
Abstract
The environment, diet, microbiota and body’s metabolism shape complex biological processes in health and disease. However, our understanding of the molecular pathways involved in these processes is still limited. The aryl hydrocarbon receptor (AHR) is a ligand-activated transcription factor that integrates environmental, dietary, microbial and metabolic cues to control complex transcriptional programmes in a ligand-specific, cell-type-specific and context-specific manner. In this Review, we summarize our current knowledge of AHR and the transcriptional programmes it controls in the immune system. Finally, we discuss the role of AHR in autoimmune and neoplastic diseases of the central nervous system, with a special focus on the gut immune system, the gut–brain axis and the therapeutic potential of targeting AHR in neurological disorders.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

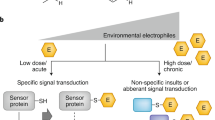
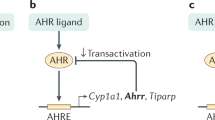
References
McIntosh, B. E., Hogenesch, J. B. & Bradfield, C. A. Mammalian Per-Arnt-Sim proteins in environmental adaptation. Annu. Rev. Physiol. 72, 625–645 (2010).
Fukunaga, B. N., Probst, M. R., Reisz-Porszasz, S. & Hankinson, O. Identification of functional domains of the aryl hydrocarbon receptor. J. Biol. Chem. 270, 29270–29278 (1995).
Antonsson, C., Whitelaw, M. L., McGuire, J., Gustafsson, J. A. & Poellinger, L. Distinct roles of the molecular chaperone hsp90 in modulating dioxin receptor function via the basic helix-loop-helix and PAS domains. Mol. Cell. Biol. 15, 756–765 (1995).
Stockinger, B., Di Meglio, P., Gialitakis, M. & Duarte, J. H. The aryl hydrocarbon receptor: multitasking in the immune system. Annu. Rev. Immunol. 32, 403–432 (2014).
Kudo, I. et al. The regulation mechanisms of AhR by molecular chaperone complex. J. Biochem. 163, 223–232 (2018).
Carver, L. A. & Bradfield, C. A. Ligand-dependent interaction of the aryl hydrocarbon receptor with a novel immunophilin homolog in vivo. J. Biol. Chem. 272, 11452–11456 (1997).
Meyer, B. K. & Perdew, G. H. Characterization of the AhR-hsp90-XAP2 core complex and the role of the immunophilin-related protein XAP2 in AhR stabilization. Biochemistry 38, 8907–8917 (1999).
Meyer, B. K., Pray-Grant, M. G., Vanden Heuvel, J. P. & Perdew, G. H. Hepatitis B virus X-associated protein 2 is a subunit of the unliganded aryl hydrocarbon receptor core complex and exhibits transcriptional enhancer activity. Mol. Cell. Biol. 18, 978–988 (1998).
Petrulis, J. R. & Perdew, G. H. The role of chaperone proteins in the aryl hydrocarbon receptor core complex. Chem. Biol. Interact. 141, 25–40 (2002).
Petrulis, J. R., Kusnadi, A., Ramadoss, P., Hollingshead, B. & Perdew, G. H. The hsp90 co-chaperone XAP2 alters importin beta recognition of the bipartite nuclear localization signal of the Ah receptor and represses transcriptional activity. J. Biol. Chem. 278, 2677–2685 (2003).
Ramadoss, P., Petrulis, J. R., Hollingshead, B. D., Kusnadi, A. & Perdew, G. H. Divergent roles of hepatitis B virus X-associated protein 2 (XAP2) in human versus mouse Ah receptor complexes. Biochemistry 43, 700–709 (2004).
Pappas, B. et al. p23 protects the human aryl hydrocarbon receptor from degradation via a heat shock protein 90-independent mechanism. Biochem. Pharmacol. 152, 34–44 (2018).
Kazlauskas, A., Poellinger, L. & Pongratz, I. Evidence that the co-chaperone p23 regulates ligand responsiveness of the dioxin (Aryl hydrocarbon) receptor. J. Biol. Chem. 274, 13519–13524 (1999).
Enan, E. & Matsumura, F. Identification of c-Src as the integral component of the cytosolic Ah receptor complex, transducing the signal of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) through the protein phosphorylation pathway. Biochem. Pharmacol. 52, 1599–1612 (1996).
Enan, E. & Matsumura, F. Evidence for a second pathway in the action mechanism of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Significance of Ah-receptor mediated activation of protein kinase under cell-free conditions. Biochem. Pharmacol. 49, 249–261 (1995).
Hwang, S. J. et al. Indoxyl 3-sulfate stimulates Th17 differentiation enhancing phosphorylation of c-Src and STAT3 to worsen experimental autoimmune encephalomyelitis. Toxicol. Lett. 220, 109–117 (2013).
Backlund, M. & Ingelman-Sundberg, M. Regulation of aryl hydrocarbon receptor signal transduction by protein tyrosine kinases. Cell Signal. 17, 39–48 (2005).
Pongratz, I., Mason, G. G. & Poellinger, L. Dual roles of the 90-kDa heat shock protein hsp90 in modulating functional activities of the dioxin receptor. Evidence that the dioxin receptor functionally belongs to a subclass of nuclear receptors which require hsp90 both for ligand binding activity and repression of intrinsic DNA binding activity. J. Biol. Chem. 267, 13728–13734 (1992).
Ikuta, T. et al. Nucleocytoplasmic shuttling of the aryl hydrocarbon receptor. J. Biochem. 127, 503–509 (2000).
Ikuta, T., Kobayashi, Y. & Kawajiri, K. Phosphorylation of nuclear localization signal inhibits the ligand-dependent nuclear import of aryl hydrocarbon receptor. Biochem. Biophys. Res. Commun. 317, 545–550 (2004).
Tsuji, N. et al. The activation mechanism of the aryl hydrocarbon receptor (AhR) by molecular chaperone HSP90. FEBS Open Bio 4, 796–803 (2014).
Yao, E. F. & Denison, M. S. DNA sequence determinants for binding of transformed Ah receptor to a dioxin-responsive enhancer. Biochemistry 31, 5060–5067 (1992).
Swanson, H. I., Tullis, K. & Denison, M. S. Binding of transformed Ah receptor complex to a dioxin responsive transcriptional enhancer: evidence for two distinct heteromeric DNA-binding forms. Biochemistry 32, 12841–12849 (1993).
Sakurai, S., Shimizu, T. & Ohto, U. The crystal structure of the AhRR-ARNT heterodimer reveals the structural basis of the repression of AhR-mediated transcription. J. Biol. Chem. 292, 17609–17616 (2017).
Mascanfroni, I. D. et al. Metabolic control of type 1 regulatory T cell differentiation by AHR and HIF1-alpha. Nat. Med. 21, 638–646 (2015). This study shows that AHR and HIF1α collaborate to integrate molecular cues from the cellular microenvironment to shape T cell polarization.
Ohtake, F. et al. Modulation of oestrogen receptor signalling by association with the activated dioxin receptor. Nature 423, 545–550 (2003). This paper shows that AHR interacts with the oestrogen receptor to target non-XRE DNA elements.
Hankinson, O. Role of coactivators in transcriptional activation by the aryl hydrocarbon receptor. Arch. Biochem. Biophys. 433, 379–386 (2005).
Yeste, A. et al. Tolerogenic nanoparticles inhibit T cell-mediated autoimmunity through SOCS2. Sci. Signal. 9, ra61 (2016).
McBerry, C., Gonzalez, R. M., Shryock, N., Dias, A. & Aliberti, J. SOCS2-induced proteasome-dependent TRAF6 degradation: a common anti-inflammatory pathway for control of innate immune responses. PLOS ONE 7, e38384 (2012). This paper is the first to identify that AHR and SOCS2 interact to control NF-κB activation.
Wilson, S. R., Joshi, A. D. & Elferink, C. J. The tumor suppressor Kruppel-like factor 6 is a novel aryl hydrocarbon receptor DNA binding partner. J. Pharmacol. Exp. Ther. 345, 419–429 (2013).
Ohtake, F. et al. Dioxin receptor is a ligand-dependent E3 ubiquitin ligase. Nature 446, 562–566 (2007).
Mejia-Garcia, A. et al. Activation of AHR mediates the ubiquitination and proteasome degradation of c-Fos through the induction of Ubcm4 gene expression. Toxicology 337, 47–57 (2015).
Bunaciu, R. P. & Yen, A. Activation of the aryl hydrocarbon receptor AhR promotes retinoic acid-induced differentiation of myeloblastic leukemia cells by restricting expression of the stem cell transcription factor Oct4. Cancer Res. 71, 2371–2380 (2011).
Luecke-Johansson, S. et al. A molecular mechanism to switch the aryl hydrocarbon receptor from a transcription factor to an E3 ubiquitin ligase. Mol. Cell. Biol. 37, e00630–16 (2017).
Vogel, C. F. et al. RelB, a new partner of aryl hydrocarbon receptor-mediated transcription. Mol. Endocrinol. 21, 2941–2955 (2007).
Salisbury, R. L. & Sulentic, C. E. The AhR and NF-kappaB/Rel proteins mediate the inhibitory effect of 2,3,7,8-tetrachlorodibenzo-p-dioxin on the 3' immunoglobulin heavy chain regulatory region. Toxicol. Sci. 148, 443–459 (2015).
Vogel, C. F. et al. Cross-talk between aryl hydrocarbon receptor and the inflammatory response: a role for nuclear factor-kappaB. J. Biol. Chem. 289, 1866–1875 (2014).
Iu, M. et al. RelB attenuates cigarette smoke extract-induced apoptosis in association with transcriptional regulation of the aryl hydrocarbon receptor. Free Radic. Biol. Med. 108, 19–31 (2017).
Kimura, A., Naka, T., Nohara, K., Fujii-Kuriyama, Y. & Kishimoto, T. Aryl hydrocarbon receptor regulates Stat1 activation and participates in the development of Th17 cells. Proc. Natl Acad. Sci. USA 105, 9721–9726 (2008).
Quintana, F. J. et al. An endogenous aryl hydrocarbon receptor ligand acts on dendritic cells and T cells to suppress experimental autoimmune encephalomyelitis. Proc. Natl Acad. Sci. USA 107, 20768–20773 (2010).
Veldhoen, M., Hirota, K., Christensen, J., O’Garra, A. & Stockinger, B. Natural agonists for aryl hydrocarbon receptor in culture medium are essential for optimal differentiation of Th17 T cells. J. Exp. Med. 206, 43–49 (2009).
Tian, Y., Ke, S., Chen, M. & Sheng, T. Interactions between the aryl hydrocarbon receptor and P-TEFb. Sequential recruitment of transcription factors and differential phosphorylation of C-terminal domain of RNA polymerase II at Cyp1a1 promoter. J. Biol. Chem. 278, 44041–44048 (2003).
Wang, S., Ge, K., Roeder, R. G. & Hankinson, O. Role of mediator in transcriptional activation by the aryl hydrocarbon receptor. J. Biol. Chem. 279, 13593–13600 (2004).
Beischlag, T. V. et al. Recruitment of the NCoA/SRC-1/p160 family of transcriptional coactivators by the aryl hydrocarbon receptor/aryl hydrocarbon receptor nuclear translocator complex. Mol. Cell. Biol. 22, 4319–4333 (2002).
Wang, S. & Hankinson, O. Functional involvement of the Brahma/SWI2-related gene 1 protein in cytochrome P4501A1 transcription mediated by the aryl hydrocarbon receptor complex. J. Biol. Chem. 277, 11821–11827 (2002).
Schnekenburger, M., Peng, L. & Puga, A. HDAC1 bound to the Cyp1a1 promoter blocks histone acetylation associated with Ah receptor-mediated trans-activation. Biochim. Biophys. Acta 1769, 569–578 (2007).
Chang, C. C., Sue, Y. M., Yang, N. J., Lee, Y. H. & Juan, S. H. 3-Methylcholanthrene, an AhR agonist, caused cell-cycle arrest by histone deacetylation through a RhoA-dependent recruitment of HDAC1 and pRb2 to E2F1 complex. PLOS ONE 9, e92793 (2014).
Liu, K. Y., Wang, L. T. & Hsu, S. H. Modification of epigenetic histone acetylation in hepatocellular carcinoma. Cancers 10, 8 (2018).
Grimaldi, G., Rajendra, S. & Matthews, J. The aryl hydrocarbon receptor regulates the expression of TIPARP and its cis long non-coding RNA, TIPARP-AS1. Biochem. Biophys. Res. Commun. 495, 2356–2362 (2018).
Garcia, G. R. et al. In vivo characterization of an AHR-dependent long noncoding RNA required for proper Sox9b expression. Mol. Pharmacol. 91, 609–619 (2017).
Liu, C. C. et al. Micro124-mediated AHR expression regulates the inflammatory response of chronic rhinosinusitis (CRS) with nasal polyps. Biochem. Biophys. Res. Commun. 500, 145–151 (2018).
Rogers, S. et al. Aryl hydrocarbon receptor (AhR)-dependent regulation of pulmonary miRNA by chronic cigarette smoke exposure. Sci. Rep. 7, 40539 (2017).
Morales-Hernandez, A. et al. Alu retrotransposons promote differentiation of human carcinoma cells through the aryl hydrocarbon receptor. Nucleic Acids Res. 44, 4665–4683 (2016).
Roman, A. C. et al. Dioxin receptor and SLUG transcription factors regulate the insulator activity of B1 SINE retrotransposons via an RNA polymerase switch. Genome Res. 21, 422–432 (2011).
Gutierrez-Vazquez, C. & Quintana, F. J. Regulation of the immune response by the aryl hydrocarbon receptor. Immunity 48, 19–33 (2018).
Bessede, A. et al. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature 511, 184–190 (2014). This paper shows that endotoxin tolerance following repeated exposure to bacterial lipopolysaccharide is mediated by AHR.
Matsumura, F. The significance of the nongenomic pathway in mediating inflammatory signaling of the dioxin-activated Ah receptor to cause toxic effects. Biochem. Pharmacol. 77, 608–626 (2009).
Han, Z. et al. Aryl hydrocarbon receptor mediates laminar fluid shear stress-induced CYP1A1 activation and cell cycle arrest in vascular endothelial cells. Cardiovasc. Res. 77, 809–818 (2008).
Xiao, W., Son, J., Vorrink, S. U., Domann, F. E. & Goswami, P. C. Ligand-independent activation of aryl hydrocarbon receptor signaling in PCB3-quinone treated HaCaT human keratinocytes. Toxicol. Lett. 233, 258–266 (2015).
Murray, I. A. et al. Evidence that ligand binding is a key determinant of Ah receptor-mediated transcriptional activity. Arch. Biochem. Biophys. 442, 59–71 (2005).
Lee, C. C. et al. Ligand independent aryl hydrocarbon receptor inhibits lung cancer cell invasion by degradation of Smad4. Cancer Lett. 376, 211–217 (2016).
Ilchmann, A. et al. Impact of culture medium on maturation of bone marrow-derived murine dendritic cells via the aryl hydrocarbon receptor. Mol. Immunol. 51, 42–50 (2012).
Oberg, M., Bergander, L., Hakansson, H., Rannug, U. & Rannug, A. Identification of the tryptophan photoproduct 6-formylindolo[3,2-b]carbazole, in cell culture medium, as a factor that controls the background aryl hydrocarbon receptor activity. Toxicol. Sci. 85, 935–943 (2005).
Denison, M. S. & Nagy, S. R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 43, 309–334 (2003). This is an outstanding review on AHR agonists focusing on exogenous AHR ligands.
Okey, A. B. An aryl hydrocarbon receptor odyssey to the shores of toxicology: the Deichmann Lecture, International Congress of Toxicology-XI. Toxicol. Sci. 98, 5–38 (2007).
Kafafi, S. A., Afeefy, H. Y., Said, H. K. & Kafafi, A. G. Relationship between aryl hydrocarbon receptor binding, induction of aryl hydrocarbon hydroxylase and 7-ethoxyresorufin O-deethylase enzymes, and toxic activities of aromatic xenobiotics in animals. A new model. Chem. Res. Toxicol. 6, 328–334 (1993).
Kerger, B. D. et al. Age- and concentration-dependent elimination half-life of 2,3,7,8-tetrachlorodibenzo-p-dioxin in Seveso children. Environ. Health Perspect. 114, 1596–1602 (2006).
Kopec, A. K. et al. Toxicogenomic evaluation of long-term hepatic effects of TCDD in immature, ovariectomized C57BL/6 mice. Toxicol. Sci. 135, 465–475 (2013).
Loub, W. D., Wattenberg, L. W. & Davis, D. W. Aryl hydrocarbon hydroxylase induction in rat tissues by naturally occurring indoles of cruciferous plants. J. Natl Cancer Inst. 54, 985–988 (1975).
Song, J. et al. A ligand for the aryl hydrocarbon receptor isolated from lung. Proc. Natl Acad. Sci. USA 99, 14694–14699 (2002).
Seok, S. H. et al. Trace derivatives of kynurenine potently activate the aryl hydrocarbon receptor (AHR). J. Biol. Chem. 293, 1994–2005 (2018). This study reveals that TEACOPs are generated from kynurenine and activate AHR with higher affinity than their precursors.
Zelante, T. et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 39, 372–385 (2013). This paper identifies the tryptophan catabolites generated from the gut microbiota (L. reuteri) as potent AHR agonists relevant for mucosal immunity. It is one of the first studies to highlight the interaction of L. reuteri and the gut immune system.
Lamas, B. et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat. Med. 22, 598–605 (2016).
Natividad, J. M. et al. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab. 28, 737–749 (2018).
Wikoff, W. R. et al. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl Acad. Sci. USA 106, 3698–3703 (2009).
Yeste, A. et al. IL-21 induces IL-22 production in CD4+ T cells. Nat. Commun. 5, 3753 (2014).
Rothhammer, V. et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 22, 586–597 (2016).
Rothhammer, V. et al. Microglial control of astrocytes in response to microbial metabolites. Nature 557, 724–728 (2018). References 77 and 78 identify AHR in astrocytes and microglia as a participant of a gut–brain axis that controls CNS inflammation.
Muku, G. E., Murray, I. A., Espin, J. C. & Perdew, G. H. Urolithin A Is a dietary microbiota-derived human aryl hydrocarbon receptor antagonist. Metabolites 8, 86 (2018).
Poland, A. & Glover, E. Characterization and strain distribution pattern of the murine Ah receptor specified by the Ahd and Ahb-3 alleles. Mol. Pharmacol. 38, 306–312 (1990).
Okey, A. B., Vella, L. M. & Harper, P. A. Detection and characterization of a low affinity form of cytosolic Ah receptor in livers of mice nonresponsive to induction of cytochrome P1-450 by 3-methylcholanthrene. Mol. Pharmacol. 35, 823–830 (1989).
Harrill, J. A. et al. Knockout of the aryl hydrocarbon receptor results in distinct hepatic and renal phenotypes in rats and mice. Toxicol. Appl. Pharmacol. 272, 503–518 (2013).
Walisser, J. A., Bunger, M. K., Glover, E., Harstad, E. B. & Bradfield, C. A. Patent ductus venosus and dioxin resistance in mice harboring a hypomorphic Arnt allele. J. Biol. Chem. 279, 16326–16331 (2004).
Lawrence, B. P. et al. Activation of the aryl hydrocarbon receptor is essential for mediating the anti-inflammatory effects of a novel low-molecular-weight compound. Blood 112, 1158–1165 (2008).
Hauben, E. et al. Activation of the aryl hydrocarbon receptor promotes allograft-specific tolerance through direct and dendritic cell-mediated effects on regulatory T cells. Blood 112, 1214–1222 (2008).
Kerkvliet, N. I. et al. Activation of aryl hydrocarbon receptor by TCDD prevents diabetes in NOD mice and increases Foxp3+T cells in pancreatic lymph nodes. Immunotherapy 1, 539–547 (2009).
Veldhoen, M., Hocking, R. J., Atkins, C. J., Locksley, R. M. & Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006).
Korn, T. et al. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature 448, 484–487 (2007).
McGeachy, M. J. et al. TGF-beta and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain T(H)-17 cell-mediated pathology. Nat. Immunol. 8, 1390–1397 (2007).
McGeachy, M. J. GM-CSF: the secret weapon in the T(H)17 arsenal. Nat. Immunol. 12, 521–522 (2011).
Lee, Y. et al. Induction and molecular signature of pathogenic TH17 cells. Nat. Immunol. 13, 991–999 (2012).
Peters, A., Lee, Y. & Kuchroo, V. K. The many faces of Th17 cells. Curr. Opin. Immunol. 23, 702–706 (2011).
Ghoreschi, K. et al. Generation of pathogenic T(H)17 cells in the absence of TGF-beta signalling. Nature 467, 967–971 (2010).
Quintana, F. J. et al. Control of T(reg) and T(H)17 cell differentiation by the aryl hydrocarbon receptor. Nature 453, 65–71 (2008).
Veldhoen, M. et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 453, 106–109 (2008).
McAleer, J. P., Fan, J., Roar, B., Primerano, D. A. & Denvir, J. Cytokine regulation in human CD4 T cells by the aryl hydrocarbon receptor and Gq-coupled receptors. Sci. Rep. 8, 10954 (2018).
Ramirez, J. M. et al. Activation of the aryl hydrocarbon receptor reveals distinct requirements for IL-22 and IL-17 production by human T helper cells. Eur. J. Immunol. 40, 2450–2459 (2010).
Trifari, S., Kaplan, C. D., Tran, E. H., Crellin, N. K. & Spits, H. Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from T(H)-17, T(H)1 and T(H)2 cells. Nat. Immunol. 10, 864–871 (2009).
Talbot, J. et al. Smoking-induced aggravation of experimental arthritis is dependent of aryl hydrocarbon receptor activation in Th17 cells. Arthritis Res. Ther. 20, 119 (2018).
Quintana, F. J. et al. Aiolos promotes TH17 differentiation by directly silencing Il2 expression. Nat. Immunol. 13, 770–777 (2012).
Gagliani, N. et al. Th17 cells transdifferentiate into regulatory T cells during resolution of inflammation. Nature 523, 221–225 (2015).
Wei, P. et al. An aryl hydrocarbon receptor ligand acts on dendritic cells and T cells to suppress the Th17 response in allergic rhinitis patients. Lab. Invest. 94, 528–535 (2014).
Longhi, M. S. et al. Bilirubin suppresses Th17 immunity in colitis by upregulating CD39. JCI Insight 2, e92791 (2017).
Apetoh, L. et al. The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nat. Immunol. 11, 854–861 (2010).
Gandhi, R. et al. Activation of the aryl hydrocarbon receptor induces human type 1 regulatory T cell-like and Foxp3(+) regulatory T cells. Nat. Immunol. 11, 846–853 (2010). References 94, 95, 104 and 105 reveal the relevance of AHR signalling for T cell polarization and the molecular mechanisms involved.
Wu, H. Y. et al. In vivo induction of Tr1 cells via mucosal dendritic cells and AHR signaling. PLOS ONE 6, e23618 (2011).
Funatake, C. J., Marshall, N. B., Steppan, L. B., Mourich, D. V. & Kerkvliet, N. I. Cutting edge: activation of the aryl hydrocarbon receptor by 2,3,7,8-tetrachlorodibenzo-p-dioxin generates a population of CD4+CD25+ cells with characteristics of regulatory T cells. J. Immunol. 175, 4184–4188 (2005).
Singh, N. P. et al. Activation of aryl hydrocarbon receptor (AhR) leads to reciprocal epigenetic regulation of FoxP3 and IL-17 expression and amelioration of experimental colitis. PLOS ONE 6, e23522 (2011).
Joller, N. et al. Treg cells expressing the coinhibitory molecule TIGIT selectively inhibit proinflammatory Th1 and Th17 cell responses. Immunity 40, 569–581 (2014).
Stockinger, B., Veldhoen, M. & Hirota, K. Modulation of Th17 development and function by activation of the aryl hydrocarbon receptor—the role of endogenous ligands. Eur. J. Immunol. 39, 652–654 (2009).
Quintana, F. J. LeA(H)Rning self-control. Cell Res. 24, 1155–1156 (2014).
Quintana, F. J. & Sherr, D. H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 65, 1148–1161 (2013).
Matikainen, T. et al. Aromatic hydrocarbon receptor-driven Bax gene expression is required for premature ovarian failure caused by biohazardous environmental chemicals. Nat. Genet. 28, 355–360 (2001).
Quintana, F. J., Yeste, A. & Mascanfroni, I. D. Role and therapeutic value of dendritic cells in central nervous system autoimmunity. Cell Death Differ. 22, 215–224 (2015).
Chng, S. H. et al. Ablating the aryl hydrocarbon receptor (AhR) in CD11c+cells perturbs intestinal epithelium development and intestinal immunity. Sci. Rep. 6, 23820 (2016).
Ettmayer, P. et al. A novel low molecular weight inhibitor of dendritic cells and B cells blocks allergic inflammation. Am. J. Respir. Crit. Care Med. 173, 599–606 (2006).
Baba, N. et al. The aryl hydrocarbon receptor (AhR) ligand VAF347 selectively acts on monocytes and naive CD4(+) Th cells to promote the development of IL-22-secreting Th cells. Hum. Immunol. 73, 795–800 (2012).
Thordardottir, S. et al. The aryl hydrocarbon receptor antagonist StemRegenin 1 promotes human plasmacytoid and myeloid dendritic cell development from CD34+hematopoietic progenitor cells. Stem Cells Dev. 23, 955–967 (2014).
Goudot, C. et al. Aryl hydrocarbon receptor controls monocyte differentiation into dendritic cells versus macrophages. Immunity 47, 582–596 (2017).
Lu, L. et al. Critical role of all-trans retinoic acid in stabilizing human natural regulatory T cells under inflammatory conditions. Proc. Natl Acad. Sci. USA 111, E3432–E3440 (2014).
Mezrich, J. D. et al. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 185, 3190–3198 (2010).
Nguyen, N. T. et al. Aryl hydrocarbon receptor negatively regulates dendritic cell immunogenicity via a kynurenine-dependent mechanism. Proc. Natl Acad. Sci. USA 107, 19961–19966 (2010).
Vogel, C. F., Goth, S. R., Dong, B., Pessah, I. N. & Matsumura, F. Aryl hydrocarbon receptor signaling mediates expression of indoleamine 2,3-dioxygenase. Biochem. Biophys. Res. Commun. 375, 331–335 (2008).
Benson, J. M. & Shepherd, D. M. Dietary ligands of the aryl hydrocarbon receptor induce anti-inflammatory and immunoregulatory effects on murine dendritic cells. Toxicol. Sci. 124, 327–338 (2011).
Mascanfroni, I. D. et al. IL-27 acts on DCs to suppress the T cell response and autoimmunity by inducing expression of the immunoregulatory molecule CD39. Nat. Immunol. 14, 1054–1063 (2013).
Bankoti, J. et al. Effects of TCDD on the fate of naive dendritic cells. Toxicol. Sci. 115, 422–434 (2010).
Jin, U. H. et al. Microbiome-derived tryptophan metabolites and their aryl hydrocarbon receptor-dependent agonist and antagonist activities. Mol. Pharmacol. 85, 777–788 (2014).
Takamura, T. et al. Activation of the aryl hydrocarbon receptor pathway may ameliorate dextran sodium sulfate-induced colitis in mice. Immunol. Cell Biol. 88, 685–689 (2010).
Arsenescu, R. et al. Role of the xenobiotic receptor in inflammatory bowel disease. Inflamm. Bowel Dis. 17, 1149–1162 (2011).
Ikuta, T., Kurosumi, M., Yatsuoka, T. & Nishimura, Y. Tissue distribution of aryl hydrocarbon receptor in the intestine: Implication of putative roles in tumor suppression. Exp. Cell Res. 343, 126–134 (2016).
Monteleone, I. et al. Aryl hydrocarbon receptor-driven signals inhibit collagen synthesis in the gut. Eur. J. Immunol. 46, 1047–1057 (2016).
Metidji, A. et al. The environmental sensor AHR protects from inflammatory damage by maintaining intestinal stem cell homeostasis and barrier integrity. Immunity 49, 353–362 (2018). This paper shows that AHR in ISCs stabilizes gut epithelial barrier function and controls the regeneration of gut epithelial cells from their precursors.
Lanis, J. M. et al. Tryptophan metabolite activation of the aryl hydrocarbon receptor regulates IL-10 receptor expression on intestinal epithelia. Mucosal Immunol. 10, 1133–1144 (2017).
Yu, K. et al. AhR activation protects intestinal epithelial barrier function through regulation of Par-6. J. Mol. Histol. 49, 449–458 (2018).
Yu, M. et al. Aryl hydrocarbon receptor activation modulates intestinal epithelial barrier function by maintaining tight junction integrity. Int. J. Biol. Sci. 14, 69–77 (2018).
Fang, L. et al. Anti-TNF therapy induces CD4+T-cell production of IL-22 and promotes epithelial repairs in patients with Crohn’s disease. Inflamm. Bowel Dis. 24, 1733–1744 (2018).
Nikoopour, E., Bellemore, S. M. & Singh, B. IL-22, cell regeneration and autoimmunity. Cytokine 74, 35–42 (2015).
Fukumoto, S. et al. Identification of a probiotic bacteria-derived activator of the aryl hydrocarbon receptor that inhibits colitis. Immunol. Cell Biol. 92, 460–465 (2014).
Zhao, Y. et al. MicroRNA-124 promotes intestinal inflammation by targeting aryl hydrocarbon receptor in Crohn’s disease. J. Crohns Colitis 10, 703–712 (2016).
Iyer, S. S. et al. Dietary and microbial oxazoles induce intestinal inflammation by modulating aryl hydrocarbon receptor responses. Cell 173, 1123–1134 (2018). This paper identifies oxazoles as a novel class of AHR agonists from dietary and microbial sources with pro-inflammatory function in gut immunity.
Li, Y. et al. Exogenous stimuli maintain intraepithelial lymphocytes via aryl hydrocarbon receptor activation. Cell 147, 629–640 (2011).
Sujino, T. et al. Tissue adaptation of regulatory and intraepithelial CD4( + ) T cells controls gut inflammation. Science 352, 1581–1586 (2016).
Cervantes-Barragan, L. et al. Lactobacillus reuteri induces gut intraepithelial CD4(+)CD8alphaalpha(+) T cells. Science 357, 806–810 (2017). This is an elegant study reporting the control of immunoregulatory IELs by microbial metabolites via AHR.
Chen, W. et al. Aryl hydrocarbon receptor activation modulates CD8alphaalpha(+)TCRalphabeta(+) IELs and suppression of colitis manifestations in mice. Biomed. Pharmacother. 87, 127–134 (2017).
Bettelli, E. et al. Myelin oligodendrocyte glycoprotein-specific T cell receptor transgenic mice develop spontaneous autoimmune optic neuritis. J. Exp. Med. 197, 1073–1081 (2003).
Kadowaki, A. et al. Gut environment-induced intraepithelial autoreactive CD4( + ) T cells suppress central nervous system autoimmunity via LAG-3. Nat. Commun. 7, 11639 (2016).
Zudaire, E. et al. The aryl hydrocarbon receptor repressor is a putative tumor suppressor gene in multiple human cancers. J. Clin. Invest. 118, 640–650 (2008).
Haarmann-Stemmann, T. & Abel, J. The arylhydrocarbon receptor repressor (AhRR): structure, expression, and function. Biol. Chem. 387, 1195–1199 (2006).
Brandstatter, O. et al. Balancing intestinal and systemic inflammation through cell type-specific expression of the aryl hydrocarbon receptor repressor. Sci. Rep. 6, 26091 (2016).
Li, S., Bostick, J. W. & Zhou, L. Regulation of innate lymphoid cells by aryl hydrocarbon receptor. Front. Immunol. 8, 1909 (2017).
Melo-Gonzalez, F. & Hepworth, M. R. Functional and phenotypic heterogeneity of group 3 innate lymphoid cells. Immunology 150, 265–275 (2017).
Lee, J. S. et al. AHR drives the development of gut ILC22 cells and postnatal lymphoid tissues via pathways dependent on and independent of Notch. Nat. Immunol. 13, 144–151 (2011).
Kiss, E. A. et al. Natural aryl hydrocarbon receptor ligands control organogenesis of intestinal lymphoid follicles. Science 334, 1561–1565 (2011).
Qiu, J. et al. The aryl hydrocarbon receptor regulates gut immunity through modulation of innate lymphoid cells. Immunity 36, 92–104 (2012).
Qiu, J. et al. Group 3 innate lymphoid cells inhibit T cell-mediated intestinal inflammation through aryl hydrocarbon receptor signaling and regulation of microflora. Immunity 39, 386–399 (2013).
Wagage, S. et al. The group 3 innate lymphoid cell defect in aryl hydrocarbon receptor deficient mice is associated with T cell hyperactivation during intestinal infection. PLOS ONE 10, e0128335 (2015).
Li, J., Doty, A. & Glover, S. C. Aryl hydrocarbon receptor signaling involves in the human intestinal ILC3/ILC1 conversion in the inflamed terminal ileum of Crohn’s disease patients. Inflamm. Cell Signal. 3, e1404 (2016).
Wheeler, M. A. & Quintana, F. J. Regulation of astrocyte functions in multiple sclerosis. Cold Spring Harb. Perspect. Med. 9, a029009 (2018).
Khakh, B. S. & Sofroniew, M. V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 18, 942–952 (2015).
Pekny, M. et al. Astrocytes: a central element in neurological diseases. Acta Neuropathol. 131, 323–345 (2016).
Sofroniew, M. V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 16, 249–263 (2015). This is a comprehensive review of astrocyte functions in the inflamed CNS.
Rothhammer, V. & Quintana, F. J. Control of autoimmune CNS inflammation by astrocytes. Semin. Immunopathol. 37, 625–638 (2015).
Rothhammer, V. & Quintana, F. J. Environmental control of autoimmune inflammation in the central nervous system. Curr. Opin. Immunol. 43, 46–53 (2016).
Colonna, M. & Butovsky, O. Microglia function in the central nervous system during health and neurodegeneration. Annu. Rev. Immunol. 35, 441–468 (2017).
Kierdorf, K. & Prinz, M. Microglia in steady state. J. Clin. Invest. 127, 3201–3209 (2017).
Liddelow, S. A. et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 541, 481–487 (2017). This study describes mechanisms mediating microglial control of neurotoxic astrocytes.
Prinz, M. & Priller, J. Microglia and brain macrophages in the molecular age: from origin to neuropsychiatric disease. Nat. Rev. Neurosci. 15, 300–312 (2014).
Nannelli, A. et al. Effect of beta-naphthoflavone on AhR-regulated genes (CYP1A1, 1A2, 1B1, 2S1, Nrf2, and GST) and antioxidant enzymes in various brain regions of pig. Toxicology 265, 69–79 (2009).
Collins, L. L. et al. 2,3,7,8-Tetracholorodibenzo-p-dioxin exposure disrupts granule neuron precursor maturation in the developing mouse cerebellum. Toxicol. Sci. 103, 125–136 (2008).
Dever, D. P. et al. Aryl hydrocarbon receptor deletion in cerebellar granule neuron precursors impairs neurogenesis. Dev. Neurobiol. 76, 533–550 (2016).
Juricek, L. et al. AhR-deficiency as a cause of demyelinating disease and inflammation. Sci. Rep. 7, 9794 (2017).
Kim, S. Y. et al. Deletion of aryl hydrocarbon receptor AHR in mice leads to subretinal accumulation of microglia and RPE atrophy. Invest. Ophthalmol. Vis. Sci. 55, 6031–6040 (2014).
Xu, K., Yang, Z., Shi, R., Luo, C. & Zhang, Z. Expression of aryl hydrocarbon receptor in rat brain lesions following traumatic brain injury. Diagn. Pathol. 11, 72 (2016).
Buttgereit, A. et al. Sall1 is a transcriptional regulator defining microglia identity and function. Nat. Immunol. 17, 1397–1406 (2016).
Ayata, P. et al. Epigenetic regulation of brain region-specific microglia clearance activity. Nat. Neurosci. 21, 1049–1060 (2018).
Lee, Y. H. et al. Aryl hydrocarbon receptor mediates both proinflammatory and anti-inflammatory effects in lipopolysaccharide-activated microglia. Glia 63, 1138–1154 (2015).
Erny, D. et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 18, 965–977 (2015).
Sampson, T. R. et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell 167, 1469–1480 (2016). References 177 and 178 describe the control of microglia by microbial metabolites.
Erny, D. & Prinz, M. Microbiology: Gut microbes augment neurodegeneration. Nature 544, 304–305 (2017).
Rothhammer, V. et al. Dynamic regulation of serum aryl hydrocarbon receptor agonists in MS. Neurol. Neuroimmunol. Neuroinflamm. 4, e359 (2017).
Kaye, J. et al. Laquinimod arrests experimental autoimmune encephalomyelitis by activating the aryl hydrocarbon receptor. Proc. Natl Acad. Sci. USA 113, E6145–E6152 (2016).
Berg, J. et al. The immunomodulatory effect of laquinimod in CNS autoimmunity is mediated by the aryl hydrocarbon receptor. J. Neuroimmunol. 298, 9–15 (2016).
Vollmer, T. L. et al. A randomized placebo-controlled phase III trial of oral laquinimod for multiple sclerosis. J. Neurol. 261, 773–783 (2014).
Bruck, W. et al. Reduced astrocytic NF-kappaB activation by laquinimod protects from cuprizone-induced demyelination. Acta Neuropathol. 124, 411–424 (2012).
Lutterotti, A. et al. Antigen-specific tolerance by autologous myelin peptide-coupled cells: a phase 1 trial in multiple sclerosis. Sci. Transl Med. 5, 188ra175 (2013).
Weiner, H. L., da Cunha, A. P., Quintana, F. & Wu, H. Oral tolerance. Immunol. Rev. 241, 241–259 (2011).
Quintana, F. J., Carmi, P., Mor, F. & Cohen, I. R. Inhibition of adjuvant arthritis by a DNA vaccine encoding human heat shock protein 60. J. Immunol. 169, 3422–3428 (2002).
Robinson, W. H. et al. Protein microarrays guide tolerizing DNA vaccine treatment of autoimmune encephalomyelitis. Nat. Biotechnol. 21, 1033–1039 (2003).
Quintana, F. J. Nanoparticles for the induction of antigen-specific Tregs. Immunotherapy 5, 437–440 (2013).
Yeste, A., Nadeau, M., Burns, E. J., Weiner, H. L. & Quintana, F. J. Nanoparticle-mediated codelivery of myelin antigen and a tolerogenic small molecule suppresses experimental autoimmune encephalomyelitis. Proc. Natl Acad. Sci. USA 109, 11270–11275 (2012).
Miani, M. et al. Gut microbiota-stimulated innate lymphoid cells support beta-defensin 14 expression in pancreatic endocrine cells, preventing autoimmune diabetes. Cell Metab. 28, 557–572 (2018).
Smith, S. H. et al. Tapinarof is a natural AhR agonist that resolves skin inflammation in mice and humans. J. Invest. Dermatol. 137, 2110–2119 (2017).
Balachandran, V. P. et al. Imatinib potentiates antitumor T cell responses in gastrointestinal stromal tumor through the inhibition of Ido. Nat. Med. 17, 1094–1100 (2011).
van Baren, N. & Van den Eynde, B. J. Tumoral immune resistance mediated by enzymes that degrade tryptophan. Cancer Immunol. Res. 3, 978–985 (2015).
Pei, Z. et al. Aminoisoxazoles as potent inhibitors of tryptophan 2,3-dioxygenase 2 (TDO2). ACS Med. Chem. Lett. 9, 417–421 (2018).
Laurans, L. et al. Genetic deficiency of indoleamine 2,3-dioxygenase promotes gut microbiota-mediated metabolic health. Nat. Med. 24, 1113–1120 (2018).
Gabriely, G., Wheeler, M. A., Takenaka, M. C. & Quintana, F. J. Role of AHR and HIF-1alpha in glioblastoma metabolism. Trends Endocrinol. Metab. 28, 428–436 (2017).
Triplett, T. A. et al. Reversal of indoleamine 2,3-dioxygenase-mediated cancer immune suppression by systemic kynurenine depletion with a therapeutic enzyme. Nat. Biotechnol. 36, 758–764 (2018).
Opitz, C. A. et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 478, 197–203 (2011). This study reports a novel role for tumour tryptophan metabolites in the pathogenesis of glioblastoma.
Gramatzki, D. et al. Aryl hydrocarbon receptor inhibition downregulates the TGF-beta/Smad pathway in human glioblastoma cells. Oncogene 28, 2593–2605 (2009).
Silginer, M. et al. The aryl hydrocarbon receptor links integrin signaling to the TGF-beta pathway. Oncogene 35, 3260–3271 (2016).
Takanaga, H., Yoshitake, T., Yatabe, E., Hara, S. & Kunimoto, M. Beta-naphthoflavone disturbs astrocytic differentiation of C6 glioma cells by inhibiting autocrine interleukin-6. J. Neurochem. 90, 750–757 (2004).
Litzenburger, U. M. et al. Constitutive IDO expression in human cancer is sustained by an autocrine signaling loop involving IL-6, STAT3 and the AHR. Oncotarget 5, 1038–1051 (2014).
Nguyen, L. P. & Bradfield, C. A. The search for endogenous activators of the aryl hydrocarbon receptor. Chem. Res. Toxicol. 21, 102–116 (2008).
Xu, S. et al. Synergy between the ectoenzymes CD39 and CD73 contributes to adenosinergic immunosuppression in human malignant gliomas. Neuro Oncol. 15, 1160–1172 (2013).
Acknowledgements
This work was supported by grants NS102807, NS087867, ES02530, AI126880 and AI093903 from the US National Institutes of Health, RSG-14-198-01-LIB from the American Cancer Society, RG4111A1 and JF2161-A-5 from the National Multiple Sclerosis Society and PA-1604-08459 from the International Progressive MS Alliance. The authors thank all members of the Quintana laboratory for helpful advice and discussions.
Author contributions
Both V.R. and F.J.Q. researched data and reviewed and edited the manuscript. V.R. wrote the manuscript.
Reviewer information
Nature Reviews Immunology thanks M. Colonna and other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
F.J.Q. is a founder of AnTolRx, a company exploring the targeting of aryl hydrocarbon receptor (AHR) with nanoparticles for the treatment of inflammatory conditions. He is also a scientific adviser for Kyn Therapeutics, a company exploring targeting AHR for cancer therapy.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Glossary
- Mediator complex
-
An evolutionarily conserved multisubunit protein complex controlling the activities of RNA polymerase II, which transcribes all protein-coding genes in eukaryotes. Mediator serves as a functional bridge between specific transcription factors, DNA-bound transcriptional activators or repressors and the transcriptional machinery required for the initiation of transcription by the basal transcriptional machinery (including RNA polymerase II and general transcription factors).
- SWI/SNF chromatin remodelling complex
-
(Switching-defective/sucrose non-fermenting chromatin remodelling complex). An ATP-dependent chromatin remodelling protein complex that was first identified in yeast. Related complexes exist in mammals (where they are known as BAF) and are involved in the chromatin remodelling of various genes.
- Kynurenine pathway
-
A metabolic pathway in which the essential amino acid tryptophan is metabolized to the cofactor NAD+ over several enzymatic steps. The first reaction in this process, mediated by the enzymes indoleamine 2,3-dioxygenase 1 (IDO1), IDO2 or tryptophan 2,3-dioxygenase (TDO), leads to the generation of kynurenine. Kynurenine and some of its metabolites act as aryl hydrocarbon receptor (AHR) ligands and exert important functions in the immune system in inflammatory, infectious and neoplastic disorders.
- T regulatory type 1 cells
-
(TR1 cells). A population of regulatory T cells that arises in the periphery after an encounter with antigen in the presence of IL-27 alone or in combination with transforming growth factor-β (TGFβ) and that regulates immune responses through the secretion of IL-10 and additional mechanisms. TR1 cells suppress T cell responses, downregulate the expression of co-stimulatory molecules and pro-inflammatory cytokines by antigen-presenting cells and favour the production of IgD, IgA and IgG by B cells.
- Intraepithelial lymphocytes
-
(IELs). A T cell population found within the epithelial layer of mammalian mucosal linings. This population consists of specialized subsets of cells, such as particular T cell receptor (TCR) γδ+ T cell subsets and TCRαβ+CD8αα+ T cells.
- Innate lymphoid cells
-
(ILCs). A population of innate immune cells that are lymphoid in morphology and developmental origin but lack properties of adaptive B cells and T cells such as recombined antigen-specific receptors. They function in the regulation of immunity, tissue homeostasis and inflammation in response to cytokine stimulation.
Rights and permissions
About this article

Cite this article
Rothhammer, V., Quintana, F.J. The aryl hydrocarbon receptor: an environmental sensor integrating immune responses in health and disease. Nat Rev Immunol 19, 184–197 (2019). https://doi.org/10.1038/s41577-019-0125-8
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41577-019-0125-8
This article is cited by
-
Bisphenol a downregulates GLUT4 expression by activating aryl hydrocarbon receptor to exacerbate polycystic ovary syndrome
Cell Communication and Signaling (2024)
-
Bioluminescence imaging of Cyp1a1-luciferase reporter mice demonstrates prolonged activation of the aryl hydrocarbon receptor in the lung
Communications Biology (2024)
-
Myeloid cell-associated aromatic amino acid metabolism facilitates CNS myelin regeneration
npj Regenerative Medicine (2024)
-
Aryl hydrocarbon receptor suppresses STING-mediated type I IFN expression in triple-negative breast cancer
Scientific Reports (2024)
-
Gastrointestinal and brain barriers: unlocking gates of communication across the microbiota–gut–brain axis
Nature Reviews Gastroenterology & Hepatology (2024)
