
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) is a frequent cause of difficult-to-treat, often fatal infections in humans1,2. Most humans have antibodies against S. aureus, but these are highly variable and often not protective in immunocompromised patients3. Previous vaccine development programs have not been successful4. A large percentage of human antibodies against S. aureus target wall teichoic acid (WTA), a ribitol-phosphate (RboP) surface polymer modified with N-acetylglucosamine (GlcNAc)5,6. It is currently unknown whether the immune evasion capacities of MRSA are due to variation of dominant surface epitopes such as those associated with WTA. Here we show that a considerable proportion of the prominent healthcare-associated and livestock-associated MRSA clones CC5 and CC398, respectively, contain prophages that encode an alternative WTA glycosyltransferase. This enzyme, TarP, transfers GlcNAc to a different hydroxyl group of the WTA RboP than the standard enzyme TarS7, with important consequences for immune recognition. TarP-glycosylated WTA elicits 7.5–40-fold lower levels of immunoglobulin G in mice than TarS-modified WTA. Consistent with this, human sera contained only low levels of antibodies against TarP-modified WTA. Notably, mice immunized with TarS-modified WTA were not protected against infection with tarP-expressing MRSA, indicating that TarP is crucial for the capacity of S. aureus to evade host defences. High-resolution structural analyses of TarP bound to WTA components and uridine diphosphate GlcNAc (UDP-GlcNAc) explain the mechanism of altered RboP glycosylation and form a template for targeted inhibition of TarP. Our study reveals an immune evasion strategy of S. aureus based on averting the immunogenicity of its dominant glycoantigen WTA. These results will help with the identification of invariant S. aureus vaccine antigens and may enable the development of TarP inhibitors as a new strategy for rendering MRSA susceptible to human host defences.
Similar content being viewed by others


Main
Novel prevention and treatment strategies against major antibiotic-resistant pathogens such as MRSA are urgently needed but are not within reach because some of the most critical virulence strategies of these pathogens are not understood8. The pathogenic potential of prominent healthcare-associated (HA)-MRSA and recently emerged livestock-associated (LA)-MRSA strains is thought to rely on particularly effective immune evasion strategies, whereas community-associated (CA)-MRSA strains often produce more aggressive toxins1,2. Most humans have high overall levels of antibodies against S. aureus as a consequence of preceding infections, but antibody titres differ strongly for specific antigens and are often not protective in immunocompromised patients, for reasons that are not clear3. A large percentage of human antibodies against S. aureus is directed against WTA5,9,10, which is largely invariant. However, some S. aureus lineages produce altered WTA, which modulates, for instance, phage susceptibility7,11.
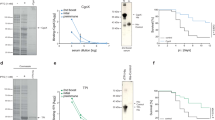
To investigate whether some prevalent S. aureus lineages use additional WTA-targeted strategies to increase their fitness and pathogenicity, we screened S. aureus genomes for potential additional paralogues of WTA biosynthesis genes. We found three S. aureus prophages that encoded a protein, TarP, that has 27% identity to the WTA-β-GlcNAc transferase TarS7 (Fig. 1a). tarP was found exclusively in isolates of the prominent HA-MRSA CC512, on a prophage that also encoded the scn, chp and sak immune evasion genes13, and on two other prophages in the emerging LA-MRSAs CC39814 and CC515. All tarP-harbouring genomes also contained tarS.

a, TarP is encoded next to different integrase types (int gene) in prophages φtarP-Sa3int (with immune evasion cluster scn, chp, sak, sep), found in HA-MRSA, and φtarP-Sa1int and φtarP-Sa9int, identified in LA-MRSA. TarP variants in φtarP-Sa1int and φtarP-Sa9int differed from TarP in φtarP-Sa3int in one amino acid each (I8M and D296N, respectively). Both residues are distant from the catalytic centre. b, Complementation of S. aureus RN4420 ΔtarM/S with either tarS or tarP restores susceptibility to infection by WTA GlcNAc-binding siphophages, as indicated by plaque formation on bacterial lawns. Data shown are representative of three independent experiments. c, tarP expression reduces siphophage Φ11-mediated transfer of SaPIbov in N315. Values indicate the ratio of transduction units (TrU) to plaque-forming units (PFU) given as mean ± s.d. of three independent experiments. Statistical significances when compared to wild type were calculated by one-way ANOVA with Dunnett’s post-test (two-sided) and significant P values (P ≤ 0.05) are indicated. NO (none obtained) indicates no obtained transductants.
When tarP from CC5 HA-MRSA strain N315 was expressed in a WTA glycosylation-deficient mutant of laboratory strain RN42207, it restored WTA glycosylation (Extended Data Fig. 1a) and susceptibility to siphophages, which need RboP WTA GlcNAc as a binding motif16 (Fig. 1b). The presence of β-GlcNAc on WTA is essential for full β-lactam resistance in MRSA strains7. When tarP was expressed in a WTA glycosylation-deficient mutant of CA-MRSA strain MW2 (CC1), it restored full oxacillin resistance (Extended Data Fig. 1b), confirming that tarP can replace tarS in several key interactions.
The expression of TarP led to susceptibility to siphophages, albeit to a lower extent than TarS (Extended Data Fig. 1c), although TarP did not incorporate less GlcNAc into WTA than TarS (Extended Data Fig. 1d, Supplementary Table 3). Similarly, the siphophage-mediated horizontal transfer of an S. aureus pathogenicity island was reduced about tenfold in S. aureus N315 expressing tarP, compared to the same strain expressing only tarS (Fig. 1c), suggesting that TarP and TarS glycosylate WTA differently. Notably, N315 was resistant to podophages, but inactivation of tarP (but not of tarS) rendered it susceptible to podophages (Fig. 2a). We analysed the overall effect of tarP on podophage susceptibility patterns in 90 clinical CC5 and CC398 isolates and found that none of the tarP-containing strains, but all of the tarP-lacking strains, were susceptible to podophages (Extended Data Table 1). Thus, TarP causes podophage resistance and TarP-mediated modification of WTA is distinct from that mediated by TarS. Nuclear magnetic resonance (NMR) analyses revealed that both TarP and TarS add GlcNAc to WTA in the β-configuration. However, the attachment site in RboP differs: TarS glycosylates the C4 position17 whereas TarP attaches GlcNAc to C3 (Fig. 2b, Extended Data Fig. 2, Supplementary Table 2). This difference may be crucial for impairing phage infection. Moreover, NMR analysis revealed that TarP is dominant over TarS because in N315, which bears both genes, GlcNAc was almost exclusively attached to RboP C3 (Fig. 2b).

a, Expression of tarP renders N315 resistant to podophages. Representative data from three independent experiments are shown. b, 1H NMR spectra reveal different ribitol hydroxyl glycosylation of N315 WTA by TarS (C4) or TarP (C3). The RboP units with attached GlcNAc are depicted above the corresponding proton resonances. Representative data from three experiments are shown. In-depth description of the structural motifs identified in the spectra is given in the Supplementary Information. c, Crystal structure of TarP homotrimer (pink, orange, grey) bound to UDP-GlcNAc (yellow) and two Mn2+ ions (lime green). The nucleotide-binding domain (NBD), acceptor-binding domain (ABD), and C-terminal trimerization domain (CTD) of the pink monomer are labelled. d, Views into the trimer interface (boxed in c). Left, polar interactions. Hydrogen bonds and salt bridges are shown as black dashed lines. The Mn2+ is 2.1 Å from each Asp316 carboxylate. Right, hydrophobic interactions, with the mutated residue Ile322 highlighted in red. e, Size-exclusion chromatography elution profiles. Based on calibration of the column, the TarP wild-type and I322E mutant proteins have estimated molecular weights of 138 kDa (n = 8) and 42 kDa (n = 3), respectively, in agreement with the calculated molecular weights of 120 kDa for a TarP trimer and 40 kDa for monomeric TarP.
We solved the TarP structure at high resolution to elucidate how TarP generates a different glycosylation product from TarS. Like TarS18, TarP forms stable homotrimers, but it uses a different trimerization strategy because it lacks the C-terminal trimerization domain found in TarS (Fig. 2c, Extended Data Fig. 3). Instead, hydrophobic and polar interactions of a small helical C-terminal domain generate the TarP trimer (Fig. 2d, e). WTA polymers comprising three or six RboP repeating units (3RboP or 6RboP-(CH2)6NH2, respectively) were synthesized and used for soaking TarP crystals (Supplementary Information Fig. 2, 3), yielding the first protein structure visualizing the binding of a WTA-based polymer (Fig. 3, Extended Data Fig. 4). In the ternary complex TarP–UDP-GlcNAc–3RboP, the distance between the C3-hydroxyl of the third unit of 3RboP (RboP3) and the anomeric C1 of GlcNAc is 4.2 Å. Furthermore, at 3.1 Å, Asp181 is well within hydrogen bonding distance of the C3-hydroxyl of RboP3. The observed distances and geometry nicely explain the unusual glycosylation of WTA at the C3-hydroxyl. We propose that TarP uses a direct SN2-like glycosyltransferase reaction, as discussed for other inverting GT-A fold enzymes19,20. In this mechanism, Asp181 would act as the catalytic base, deprotonating the C3-hydroxyl on RboP3 and enabling a nucleophilic attack on the GlcNAc C1, thus yielding a β-O-GlcNAcylated polyRboP (Fig. 3c). Mutagenesis of Asp181 to alanine rendered TarP inactive, supporting this putative mechanism (Extended Data Table 2).

a, 3RboP binding site in the TarP–3RboP complex, with key amino acids shown (cyan). Asp181 is highlighted in red. The ribitol of 3RboP is coloured green and d-ribitol-5-phosphate units 1, 2 and 3 (RboP1, RboP2, and RboP3) are labelled. Hydrogen bonds and salt bridges are shown as black dashed lines. b, Ternary complex of TarP with UDP-GlcNAc and 3RboP. UDP-GlcNAc, Mg2+ and 3RboP are shown as full-atom models coloured yellow, magenta, and green, respectively. c, View into the active site of TarP. C1 of UDP-GlcNAc and Asp181 are highlighted with red labels. The arrow indicates how the C3-hydroxyl in RboP3 could nucleophilically attack GlcNAc C1. d, Comparison of the polyRboP-binding site of TarP with the corresponding region in TarS. Residues of TarP and 3RboP are coloured as in a. TarS residues are coloured violet and the two sulfates are labelled S1 and S2. Only residues of TarP are labelled, for clarity. Key TarP and TarS residues lining the polyRboP-binding site are shown at the bottom, with three identical (red) and one conserved amino acids (blue). e, Superposition of UDP-GlcNAc-bound TarS with the ternary TarP–UDP-GlcNAc–3RboP complex. UDP-GlcNAc and 3RboP bound to TarP are coloured as in b, whereas UDP-GlcNAc bound to TarS is coloured in cyan. Only the TarS residues are shown (coloured as in d), for clarity. The arrows indicate the C1 positions of UDP-GlcNAc in TarP and TarS.
The ternary structure of TarP–UDP-GlcNAc–3RboP allows us to predict how polyRboP binds to the homologous TarS enzyme. Three residues that are critical for binding and catalysis (including Asp181) are identical in TarP and TarS, while five other residues differ18 (Fig. 3d). Lys255 and Arg262, for instance, which interact electrostatically with a WTA phosphate group in TarP, are replaced with Glu248 and Ser255, respectively, in TarS, which may lead to reduced affinity for WTA and might explain why TarP is dominant over TarS in vivo. On the basis of the location of UDP-GlcNAc, the identical Tyr149, Asp178 and Arg252 side chains, the conserved aromatic side chain of Phe256, and a site that contains a bound sulfate ion from the crystallization solution (S1) and probably binds phosphate in TarS (Fig. 3e), the polyRboP chain would be shifted to the upper right, and the relative position of RboP units in the binding site would be altered in TarS. Such an altered binding mode would move the C4-hydroxyl of the target RboP towards C1 of GlcNAc, allowing TarS to glycosylate at the C4 position.
S. aureus WTA is a dominant antigen for adaptive immune responses5,9. The observation that the position of GlcNAc on RboP had a profound impact on binding by podophage receptors raises the question of whether human antibodies also discriminate between the two isomeric polymers and whether MRSA clones use TarP to subvert immune recognition. We analysed several human antibody preparations for their capacity to opsonize a panel of N315 strains with or without tarP and/or tarS. The mutant lacking any WTA glycosylation bound the lowest amount of IgG compared to WTA glycosylation-positive strains (Fig. 4a), demonstrating that glycosylated WTA is a prominent S. aureus antigen in humans. Exclusive expression of tarS led to strongly increased IgG binding compared to the glycosylation-deficient mutant, indicating that β-GlcNAc on RboP C4 is an important epitope for human anti-S. aureus antibodies. By contrast, expression of tarP in the presence or absence of tarS led to only slightly increased IgG binding compared to the glycosylation-deficient mutant. The capacity of TarP to impair the deposition of IgG on S. aureus differed with individual serum donors and reached average levels in pooled serum preparations (Fig. 4a). When tarP was deleted in three further CC5 isolates, they showed similarly increased capacities to bind human serum antibodies compared to the wild-type strains (Extended Data Fig. 1e). Additionally, tarP deletion led to a substantially increased capacity of human neutrophils to phagocytose opsonized S. aureus (Fig. 4b, Extended Data Fig. 1g). Thus, only a small percentage of S. aureus-specific antibodies can bind WTA with β-GlcNAc on RboP C3, and tarP-expressing S. aureus are less likely to be detected and eliminated by human phagocytes.

a, TarP expression reduces deposition of IgG from human serum on N315 cells. The protein A gene spa was deleted in all strains. Top, human IgG isolated from three individual healthy donors (A, B, and C; n = 4); bottom, left, IgG from human serum enriched for RN4220 WTA binding (n = 4); middle and right, pooled human IgG from different suppliers (Abcam, n = 4; Athens R&T, n = 6). Results were normalized against wild type and shown as means with s.d. of n experiments. P values for comparison with wild type were calculated by one-way ANOVA with Dunnett’s post-test (two-sided), and P ≤ 0.05 was considered significant. Significant P values are displayed. b, TarP reduces neutrophil phagocytosis of N315 strains lacking protein A, opsonized with indicated concentrations of IgG enriched for WTA binding. Values are depicted as mean fluorescence intensity (MFI). Means of two dependent replicates of a representative experiment are shown. The other two representative experiments can be found in Extended Data Fig. 1g. c, TarP abrogates IgG response of mice towards WTA. For each experiment, WTA from N315 ΔtarP or ΔtarS was isolated independently. At least three mice per group were vaccinated and analysed for specific IgG at indicated time points after vaccination. Results are depicted as mean absorbance with s.d. Individual mice are indicated by colour. Increase in IgG levels was assessed by one-way ANOVA with Tukey’s post-test (two-sided). Significant differences (P ≤ 0.05) are indicated with corresponding P values. d, Vaccination with WTA does not protect mice against tarP-expressing N315, as shown for bacterial loads in kidney upon intravenous infection. No significant differences between groups of either five vaccinated mice or four mice for the alum control group (means indicated), calculated by one-way ANOVA, were observed.
We purified N315 WTA that had been glycosylated by TarS or TarP and used it to immunize mice. Antibodies binding to regular (TarS-modified) WTA increased continuously over three weeks after vaccination (Fig. 4c). By contrast, no or only very low amounts of IgG directed against TarP-glycosylated WTA emerged, indicating that WTA modified at RboP C3 is much less immunogenic than WTA modified at RboP C4. This experiment was repeated three times with three different WTA preparations and yielded broadly similar data.
Vaccination with S. aureus WTA bearing GlcNAc at RboP C4 protects mice against infection by CA-MRSA strains USA300 (CC8) or USA400 (CC1), which both lack tarP5,21. Remarkably, vaccination with regular (TarS-modified) or TarP-modified WTA did not lead to any notable protection against subsequent infection with tarP-expressing N315 compared to mock vaccination, despite the robust antibody response against regular WTA (Fig. 4d). Together, our results demonstrate that tarP protects S. aureus against adaptive host defences by allowing bacteria to evade recognition by preexisting anti-S. aureus antibodies and by exploiting the poor immunogenicity of TarP-modified WTA.
It is possible that TarP-modified WTA mimics a currently unknown autoantigen and is therefore hardly immunogenic. On the other hand, regular S. aureus WTA can be ingested by antigen-presenting cells and presented to T cells, in a largely unexplored way, thereby evoking specific immunoglobulins and immunological memory22,23. It is possible that TarP-modified WTA is refractory to this process. Thus, TarS- and TarP-modified WTA could be helpful for decoding glycopolymer presentation pathways and for defining the most promising WTA epitopes for the development of protective vaccines against S. aureus.
Protection against S. aureus infections is urgently needed, in particular for hospitalized and immunocompromised patients2,4. Antibodies can in principle protect against S. aureus, but their titres and specificities vary largely among humans and they are often not protective in immunocompromised patients3, probably in particular against S. aureus clones that mask dominant epitopes, for instance using TarP. Unfortunately, all previous human vaccination attempts with protein or glycopolymer antigens have failed, for reasons that are unclear24. Our study identifies a new strategy used by pandemic MRSA clones to subvert antibody-mediated immunity, which should be considered in future vaccination approaches. S. aureus WTA with GlcNAc at RboP C3 has been reported as a type-336 antigen, but was not further explored25. We found that tarP is present in type-336 S. aureus (Extended Data Fig. 1f). However, TarP-modified WTA is a very poor antigen and vaccines directed against GlcNAc at WTA RboP C3 or C4 may fail against many of the pandemic MRSA clones. The structural characterization of TarP will instruct the development of specific TarP inhibitors that could become important in combination with anti-WTA vaccines or antibiotic therapies. We found tarP-encoding prophages in 70–80% of south-west German HA-MRSA CC5 and 40% of Danish LA-MRSA CC398 isolates (Extended Data Table 1), pointing to a crucial role of tarP in the fitness of these lineages and raising concerns of further dissemination by horizontal gene transfer. TarP is a new and probably crucial component of the S. aureus virulence factor arsenal26,27, highlighting the important roles of adaptive immunity and its evasion in S. aureus infections.
Methods
No statistical methods were used to predetermine sample size. The experiments were not randomized. The investigators were not blinded to allocation during experiments and outcome assessment.
Bacterial strains and growth conditions
S. aureus strains N315, RN4220, and MW2 (wild type and mutants) were used for this study. Collections of CC5 isolates of the Rhine-Hesse pulsed-field gel electrophoresis type28 and of the LA-MRSA lineage CC398 from the Danish Statens Serum Institut29,30 were analysed for the presence of tarP and for podophage susceptibility. Additionally, 48 spa-type t002 (ST5) and 16 spa-type t003 (ST225) isolates were obtained from the MRSA collection of the University Hospital Tübingen and analysed for tarP presence by PCR. S. aureus strains were cultivated in tryptic soy broth (TSB) or basic medium (BM; 1% tryptone, 0.5% yeast extract, 0.5% NaCl, 0.1% glucose, 0.1% K2HPO4, w/v). MICs of oxacillin were determined by microbroth dilution according to established guidelines31.
Experiments with phages
tarP-encoding phages were identified in genome sequences using the webtool Phaster32 in representative strains listed with GenBank accession: ΦtarP-Sa3int with immune evasion cluster (IEC) in CC5 (strain N315, BA000018.3), ΦtarP-Sa1int, found in LA-MRSA of CC5 (strain ISU935, CP017090), and ΦtarP-Sa9int found in CC398 (strain E154, CP013218).
Phage susceptibility was determined using a soft-agar overlay method16. In brief, 10 µl phage lysate of 104–106 PFU was dropped onto soft agar containing 100 µl bacterial suspension (OD600 of 0.1). Plates were incubated at 37 °C overnight. The efficiency of plating was determined as described33. Transfer of SaPIs was determined according to previously described methods11. In brief, SaPI particle lysates were generated from S. aureus strain JP1794, which encodes a SaPI with a resistance marker for tetracycline34. PFU of SaPI lysate was determined on RN4220. 200 µl bacterial culture (OD600 of 0.5) was mixed with 100 µl of SaPI particle lysate (SaPIbov1 (Φ11), 106 PFU/ml), incubated at 37 °C for 15 min. Appropriate dilutions were plated on TSB plates containing 3 µg/ml of tetracycline, and CFU were checked after overnight incubation.
WTA isolation and structure analysis
WTA from S. aureus was isolated and purified according to previously described methods11. In brief, WTA was released from purified peptidoglycan by treatment with 5% trichloroacetic acid and dialysed extensively against water using a Spectra/Por3 dialysis membrane (MWCO of 3.5 kDa; VWR International GmbH). Obtained soluble WTA was quantified by determining the content of phosphate35 and GlcNAc36. For PAGE analysis of WTA, samples (400 nmol of phosphate per lane) were applied to a 26% polyacrylamide (Rotiphorese Gel 40 (19:1)) resolving gel and separated at 25 mA for 16 h37. The gel was equilibrated in a solution of 40% ethanol and 5% acidic acid at room temperature for 1 h and the WTA ladders were visualized by incubation with alcian blue (0.005%) for several hours.
NMR spectroscopy experiments were carried out on a Bruker DRX-600 spectrometer equipped with a cryo-probe, at 288 K (WT-WTA, TarS-WTA, and TarP-WTA) or 298 K (double-mutant WTA lacking any glycosylation). Chemical shifts of spectra recorded in D2O were calculated in p.p.m. relative to internal acetone (2.225 and 31.45 p.p.m.). The spectral width was set to 10 p.p.m. and the frequency carrier placed at the residual HOD peak, suppressed by pre-saturation. Two-dimensional spectra (TOCSY, gHSQC, gHMBC, and HSQC-TOCSY) were measured using standard Bruker software. For all experiments, 512 FIDs of 2,048 complex data points were collected, 32 scans per FID were acquired for homonuclear spectra, and 20 or 100 ms of mixing time was used for TOCSY spectra. Heteronuclear 1H-13C spectra were measured in the 1H-detected mode, gHSQC spectrum was acquired with 40 scans per FID, the GARP sequence was used for 13C decoupling during acquisition; gHMBC scans doubled those of gHSQC spectrum. As for HSQC-TOCSY, the multiplicity editing during selection step version was used, scans tripled those of the HSQC spectrum and two experiments were acquired by setting the mixing time to 20 or 80 ms. During processing, each data matrix was zero-filled in both dimensions to give a matrix of 4K × 2K points and was resolution-enhanced in both dimensions by a cosine-bell function before Fourier transformation; data processing and analysis were performed with the Bruker Topspin 3 program.
Molecular biology
All primers used for PCR, cloning, and mutagenesis are listed in Supplementary Table 1. tarP (UniProt A0A0H3JNB0, NCBI Gene ID 1260584) was amplified using genomic DNA of S. aureus N315 and inserted in Eschericha coli/S. aureus shuttle vector pRB47438 at the BamHI and SacI sites, to transform S. aureus, or into pQE80L at BamH1 and HindIII sites, to transform E. coli BL21(DE3). A thrombin cleavage site was inserted between the His-tag and mature protein in pQE80L. Single mutations of TarP were introduced by PCR-based site-directed mutagenesis39. The obtained amplicons were confirmed by sequencing. For the construction of marker-less S. aureus deletion mutants of tarS or tarP, the pIMAY shuttle vector was used40. The IgG-binding surface protein A gene (spa) was deleted using the pKORI shuttle vector41. Protein A deletion had no impact on phage siphophage or podophage susceptibility, indicating that it did not alter WTA amount or structure.
Protein expression, purification, and activity assay
E. coli BL21(DE3) were grown in LB medium at 30 °C. Expression of tarP was induced with 1 mM IPTG at 22 °C at an OD600 of 0.6. After 15 h, cells were harvested, washed with wash buffer (50 mM Tris-HCl, pH 8.0, 1 mM EDTA), and lysed by sonication with lysis buffer (70 mM NaH2PO4, pH 8.0, 1 M NaCl, 20% glycerol, 10 U/ml of benzonase nuclease). After centrifugation (15,000g), the supernatant was filtered with a 0.45 µm filter, loaded onto a His Trap FF column (GE Healthcare, 5 ml), and washed with buffer A (50 mM NaH2PO4, pH 8.0, 1 M NaCl, 20% glycerol) supplemented with 45 mM imidazole and buffer B (buffer A with 90 mM imidazole). Finally, the protein was eluted with buffer C (buffer A with 500 mM imidazole), and the fractions were pooled, and further purified by size-exclusion chromatography on a Superdex 200 10/30 column equilibrated with buffer D (20 mM MOPS, pH 7.6, 400 mM LiCl, 10 mM MgCl2, 5 mM β-mercaptoethanol, 5% glycerol). The peak fractions were pooled and concentrated to 1.4 mg/ml for crystallization. For selenomethionyl-form TarP production, bacteria were grown in a selenomethionine-containing medium (Molecular Dimension) and auto-induction was carried out. The protein was purified as described above. The activity of wild-type and mutated TarP, as well as donor substrate specificity of TarP were determined with the ADP Quest Assay kit (DiscoverRx, Extended Data Tables 2, 3). The reaction volume was 20 µl with 1 mM UDP-GlcNAc, 1.5 mM purified WTA from RN4220 ΔtarM/S. The reaction was started with protein and incubated at room temperature for 1 h. Released UDP, coupled into a fluorescence signal, was detected in a 384-well black assay plate with 530 nm excitation and 590 nm emission wavelengths using TECAN Infinite M200.
Crystallization and data collection
Crystals were obtained by vapour diffusion at 20 °C. 1 µl protein solution was mixed with 1 µl reservoir solution containing 25% PEG 3350, 250 mM MgCl2, and 0.1 M sodium citrate, pH 5.7. The selenomethionyl-form protein was crystallized under the same conditions. For crystals of TarP with UDP-GlcNAc, 27 mM UDP-GlcNAc was introduced in the reservoir solution containing 250 mM MgCl2 or 230 mM MnCl2. Crystals of TarP with Mg2+ were used for soaking of synthetic 3RboP (60 mM), 6RboP-(CH2)6NH2 (41 mM), or UDP-GlcNAc (20 mM) combined with 3RboP (52 mM) for 5 min. For data collection the crystals were cryo-protected with 20% glycerol in reservoir solution and flash-frozen in liquid nitrogen. Diffraction data were collected at beamline X06DA of Swiss Light Source in Villigen, Switzerland, or at beamline BL14.1 at BESSY-II, Helmholtz Zentrum Berlin.
Phasing, model building, and refinement
For phase determination, two data sets from a selenomethionine-containing TarP crystal were collected at wavelengths of 0.97941 Å (peak) and 0.97952 Å (inflection). The structure was solved by multi-wavelength anomalous dispersion (MAD) at 2.60 Å resolution. All data were reduced using XDS/XSCALE software packages42. Initial phases were derived from the substructure of 26 selenium atom sites per asymmetric unit with the program suite SHELX C/D/E43. The heavy atom parameters were further refined and the initial phases were improved by SHARP/autoSHARP44. The initial model was generated with PHENIX45 and the final model was achieved by cycles of iterative model modification using COOT46, and restrained refinement with REFMAC. TLS was used in the later stages47,48. The four binary and one ternary complex structures were solved by molecular replacement using PHASER49 and the unliganded TarP structure was used as a search model. UDP-GlcNAc, 3RboP, Mg2+, or Mn2+ were removed from the models to calculate the simulated annealing (mFo − DFc) omit maps using PHENIX. The anomalous difference map of Mn2+ at 1.89259 Ǻ was generated by FFT within CCP4, from which two Mn2+ in the active site and one Mn2+ at the trimer interface were identified. The coordinate and parameter files for 3RboP and 6RboP-(CH2)6NH2 were calculated using the PRODRG server50. The structure figures were generated by PyMOL51 and the models were evaluated using MolProbity52. Statistics for the data collection, phasing, and refinement are reported in Extended Data Tables 4 and 5.
Synthesis of ribitol phosphate oligomers
Synthesis of 3RboP
Target compound 1, d-ribitol-5-phosphate trimer (3RboP), was prepared by the phosphoramidite method53,54 (Supplementary Fig. 2). In brief, the primary alcohol of commercially available compound 2 was converted into levulinoyl ester by using levulinic acid and N,N'-dicyclohexylcarbodiimide (DCC), and the allyl group of 3 was removed with tetrakis(triphenylphosphine)palladium to produce compound 4. The primary alcohol of 4 reacted with phosphine derivative 5 in the presence of diisopropylammonium tetrazolide55 to generate phosphoramidite 6. At the same time, compound 4 was coupled with dibenzyl N,N-diisopropylphosphoramidite 7, which was catalysed by 1H-tetrazole, and the product was further oxidized by tert-butyl hydroperoxide, yielding protected d-ribitol-5-phosphate 8. Cleavage of the levulinoyl ester of 8 with hydrazine hydrate resulted in benzyl protected d-ribitol-5-phosphate 9, which was further coupled with phosphoramidite 6 and oxidized with tert-butyl hydroperoxide to yield protected dimers of d-ribitol-5-phosphate 10. After removal of the levulinoyl group, the dimer 11 was coupled with phosphoramidite 6 using the same conditions as above to obtain a protected trimer of d-ribitol-5-phosphate 12. Subsequent removal of the levulinoyl group and hydrogenolysis of 13 to remove all benzyl groups yielded 3RboP 1. All chemicals and experimental procedures as well as characterization of products can be found in the Supplementary Methods.
Synthesis of 6RboP-(CH2)6NH2
Aminohexyl d-ribitol-5-phosphate hexamer (6RboP-(CH2)6NH2) was synthesized using a new method (Supplementary Fig. 3). All chemicals (Acros, Biosolve, Sigma-Aldrich and TCI) for the synthesis were used as received and all reactions were performed under a protective argon atmosphere at room temperature, unless otherwise stated. Procedures for phosphoramidite coupling, oxidation, detritylation, global deprotection, TLC analysis and characterization of these compounds can be found in Supplementary Methods.
Human samples
Venous blood samples were obtained from male and female healthy volunteers (20–50 years) with protocols approved by the Institutional Review Board for Human Subjects at the University of Tübingen (014/2014BO2 und 549/2018BO2). Informed written consent was obtained from all volunteers. Blood samples were used for purification of either serum IgGs or neutrophils as described below.
IgG from human plasma
IgG was purified from plasma of human donors using the NAb Protein G Spin Kit (ThermoFisher), purity was checked by SDS PAGE, and protein concentration was determined using Bradford assay. Anti-WTA-IgG was prepared as described9. To analyse the IgG-binding capacity of S. aureus cells, exponentially growing bacterial cultures were adjusted to an OD600 of 0.5, diluted 1:10 in PBS, and 100 µl of diluted bacteria was mixed with 100 µl of IgG diluted in PBS with 1% BSA. The concentration of IgG was 250 ng/ml for IgG enriched for WTA binding, 10 µg/ml for IgG from pooled human serum (Athens R&T 16-16-090707, Abcam ab98981), or 5 µg/ml for single-donor IgG preparations. A control without IgG was included in all experiments for all mutants. Samples were incubated at 4 °C for 1 h, centrifuged, washed 2–3 times with PBS, and further incubated with 100 µl FITC-labelled anti-human IgG (Thermo Scientific, 62-8411, 1:100 in PBS with 1% BSA, 62-8411) at 4 °C for 1 h. Bacteria were centrifuged, washed 2–3 times with PBS, and fixed with 2% paraformaldehyde (PFA). Surface-bound IgG was quantified by flow cytometry using a BD FACSCalibur. For all flow cytometry experiments a mutant panel lacking spa, the gene for the IgG-binding protein A, was used. The subsequent gating strategy is exemplified in Extended Data Fig. 5a.
IgG-mediated phagocytosis
Stationary-phase S. aureus cells were washed once with PBS and labelled by incubation in PBS containing 10 µM carboxyfluorescein succinimidyl ester (CFSE; OD600 of 1.7) at 37 °C for 1 h. The bacteria were washed three times and resuspended in PBS. CFU were determined by plating on TSB plates and bacteria were heat-inactivated at 70 °C for 20 min. CFSE-labelled S. aureus (1 × 107 cells/ml) in PBS with 0.5% BSA were opsonized with anti-WTA-IgG (0.15 or 0.3 ng/µl) at 4 °C for 40 min. Neutrophils from human donors, isolated via Ficoll-Histopaque density gradient centrifugation56, were diluted to a concentration of 2.5 × 106/ml in neutrophil medium (10% HSA, 2 mM l-glutamine, 2 mM sodium pyruvate, 10 mM HEPES). 200 µl neutrophil suspension was incubated with 25 µl opsonized bacteria (final MOI 0.5) in a 96-well plate at 37 °C for 30 min, centrifuged (350g, 10 min), washed once with 200 µl PBS, and fixed with 2% PFA at room temperature for 15 min. Cells were washed twice with PBS and analysed by flow cytometry, whereby surface-bound and ingested bacteria were measured without discrimination. An example of the neutrophil gating strategy can be found in Extended Data Fig. 5b.
Mice
Six-week-old sex-matched wild-type C57BL/6J mice, purchased from ORIENT BIO (Charles River Breeding Laboratories in Korea), were kept in micro-isolator cages in a pathogen-free animal facility. The conducted experiments were performed according to guidelines and approval (PNU-2017-1503) by the Pusan National University-Institutional Animal Care and Use Committee (PNU-IACUC). Sample size was chosen to obtain significant outcomes (alpha error ≤ 5%), based on results from previous experiments21. Experiments were performed in a non-blinded, non-randomized fashion.
Mouse vaccination and infection
30 µg of purified WTA from S. aureus N315 wild-type or isogenic ΔtarP, or ΔtarS mutants was dissolved in 15 µl PBS and mixed with the same volume of aluminium hydroxide gel adjuvant (Alhydrogelr 1.3%, 6.5 mg/ml, Brennatag). The mixtures were incubated at 37 °C with agitation for 1 h and injected three times at one-week intervals via mouse footpads. Seven days after the third injection, blood was obtained from the retro-orbital sinus and centrifuged (9,000g) at 4 °C for 10 min. The supernatants were aliquoted (50 µl) and stored at –80 °C for ELISA quantification of WTA-binding IgG as described57. Sera were diluted 1:100 and tested by ELISA on 96-well plates coated with 2.5 µg/ml sonicated WTA preparations (WTA from N315, ΔtarS or ΔtarP, respectively).
To prepare an inoculum for infection, N315 wild-type bacteria were grown in TBS at 37 °C with agitation (180 r.p.m.) until they reached an OD600 of 1.0. After centrifugation (3,500g) at 4 °C for 10 min, bacteria adjusted to 5 × 107 CFU in 50 µl PBS containing 0.01% BSA were intravenously injected (n = 5 per group). Injected bacterial numbers were verified by plating serial dilutions of the inoculum onto TSA plates. To determine residual bacterial dissemination to kidneys, challenged mice were euthanized, and organs were extracted aseptically and homogenized in 1 ml of saline using a Polytron homogenizer (PT3100). The homogenates were serially diluted and plated on TSA to determine CFU counts. CFU were calculated per 1 ml of kidney.
Statistical analyses
Statistical analysis was performed by using GraphPad Prism (GraphPad Software, Inc.). Statistically significant differences were calculated by appropriate statistical methods as indicated. P values of ≤ 0.05 were considered significant.
Reporting summary
Further information on experimental design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
All major data generated or analysed in this study are included in the article or its supplementary information files. The coordinates and structure factors were deposited in the Protein Data Bank under accession numbers 6H1J, 6H21, 6H2N, 6H4F, 6H4M and 6HNQ. Source data for experiments with animals (Fig. 4c, d) are provided. Additionally, a gel image of Extended Data Fig. 1f is supplied as Supplementary Fig. 1. All other data relating to this study are available from the corresponding authors on reasonable request.
References
Tong, S. Y., Davis, J. S., Eichenberger, E., Holland, T. L. & Fowler, V. G. Jr. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 28, 603–661 (2015).
Lee, A. S. et al. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 4, 18033 (2018).
Stentzel, S. et al. Specific serum IgG at diagnosis of Staphylococcus aureus bloodstream invasion is correlated with disease progression. J. Proteomics 128, 1–7 (2015).
Missiakas, D. & Schneewind, O. Staphylococcus aureus vaccines: deviating from the carol. J. Exp. Med. 213, 1645–1653 (2016).
Lehar, S. M. et al. Novel antibody–antibiotic conjugate eliminates intracellular S. aureus. Nature 527, 323–328 (2015).
Weidenmaier, C. & Peschel, A. Teichoic acids and related cell-wall glycopolymers in Gram-positive physiology and host interactions. Nat. Rev. Microbiol. 6, 276–287 (2008).
Brown, S. et al. Methicillin resistance in Staphylococcus aureus requires glycosylated wall teichoic acids. Proc. Natl Acad. Sci. USA 109, 18909–18914 (2012).
Tacconelli, E. et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327 (2018).
Kurokawa, K. et al. Glycoepitopes of staphylococcal wall teichoic acid govern complement-mediated opsonophagocytosis via human serum antibody and mannose-binding lectin. J. Biol. Chem. 288, 30956–30968 (2013).
Lee, J. H. et al. Surface glycopolymers are crucial for in vitro anti-wall teichoic acid IgG-mediated complement activation and opsonophagocytosis of Staphylococcus aureus. Infect. Immun. 83, 4247–4255 (2015).
Winstel, V. et al. Wall teichoic acid structure governs horizontal gene transfer between major bacterial pathogens. Nat. Commun. 4, 2345 (2013).
Nübel, U. et al. Frequent emergence and limited geographic dispersal of methicillin-resistant Staphylococcus aureus. Proc. Natl Acad. Sci. USA 105, 14130–14135 (2008).
McCarthy, A. J. & Lindsay, J. A. Staphylococcus aureus innate immune evasion is lineage-specific: a bioinfomatics study. Infect. Genet. Evol. 19, 7–14 (2013).
Bal, A. M. et al. Genomic insights into the emergence and spread of international clones of healthcare-, community- and livestock-associated meticillin-resistant Staphylococcus aureus: Blurring of the traditional definitions. J. Glob. Antimicrob. Resist. 6, 95–101 (2016).
Hau, S. J., Bayles, D. O., Alt, D. P., Frana, T. S. & Nicholson, T. L. Draft genome sequences of 63 swine-associated methicillin-resistant Staphylococcus aureus sequence type 5 isolates from the United States. Genome Announc. 5, e01081-17 (2017).
Xia, G. et al. Wall teichoic acid-dependent adsorption of staphylococcal siphovirus and myovirus. J. Bacteriol. 193, 4006–4009 (2011).
Vinogradov, E., Sadovskaya, I., Li, J. & Jabbouri, S. Structural elucidation of the extracellular and cell-wall teichoic acids of Staphylococcus aureus MN8m, a biofilm forming strain. Carbohydr. Res. 341, 738–743 (2006).
Sobhanifar, S. et al. Structure and mechanism of Staphylococcus aureus TarS, the wall teichoic acid β-glycosyltransferase involved in methicillin resistance. PLoS Pathog. 12, e1006067 (2016).
Lairson, L. L., Henrissat, B., Davies, G. J. & Withers, S. G. Glycosyltransferases: structures, functions, and mechanisms. Annu. Rev. Biochem. 77, 521–555 (2008).
Kozmon, S. & Tvaroska, I. Catalytic mechanism of glycosyltransferases: hybrid quantum mechanical/molecular mechanical study of the inverting N-acetylglucosaminyltransferase I. J. Am. Chem. Soc. 128, 16921–16927 (2006).
Takahashi, K. et al. Intradermal immunization with wall teichoic acid (WTA) elicits and augments an anti-WTA IgG response that protects mice from methicillin-resistant Staphylococcus aureus infection independent of mannose-binding lectin status. PLoS One 8, e69739 (2013).
Weidenmaier, C., McLoughlin, R. M. & Lee, J. C. The zwitterionic cell wall teichoic acid of Staphylococcus aureus provokes skin abscesses in mice by a novel CD4+ T-cell-dependent mechanism. PLoS One 5, e13227 (2010).
Wanner, S. et al. Wall teichoic acids mediate increased virulence in Staphylococcus aureus. Nat. Microbiol. 2, 16257 (2017).
Pozzi, C. et al. Vaccines for Staphylococcus aureus and target populations. Curr. Top. Microbiol. Immunol. 409, 491–528 (2017).
Fattom, A., Sarwar, J., Kossaczka, Z., Taylor, K. & Ennifar, S. Method of protecting against staphylococcal infection. US Patent US20060228368A1 (2006).
Thammavongsa, V., Kim, H. K., Missiakas, D. & Schneewind, O. Staphylococcal manipulation of host immune responses. Nat. Rev. Microbiol. 13, 529–543 (2015).
Spaan, A. N., Surewaard, B. G., Nijland, R. & van Strijp, J. A. Neutrophils versus Staphylococcus aureus: a biological tug of war. Annu. Rev. Microbiol. 67, 629–650 (2013).
Schulte, B., Bierbaum, G., Pohl, K., Goerke, C. & Wolz, C. Diversification of clonal complex 5 methicillin-resistant Staphylococcus aureus strains (Rhine-Hesse clone) within Germany. J. Clin. Microbiol. 51, 212–216 (2013).
Larsen, J. et al. Meticillin-resistant Staphylococcus aureus CC398 is an increasing cause of disease in people with no livestock contact in Denmark, 1999 to 2011. Euro Surveill. 20, 30021 (2015).
Sieber, R. N. et al. Drivers and dynamics of methicillin-resistant livestock-associated Staphylococcus aureus CC398 in pigs and humans in Denmark. mBio 9, e02142-18 (2018).
European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 9, ix–xv (2003).
Arndt, D. et al. PHASTER: a better, faster version of the PHAST phage search tool. Nucleic Acids Res. 44, W16–W21 (2016).
Winstel, V., Sanchez-Carballo, P., Holst, O., Xia, G. & Peschel, A. Biosynthesis of the unique wall teichoic acid of Staphylococcus aureus lineage ST395. MBio 5, e00869 (2014).
Tormo, M. A. et al. Staphylococcus aureus pathogenicity island DNA is packaged in particles composed of phage proteins. J. Bacteriol. 190, 2434–2440 (2008).
Chen, P. S., Toribara, T. Y. & Warner, H. Microdetermination of phosphorus. Anal. Chem. 28, 1756–1758 (1956).
Smith, R. L. & Gilkerson, E. Quantitation of glycosaminoglycan hexosamine using 3-methyl-2-benzothiazolone hydrazone hydrochloride. Anal. Biochem. 98, 478–480 (1979).
Xia, G. et al. Glycosylation of wall teichoic acid in Staphylococcus aureus by TarM. J. Biol. Chem. 285, 13405–13415 (2010).
Brückner, R. A series of shuttle vectors for Bacillus subtilis and Escherichia coli. Gene 122, 187–192 (1992).
Liu, H. & Naismith, J. H. An efficient one-step site-directed deletion, insertion, single and multiple-site plasmid mutagenesis protocol. BMC Biotechnol. 8, 91 (2008).
Monk, I. R., Shah, I. M., Xu, M., Tan, M. W. & Foster, T. J. Transforming the untransformable: application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. MBio 3, e00277-11 (2012).
Bae, T. & Schneewind, O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 55, 58–63 (2006).
Kabsch, W. Xds. Acta Crystallogr. D Biol. Crystallogr. 66, 125–132 (2010).
Sheldrick, G. M. Experimental phasing with SHELXC/D/E: combining chain tracing with density modification. Acta Crystallogr. D Biol. Crystallogr. 66, 479–485 (2010).
Vonrhein, C., Blanc, E., Roversi, P. & Bricogne, G. Automated structure solution with autoSHARP. Methods Mol. Biol. 364, 215–230 (2007).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66, 213–221 (2010).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Murshudov, G. N. et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D Biol. Crystallogr. 67, 355–367 (2011).
Murshudov, G. N., Vagin, A. A. & Dodson, E. J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 53, 240–255 (1997).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Schüttelkopf, A. W. & van Aalten, D. M. PRODRG: a tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. D Biol. Crystallogr. 60, 1355–1363 (2004).
Schrodinger, LLC. The PyMOL Molecular Graphics System, Version 1.8 (2015).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12–21 (2010).
Beaucage, S. L. & Caruthers, M. H. Deoxynucleoside phosphoramidites—a new class of key intermediates for deoxypolynucleotide synthesis. Tetrahedr. Lett. 22, 1859–1862 (1981).
Elie, C. J. J. et al. Synthesis of fragments of the capsular polysaccharide of Haemophilus influenzae type b: Part IIII-3. A solid-phase synthesis of a spacer-containing ribosylribitol phosphate hexamer. Recl. Trav. Chim. Pays Bas 108, 219–223 (1989).
Dreef, C. E., Elie, C. J. J., Hoogerhout, P., van der Marel, G. A. & van Boom, J. H. Synthesis of 1-O-(1,2-di-O-palmitoyl-sn-glycero-3-phospho)-d-myo-inositol 4,5-bisphosphate: an analogue of naturally occurring (ptd)Ins(4,5)P2. Tetrahedr. Lett. 29, 6513–6515 (1988).
Dürr, M. C. et al. Neutrophil chemotaxis by pathogen-associated molecular patterns—formylated peptides are crucial but not the sole neutrophil attractants produced by Staphylococcus aureus. Cell. Microbiol. 8, 207–217 (2006).
Caulfield, M. J. et al. Small molecule mimetics of an HIV-1 gp41 fusion intermediate as vaccine leads. J. Biol. Chem. 285, 40604–40611 (2010).
Acknowledgements
We thank S. Popovich and P. Kühner for technical assistance; E. Weiß for help with phagocytosis experiments; R. Rosenstein and X. Li for discussions; B. Blaum and G. Zocher for assistance with NMR analysis and support for structure phasing and discussion; and the Swiss Lightsource beamline staff of the Paul Scherrer Institute for beam time and technical support. This work was financed by grants from the German Research Foundation to A.P. (TRR34, CRC766, TRR156, RTG1708), T.S. (TRR34, CRC766), C.W. (TRR34, CRC766, TRR156, RTG1708), and G.X. (CRC766); the German Center of Infection Research to A.P. (HAARBI); the Ministry of Science and Technology, Thailand Government to W.S.; the Korean Drug Development Foundation to S.-H.K. and B.L.L. (KDDF-201703-1); and the Max-Planck-Society to P.H.S.
Reviewer information
Nature thanks M. Crispin, F. DeLeo, M. Gilmore and J. Zimmer for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
D.G. characterized TarP in vivo and its genomic context, created mutants, designed experiments, purified WTA, and performed experiments with human IgGs. Y.G. designed experiments, purified proteins, crystallized proteins, solved the structures, and performed in vitro analysis of TarP. C.D.C. performed NMR experiments. C.D.C. and A.M. analysed the NMR data and wrote the NMR discussion. S.-H.K. performed and B.L.L. designed and interpreted mouse immunization and infection experiments. K.S. designed IgG deposition experiments. B.S. and C.W. collected and characterized CC5 MRSA strains. J.L. collected and characterized CC398 strains. J.L. and C.W. analysed S. aureus genomes. F.-F.X, C.P., and P.H.S. designed and synthesized 3RboP. S.A. and J.C. designed and synthesized 6RboP-(CH2)6NH2. W.S. performed MIC experiments. G.X. identified tarP, and characterized and interpreted MIC data. D.G., Y.G., A.P., T.S., and G.X. designed the study, analysed results, and wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Characterization of TarP, deposition of human IgGs, and presence of tarP in the producer of antigen 336.
a, Analysis of WTA by PAGE. WTA from RN4220 ΔtarM/S expressing either tarP or tarS was compared with non-glycosylated WTA. Data shown are representative of two experiments. b, MIC values of oxacillin against MW2 wild type, tarS mutant, and tarP-complemented tarS mutant. Data are medians of ten independent experiments. c, Efficiency of plating (EOP) of phage Φ11 against tarS or tarP-expressing RN4420 ΔtarM/S. Values of tarP relative to tarS expression are given as mean ± s.d. (n = 3). Statistical significance was calculated by paired Student’s t-test (two-sided) with significant P values (P ≤ 0.05) indicated. d, The level of WTA glycosylation catalysed by TarP or TarS was determined by analysing the GlcNAc and phosphate content of WTA isolated from a N315 strain panel. Depicted is the ratio of GlcNAc and phosphate as mean with s.d. of three technical replicates. The values are in good agreement with NMR data (Supplementary Table 3). e, Relative deposition of IgG from intravenous immunoglobulins enriched for WTA binding on different CC5 wild-type and tarP mutant cells. Values are given as mean percentage ± s.d. of four independent experiments. Statistical significance was calculated by paired Student’s t-test (two-sided). P values ≤ 0.05 were considered significant and are indicated. f, Presence of tarP and tarS in S. aureus ATCC55804, expressing antigen 336, described as 3-O-GlcNAc-WTA25. Shown is a representative of two independent replicates. g, TarP reduces neutrophil phagocytosis of N315 strains lacking protein A, opsonized with indicated concentrations of IgG enriched for WTA binding. Values are depicted as mean fluorescence intensity (MFI). Shown are two independent experiments with neutrophils from different donors. These data supplement data presented in Fig. 4b: upper panel, mean of three technical replicates of an independent experiment, lower panel, mean of two technical replicates.
Extended Data Fig. 2 NMR analysis of WTA from N315 mutant panel.
All depicted experiments were repeated twice. y-axes and x-axes show 13C and 1H chemical shifts, respectively. a–d, NMR spectra of non-glycosylated WTA (ΔtarSΔtarP mutant). a, HSQC expansion of the region containing the ribitol and glycerol protons shifted by acylation; b, c, HSQC-TOCSY-20 and HSQC-TOCSY-80 spectra, respectively. d, HSQC area of the non-acylated ribitol and glycerol proton. e–h, NMR spectra of TarS-WTA (ΔtarP mutant). e, HSQC expansion of the region containing the ribitol and glycerol protons shifted by acylation. f, g, HSQC-TOCSY-20 and HSQC-TOCSY-80, respectively. h, HSQC area of the non-acylated ribitol and glycerol proton. i–o, NMR spectra of TarP-WTA (ΔtarS mutant). i, HSQC expansion of the region containing the ribitol and glycerol protons shifted by acylation. j, k, HSQC-TOCSY-20 and HSQC-TOCSY-80 spectra, respectively. l, HSQC area of the non-acylated ribitol and glycerol protons. m, Expansion of l with HSQC (black/grey) overlapped with HSQC-TOCSY-20 (cyan). n, Overlap of HSQC-TOCSY-20 (cyan) and HSQC-TOCSY-80 (black). o, HSQC (black) and HMBC (grey) detailing the GlcNAc signals. p, NOESY expansion detailing the correlations of the β-GlcNAc anomeric protons: GlcNAc ‘b*’ differs from unit ‘b’, which has the same anomeric proton chemical shift, but is linked to a different ribitol unit. All densities are labelled with the letters used in Supplementary Table 2. The density marked with an asterisk in m is consistent with ribitol glycosylated at O-4.
Extended Data Fig. 3 Secondary structure of a TarP monomer and interactions with UDP-GlcNAc.
a, Cartoon representation of a TarP monomer bound to UDP-GlcNAc (yellow) and Mn2+ (lime green). The CTD is coloured red. b, Interactions of TarP with UDP-GlcNAc and Mn2+, coloured as in a. Hydrogen bonds and salt bridges are shown as black dashed lines. c, Interactions of TarP with UDP-GlcNAc (yellow) and Mg2+ (magenta). d, Simulated-annealing (mFo − DFc) omit map of UDP-GlcNAc (grey mesh, contoured at 2.0σ) and Mn2+ (magenta mesh, at 3.0σ) in the TarP–UDP-GlcNAc–Mn2+ complex structure. UDP-GlcNAc and Mn2+ are coloured as in a. e, Simulated-annealing (mFo − DFc) omit map of UDP-GlcNAc (grey mesh, at 2.0σ) and Mg2+ (blue mesh, at 2.0σ) in the TarP–UDP-GlcNAc–Mg2+ complex structure. UDP-GlcNAc and Mg2+ are coloured as in c.
Extended Data Fig. 4 Simulated-annealing (mFo − DFc) omit maps of 3RboP and UDP-GlcNAc, and characterization of TarP mutant proteins.
a, Chemical structures of synthetic 3RboP and 6RboP-(CH2)6NH2. The unit numbers are indicated. b, Simulated-annealing (mFo − DFc) omit map of 3RboP (lime green) in the binary structure (magenta mesh, contoured at 2.0σ). c, Simulated-annealing (mFo − DFc) omit map of UDP-GlcNAc (yellow), Mg2+ (magenta) and 3RboP (lime green) in the ternary complex structure (red mesh, at 1.8σ, blue mesh, at 2.0σ or magenta mesh, at 1.5σ). d, Circular dichroism spectra of wild-type and mutant TarP proteins (for wild type, R76A and D181A, n = 3; for D92A, Y152A and R259A, n = 2). e, Size-exclusion chromatography elution profiles of wild-type and mutant TarP proteins (for wild type, n = 8; for R76A, D181A and R259A, n = 3; for D92A and Y152A, n = 2, all with similar results). Mutant proteins D94A, E180A, D209A, K255A, R262A, and H263A showed similar circular dichroism spectra and size-exclusion chromatography elution profiles (data not shown).
Extended Data Fig. 5 Gating strategy for flow cytometry experiments.
a, Gating strategy for IgG deposition experiments. To distinguish bacteria from background signals, pure PBS was measured. Left, bacterial gating occurred at the FSC/SCC density plot omitting PBS-derived signals. Bacterial aggregates of high SSC and FSC values were excluded from the gated population as well. Right, the mean fluorescence of the bacterial population (black) was determined and compared with non-IgG-treated bacteria (grey) to control for nonspecific binding of the secondary FITC-labelled antibody. Subsequently, mean fluorescence values of individual mutants were compared relatively to the corresponding wild-type strain. b, Gating strategy for phagocytosis experiments. Neutrophils were separated by Histopaque/Ficoll gradient and subsequent gating of neutrophils occurred at the FSC/SCC density plot upon size and complexity (left). Histopaque/Ficoll gradient isolations showed a neutrophil purity of more than 80%. Using the CFSE-fluorescence channel, the gated population was subdivided into fluorescence-positive and -negative cells (right). Successful phagocytosis was indicated by uptake of CFSE-labelled bacteria. The phagocytic efficiency was expressed as product of the mean fluorescence of the fluorescence-positive population and their relative abundance (mean fluorescence intensity, MFI).
Supplementary information
Supplementary Figure 1
Photo of gel electrophoresis for Extended Data Fig. 1f
Supplementary Information
This file contains Supplementary Information Sections 1-3, including a Supplementary Discussion, Supplementary Tables 1-3, Supplementary Figures 2- 3, NMR Spectra data and Supplementary References
Source data
Rights and permissions
About this article

Cite this article
Gerlach, D., Guo, Y., De Castro, C. et al. Methicillin-resistant Staphylococcus aureus alters cell wall glycosylation to evade immunity. Nature 563, 705–709 (2018). https://doi.org/10.1038/s41586-018-0730-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0730-x
Keywords
This article is cited by
-
Clinical characteristics and homology analysis of Staphylococcus aureus from would infection at a tertiary hospital in southern Zhejiang, China
BMC Microbiology (2023)
-
A multi-enzyme machine polymerizes the Haemophilus influenzae type b capsule
Nature Chemical Biology (2023)
-
Van-mediated self-aggregating photothermal agents combined with multifunctional magnetic nickel oxide nanoparticles for precise elimination of bacterial infections
Journal of Nanobiotechnology (2022)
-
The crafty opponent: the defense systems of Staphylococcus aureus and response measures
Folia Microbiologica (2022)
-
Platelet membrane-camouflaged silver metal-organic framework drug system against infections caused by methicillin-resistant Staphylococcus aureus
Journal of Nanobiotechnology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
