary Table 8. For conservative reasons, taxonomic assignation of hand remains of OH7 and several SKX specimens are considered to be uncertain (see main text and Methods). Scale bars, 10 mm.
Extended Data Fig. 6 Procrustes analyses of the intermediate manual phalanx of H. luzonensis.
CCH2 compared to specimens attributed to Australopithecus (A. afarensis, n = 1; A. africanus, n = 1; A. sediba, n = 2), Paranthropus/early Homo (Swartkrans, Member 1, n = 3; Member 3, n = 2), H. naledi (Hand 1, n = 2), H. floresiensis (LB1/48 and LB6/9) and recent H. sapiens separated into 3 samples corresponding to ray number (n = 15, 21 and 19 for rays 2, 3 and 4, respectively). A detailed list of specimens can be found in Supplementary Table 9. a–c, bgPCA of Procrustes-registered landmarks and semilandmarks (H. sapiens sample includes ray 3 only): scatter plot of individual scores for bgPC1 versus bgPC2 (a); scatter plot of individual scores for bgPC2 versus bgPC3 (b); shape variation associated with bgPC1, bgPC2 and bgPC3 (c). d, bgPCA of Procrustes-registered landmarks and semilandmarks (H. sapiens sample includes ray 4 only): scatter plot of individual scores for bgPC1 versus bgPC2. e, bgPCA of Procrustes-registered landmarks and semilandmarks (H. sapiens sample includes ray 2 only): scatter plot of individual scores for bgPC1 versus bgPC2. f, Box-and-whisker plot depicting centroid size (Procrustes registration of all specimens together). Box, 25–75th percentiles; centre line, median; whiskers, non-outlier range; dots, outliers; dotted line, value for CCH2. g, Protocol for three-dimensional Procrustes analysis: landmarks placed on the main anatomical features (n = 23, in red) and equally spaced semilandmarks (n = 250, in green) placed on the whole surface of the intermediate manual phalanx (template shown on the intermediate manual phalanx, ray 3, of the recent H. sapiens 17980).
Extended Data Fig. 7 CCH5, distal manual phalanx of H. luzonensis.
a, Photograph of the original specimen CCH5 in palmar view. b, Three-dimensional rendering of CCH5. From left to right: palmar, lateral/medial, dorsal, medial/lateral, distal (top), proximal (bottom), disto-lateral/medial and proximo-lateral/medial aspects. c, Comparison of CCH5 with Pliocene (A.L.333-11 and A.L.333-50), Lower Pleistocene (OH7 (FLK-NN-B) and SKX 27504), Upper Pleistocene (LB6/12) and recent distal manual phalanges (PAPO-74-53 and PAPO-74-11) in palmar (top) and side (bottom) views. All specimens are from rays 2–5 of unknown side, except for OH7 (second to fourth ray, probably from the right hand of a juvenile individual). d, e, Box-and-whisker plots depicting the expansion index ((apical tuft maximum medio-lateral width/maximum medio-lateral width of the base) × 100)) (d) and the robusticity index ((apical tuft maximum medio-lateral width/biomechanical length) × 100) (e) of the distal manual phalanx CCH5. Box, 25–75th percentiles, centre line, median; whiskers, non-outlier range; dots, outliers; dotted line, value for CCH5. A detailed list of specimens can be found in Supplementary Table 10. For conservative reasons, taxonomic assignation of hand remains OH7 and SKX 27504 are considered to be uncertain (see main text and Methods). Scale bars, 10 mm.
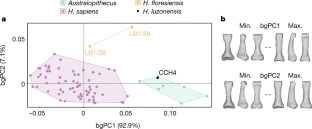
Extended Data Fig. 8 CCH4, proximal pedal phalanx of H. luzonensis.
a, Photograph of the original specimen CCH4 in lateral view. b, Three-dimensional rendering of CCH4. From left to right: plantar, lateral, dorsal, medial, distal (top), proximal (bottom), disto-lateral and proximo-lateral aspects. c, Comparison of CCH4 with Pliocene (A.L.333-115h, left third proximal phalanx, mirrored), Pleistocene (LB1/36, unknown side and rays 2–5) and recent (‘Negrito’ 9764, right third proximal pedal phalanx), proximal pedal phalanges in plantar (top) and side (bottom) views. d, Transverse (1, 2, 3) and mid-sagittal (4) micro-CT slices of CCH4 (plantar aspect of three-dimensional rendering, during the segmentation process; 1, 2, 3, dorsal is up, plantar is down, lateral is left, medial is right; 4, distal is up, proximal is down, dorsal is left, plantar is right). e, Protocol for three-dimensional Procrustes analysis: landmarks placed on the main anatomical features (n = 20, in red) and equally spaced semilandmarks (n = 250, in green) placed on the whole surface of the proximal pedal phalanx (template shown on the proximal pedal phalanx, ray 2 of the right foot of the recent H. sapiens 35071). f, g, Box-and-whisker plots depicting comparisons of the articular angle a (f) and the dorsal canting angle α (g) of the proximal pedal phalanx CCH4. Box, 25–75th percentiles; centre line, median; whiskers, non-outlier range; dots, outliers; dotted line, value for CCH4. A detailed list of specimens can be found in Supplementary Table 11. Scale bars, 10 mm.
Extended Data Fig. 9 CCH3, intermediate pedal phalanx of H. luzonensis.
a, Photograph of the original specimen CCH3 in plantar view. b, Three-dimensional rendering of CCH3. From left to right: plantar, medial, dorsal, lateral, distal (top), proximal (bottom), disto-medial and proximo-medial aspects. c, Comparison of CCH3 with Pliocene (A.L.333-21a, unknown side and rays 2–5, and A.L.333-115k, fourth intermediate phalanx), Upper Pleistocene (LB1/56, LB1-15 and LB1/39: unknown side and rays 2–5) and recent (PAPO-74-150) intermediate pedal phalanges in plantar (top) and side (bottom) views. Note the variation in shape and size both between taxa (for example, H. sapiens and H. floresiensis) and in the same individual (for example, LB1 and PAPO-74-150). A detailed list of specimens can be found in Supplementary Table 12. Scale bars, 10 mm.
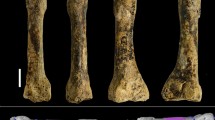
Extended Data Fig. 10 CCH7, femoral shaft of a juvenile individual of H. luzonensis.
a, Photograph of the original specimen CCH7 (posterior aspect). b, Three-dimensional rendering of CCH7. From left to right: anterior, medial, posterior and lateral aspects. Scale bar, 20 mm. c, Transverse micro-CT slices of CCH7 at proximal diaphysis (top), midshaft (middle) and distal diaphysis (bottom), and posterior aspect of the three-dimensional rendering of the femoral shaft, during the segmentation process (orientation of slices: anterior is up, posterior is down, lateral is left, medial is right).
Supplementary information
Supplementary Information
This file contains detailed morphological descriptions of the new Callao Cave Hominin (CCH) fossils attributed to Homo luzonensis, sp. nov., Supplementary Tables 1-12 and additional references.
Rights and permissions
About this article

Cite this article
Détroit, F., Mijares, A.S., Corny, J. et al. A new species of Homo from the Late Pleistocene of the Philippines. Nature 568, 181–186 (2019). https://doi.org/10.1038/s41586-019-1067-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-1067-9
This article is cited by
-
More than a decade of genetic research on the Denisovans
Nature Reviews Genetics (2024)
-
Early presence of Homo sapiens in Southeast Asia by 86–68 kyr at Tam Pà Ling, Northern Laos
Nature Communications (2023)
-
A Middle Pleistocene Denisovan molar from the Annamite Chain of northern Laos
Nature Communications (2022)
-
Constraining the chronology and ecology of Late Acheulean and Middle Palaeolithic occupations at the margins of the monsoon
Scientific Reports (2021)
-
Widespread Denisovan ancestry in Island Southeast Asia but no evidence of substantial super-archaic hominin admixture
Nature Ecology & Evolution (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
