
Abstract
Perturbations in metabolic processes are associated with diseases such as obesity, type 2 diabetes mellitus, certain infections and some cancers. A resurgence of interest in creatine biology is developing, with new insights into a diverse set of regulatory functions for creatine. This resurgence is primarily driven by technological advances in genetic engineering and metabolism as well as by the realization that this metabolite has key roles in cells beyond the muscle and brain. Herein, we highlight the latest advances in creatine biology in tissues and cell types that have historically received little attention in the field. In adipose tissue, creatine controls thermogenic respiration and loss of this metabolite impairs whole-body energy expenditure, leading to obesity. We also cover the various roles that creatine metabolism has in cancer cell survival and the function of the immune system. Renewed interest in this area has begun to showcase the therapeutic potential that lies in understanding how changes in creatine metabolism lead to metabolic disease.
Key points
-
Mitochondria in brown adipose tissue are capable of normal oxidative phosphorylation, with P:O ratios similar to those of other tissues.
-
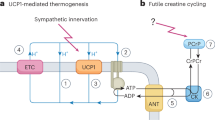
Atypical actions of creatine involve phosphocreatine transport into colorectal cancer cells, super-stoichiometric ADP liberation to trigger respiration in thermogenic adipocytes and chromatin remodelling to modulate macrophage polarity.
-
Cyclocreatine and creatine can both inhibit tumour progression, suggesting that the pro-cancer role of creatine is independent of its function in energy buffering.
-
The mitochondrial network transduces energy over long distances, thus minimizing the requirement for metabolite diffusion, whereas cells with a disrupted mitochondrial network might buffer energy via the creatine kinase–phosphocreatine circuit.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


References
da Silva, R. P., Clow, K., Brosnan, J. T. & Brosnan, M. E. Synthesis of guanidinoacetate and creatine from amino acids by rat pancreas. Br. J. Nutr. 111, 571–577 (2014).
Kazak, L. et al. Genetic depletion of adipocyte creatine metabolism inhibits diet-induced thermogenesis and drives obesity. Cell Metab. 26, 660–671 (2017).
Russell, A. P. et al. Creatine transporter (SLC6A8) knockout mice display an increased capacity for in vitro creatine biosynthesis in skeletal muscle. Front. Physiol. 5, 314 (2014).
Haas, R. C. & Strauss, A. W. Separate nuclear genes encode sarcomere-specific and ubiquitous human mitochondrial creatine kinase isoenzymes. J. Biol. Chem. 265, 6921–6927 (1990).
Hossle, J. P. et al. Distinct tissue specific mitochondrial creatine kinases from chicken brain and striated muscle with a conserved CK framework. Biochem. Biophys. Res. Commun. 151, 408–416 (1988).
Wallimann, T., Wyss, M., Brdiczka, D., Nicolay, K. & Eppenberger, H. M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: the ‘phosphocreatine circuit’ for cellular energy homeostasis. Biochem. J. 281, 21–40 (1992).
Li, H. et al. Regulation of the creatine transporter by AMP-activated protein kinase in kidney epithelial cells. Am. J. Physiol. Ren. Physiol. 299, F167–F177 (2010).
Bessman, S. P. & Geiger, P. J. Transport of energy in muscle: the phosphorylcreatine shuttle. Science 211, 448–452 (1981).
Glancy, B. et al. Mitochondrial reticulum for cellular energy distribution in muscle. Nature 523, 617–620 (2015).
Meyer, R. A., Sweeney, H. L. & Kushmerick, M. J. A simple analysis of the ‘‘phosphocreatine shuttle’’. Am. J. Physiol. 246, C365–C377 (1984).
Kenyon, G. L. & Reed, G. H. Creatine kinase: structure-activity relationships. Adv. Enzymol. Relat. Areas Mol. Biol. 54, 367–426 (1983).
Schlattner, U., Tokarska-Schlattner, M. & Wallimann, T. Mitochondrial creatine kinase in human health and disease. Biochim. Biophys. Acta 1762, 164–180 (2006).
Bessman, S. P. The physiological significance of the creatine phosphate shuttle. Adv. Exp. Med. Biol. 194, 1–11 (1986).
Riesberg, L. A., Weed, S. A., McDonald, T. L., Eckerson, J. M. & Drescher, K. M. Beyond muscles: the untapped potential of creatine. Int. Immunopharmacol. 37, 31–42 (2016).
Wyss, M. & Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 80, 1107–1213 (2000).
Dzeja, P. P. & Terzic, A. Phosphotransfer networks and cellular energetics. J. Expt. Biol. 206, 2039–2047 (2003).
Schlattner, U. et al. Cellular compartmentation of energy metabolism: creatine kinase microcompartments and recruitment of B-type creatine kinase to specific subcellular sites. Amino Acids 48, 1751–1774 (2016).
Braun, K., Oeckl, J., Westermeier, J., Li, Y. & Klingenspor, M. Non-adrenergic control of lipolysis and thermogenesis in adipose tissues. J. Exp. Biol. 221 (Suppl. 1), jeb165381 (2018).
Rosen, E. D. & Spiegelman, B. M. What we talk about when we talk about fat. Cell 156, 20–44 (2014).
Foster, D. O. & Frydman, M. L. Brown adipose tissue: the dominant site of nonshivering thermogenesis in the rat. Experientia Suppl. 32, 147–151 (1978).
Rothwell, N. J. & Stock, M. J. A role for brown adipose tissue in diet-induced thermogenesis. Nature 281, 31–35 (1979).
Chouchani, E. T., Kazak, L. & Spiegelman, B. M. New advances in adaptive thermogenesis: UCP1 and beyond. Cell Metab. 29, 27–37 (2019).
Lowell, B. B. et al. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 366, 740–742 (1993).
Cohen, P. et al. Ablation of PRDM16 and beige adipose causes metabolic dysfunction and a subcutaneous to visceral fat switch. Cell 156, 304–316 (2014).
van Marken Lichtenbelt, W. D. et al. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 360, 1500–1508 (2009).
Carpentier, A. C. et al. Brown adipose tissue energy metabolism in humans. Front. Endocrinol. 9, 447 (2018).
Leitner, B. P. et al. Mapping of human brown adipose tissue in lean and obese young men. Proc. Natl Acad. Sci. USA 114, 8649–8654 (2017).
Golozoubova, V. et al. Only UCP1 can mediate adaptive nonshivering thermogenesis in the cold. FASEB J. 15, 2048–2050 (2001).
Meyer, C. W. et al. Adaptive thermogenesis and thermal conductance in wild-type and UCP1-KO mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 299, R1396–R1406 (2010).
Ukropec, J., Anunciado, R. P., Ravussin, Y., Hulver, M. W. & Kozak, L. P. UCP1-independent thermogenesis in white adipose tissue of cold-acclimated Ucp1 −/− mice. J. Biol. Chem. 281, 31894–31908 (2006).
Keipert, S. et al. Long-term cold adaptation does not require FGF21 or UCP1. Cell Metab. 26, 437–446 (2017).
Golozoubova, V., Cannon, B. & Nedergaard, J. UCP1 is essential for adaptive adrenergic nonshivering thermogenesis. Am. J. Physiol. Endocrinol. Metab. 291, E350–E357 (2006).
Hofmann, W. E., Liu, X., Bearden, C. M., Harper, M. E. & Kozak, L. P. Effects of genetic background on thermoregulation and fatty acid-induced uncoupling of mitochondria in UCP1-deficient mice. J. Biol. Chem. 276, 12460–12465 (2001).
Kazak, L. et al. UCP1 deficiency causes brown fat respiratory chain depletion and sensitizes mitochondria to calcium overload-induced dysfunction. Proc. Natl Acad. Sci. USA 114, 7981–7986 (2017).
Cannon, B. & Nedergaard, J. Nonshivering thermogenesis and its adequate measurement in metabolic studies. J. Exp. Biol. 214, 242–253 (2011).
Matthias, A. et al. Thermogenic responses in brown fat cells are fully UCP1-dependent. UCP2 or UCP3 do not substitute for UCP1 in adrenergically or fatty scid-induced thermogenesis. J. Biol. Chem. 275, 25073–25081 (2000).
Oelkrug, R., Kutschke, M., Meyer, C. W., Heldmaier, G. & Jastroch, M. Uncoupling protein 1 decreases superoxide production in brown adipose tissue mitochondria. J. Biol. Chem. 285, 21961–21968 (2010).
Enerback, S. et al. Mice lacking mitochondrial uncoupling protein are cold-sensitive but not obese. Nature 387, 90–94 (1997).
Rothwell, N. J. & Stock, M. J. A role for brown adipose tissue in diet-induced thermogenesis. Obes. Res. 5, 650–656 (1997).
Feldmann, H. M., Golozoubova, V., Cannon, B. & Nedergaard, J. UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metab. 9, 203–209 (2009).
Maurer, S. F., Fromme, T., Mocek, S., Zimmermann, A. & Klingenspor, M. Uncoupling protein 1 and the capacity for non-shivering thermogenesis are components of the glucose homeostatic system. Am. J. Physiol. Endocrinol. Metab. 318, E198–E215 (2020).
Fromme, T. et al. Bile acid supplementation decreases body mass gain in C57BL/6J but not 129S6/SvEvTac mice without increasing energy expenditure. Sci. Rep. 9, 131 (2019).
Liu, X. et al. Paradoxical resistance to diet-induced obesity in UCP1-deficient mice. J. Clin. Invest. 111, 399–407 (2003).
Zietak, M. & Kozak, L. P. Bile acids induce uncoupling protein 1-dependent thermogenesis and stimulate energy expenditure at thermoneutrality in mice. Am. J. Physiol. Endocrinol. Metab. 310, E346–E354 (2016).
Bachman, E. S. et al. betaAR signaling required for diet-induced thermogenesis and obesity resistance. Science 297, 843–845 (2002).
Shi, F. & Collins, S. Second messenger signaling mechanisms of the brown adipocyte thermogenic program: an integrative perspective. Horm. Mol. Biol. Clin. Investig. https://doi.org/10.1515/hmbci-2017-0062 (2017).
Hamann, A., Flier, J. S. & Lowell, B. B. Decreased brown fat markedly enhances susceptibility to diet-induced obesity, diabetes, and hyperlipidemia. Endocrinology 137, 21–29 (1996).
Douris, N. et al. Beta-adrenergic receptors are critical for weight loss but not for other metabolic adaptations to the consumption of a ketogenic diet in male mice. Mol. Metab. 6, 854–862 (2017).
Anunciado-Koza, R., Ukropec, J., Koza, R. A. & Kozak, L. P. Inactivation of UCP1 and the glycerol phosphate cycle synergistically increases energy expenditure to resist diet-induced obesity. J. Biol. Chem. 283, 27688–27697 (2008).
Granneman, J. G., Burnazi, M., Zhu, Z. & Schwamb, L. A. White adipose tissue contributes to UCP1-independent thermogenesis. Am. J. Physiol. Endocrinol. Metab. 285, E1230–E1236 (2003).
Ukropec, J., Anunciado, R. V., Ravussin, Y. & Kozak, L. P. Leptin is required for uncoupling protein-1-independent thermogenesis during cold stress. Endocrinology 147, 2468–2480 (2006).
Shabalina, I. G. et al. UCP1 in brite/beige adipose tissue mitochondria is functionally thermogenic. Cell Rep. 5, 1196–1203 (2013).
Kazak, L. et al. A creatine-driven substrate cycle enhances energy expenditure and thermogenesis in beige fat. Cell 163, 643–655 (2015).
Antonacci, M. A. et al. Direct detection of brown adipose tissue thermogenesis in UCP1−/− mice by hyperpolarized (129)Xe MR thermometry. Sci. Rep. 9, 14865 (2019).
Kontani, Y. et al. UCP1 deficiency increases susceptibility to diet-induced obesity with age. Aging Cell 4, 147–155 (2005).
Pfannenberg, C. et al. Impact of age on the relationships of brown adipose tissue with sex and adiposity in humans. Diabetes 59, 1789–1793 (2010).
Cypess, A. M. et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 360, 1509–1517 (2009).
Harms, M. J. et al. Prdm16 is required for the maintenance of brown adipocyte identity and function in adult mice. Cell Metab. 19, 593–604 (2014).
Muller, S. et al. Proteomic analysis of human brown adipose tissue reveals utilization of coupled and uncoupled energy expenditure pathways. Sci. Rep. 6, 30030 (2016).
Bertholet, A. M. et al. Mitochondrial patch clamp of beige adipocytes reveals UCP1-positive and UCP1-negative cells both exhibiting futile creatine cycling. Cell Metab. 25, 811–822 (2017).
Kazak, L. et al. Ablation of adipocyte creatine transport impairs thermogenesis and causes diet-induced obesity. Nat. Metab. 1, 360–370 (2019).
Yamashita, H. et al. Increased growth of brown adipose tissue but its reduced thermogenic activity in creatine-depleted rats fed β-guanidinopropionic acid. Biochim. Biophys. Acta 1230, 69–73 (1995).
Wakatsuki, T. et al. Thermogenic responses to high-energy phosphate contents and/or hindlimb suspension in rats. Jpn. J. Physiol. 46, 171–175 (1996).
Svensson, P. A. et al. Gene expression in human brown adipose tissue. Int. J. Mol. Med. 27, 227–232 (2011).
Wada, S. et al. The tumor suppressor FLCN mediates an alternate mTOR pathway to regulate browning of adipose tissue. Genes Dev. 30, 2551–2564 (2016).
Perna, M. K. et al. Creatine transporter deficiency leads to increased whole body and cellular metabolism. Amino Acids 48, 2057–2065 (2016).
Schulz, T. J. et al. Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 495, 379–383 (2013).
Rowland, L. A., Bal, N. C., Kozak, L. P. & Periasamy, M. Uncoupling protein 1 and sarcolipin are required to maintain optimal thermogenesis, and loss of both systems compromises survival of mice under cold stress. J. Biol. Chem. 290, 12282–12289 (2015).
Berlet, H. H., Bonsmann, I. & Birringer, H. Occurrence of free creatine, phosphocreatine and creatine phosphokinase in adipose tissue. Biochim. Biophys. Acta 437, 166–174 (1976).
Din, M. U. et al. Postprandial oxidative metabolism of human brown fat indicates thermogenesis. Cell Metab. 28, 207–216.e3 (2018).
Jash, S., Banerjee, S., Lee, M. J., Farmer, S. R. & Puri, V. CIDEA transcriptionally regulates UCP1 for britening and thermogenesis in human fat cells. iScience 20, 73–89 (2019).
Pydi, S. P. et al. Adipocyte β-arrestin-2 is essential for maintaining whole body glucose and energy homeostasis. Nat. Commun. 10, 2936 (2019).
Wang, L. et al. Selective activation of Gs signaling in adipocytes causes striking metabolic improvements in mice. Mol. Metab. 27, 83–91 (2019).
Pierce, K. L., Premont, R. T. & Lefkowitz, R. J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 3, 639–650 (2002).
Lee, C. C. et al. Naa10p inhibits beige adipocyte-mediated thermogenesis through N-α-acetylation of Pgc1alpha. Mol. Cell 76, 500–515.e8 (2019).
Deshmukh, A. S. et al. Proteomics-based comparative mapping of the secretomes of human brown and white adipocytes reveals EPDR1 as a novel batokine. Cell Metab. 30, 963–975 (2019).
Levy, S. E., Chen, Y. S., Graham, B. H. & Wallace, D. C. Expression and sequence analysis of the mouse adenine nucleotide translocase 1 and 2 genes. Gene 254, 57–66 (2000).
Jacobus, W. E. & Lehninger, A. L. Creatine kinase of rat heart mitochondria. Coupling of creatine phosphorylation to electron transport. J. Biol. Chem. 248, 4803–4810 (1973).
Seo, J. B. et al. Knockdown of Ant2 reduces adipocyte Hypoxia and improves insulin resistance in obesity. Nat. Metab. 1, 86–97 (2019).
Brand, M. D. et al. The basal proton conductance of mitochondria depends on adenine nucleotide translocase content. Biochem. J. 392, 353–362 (2005).
Bertholet, A. M. et al. H+ transport is an integral function of the mitochondrial ADP/ATP carrier. Nature 571, 515–520 (2019).
Streijger, F. et al. Mice lacking brain-type creatine kinase activity show defective thermoregulation. Physiol. Behav. 97, 76–86 (2009).
Streijger, F. et al. Structural and behavioural consequences of double deficiency for creatine kinases BCK and UbCKmit. Behav. Brain Res. 157, 219–234 (2005).
Virtue, S. & Vidal-Puig, A. Assessment of brown adipose tissue function. Front. Physiol. 4, 128 (2013).
Wikstrom, J. D. et al. Hormone-induced mitochondrial fission is utilized by brown adipocytes as an amplification pathway for energy expenditure. EMBO J. 33, 418–436 (2014).
Friedman, J. R. et al. ER tubules mark sites of mitochondrial division. Science 334, 358–362 (2011).
Zotter, A., Bauerle, F., Dey, D., Kiss, V. & Schreiber, G. Quantifying enzyme activity in living cells. J. Biol. Chem. 292, 15838–15848 (2017).
Schlattner, U. et al. Divergent enzyme kinetics and structural properties of the two human mitochondrial creatine kinase isoenzymes. Biol. Chem. 381, 1063–1070 (2000).
Menniti, F. S., Knoth, J., Peterson, D. S. & Diliberto, E. J. Jr. The in situ kinetics of dopamine beta-hydroxylase in bovine adrenomedullary chromaffin cells. Intravesicular compartmentation reduces apparent affinity for the cofactor ascorbate. J. Biol. Chem. 262, 7651–7657 (1987).
Nakae, Y. & Stoward, P. J. Kinetic parameters of lactate dehydrogenase in liver and gastrocnemius determined by three quantitative histochemical methods. J. Histochem. Cytochem. 45, 1427–1431 (1997).
Joel, C. D., Neaves, W. B. & Rabb, J. M. Mitochondria of brown fat: oxidative phosphorylation sensitive to 2,4,-dinitrophenol. Biochem. Biophys. Res. Commun. 29, 490–495 (1967).
Guillory, R. J. & Racker, E. Oxidative phosphorylation in brown adipose mitochondria. Biochim. Biophys. Acta 153, 490–493 (1968).
Hittelman, K. J., Lindberg, O. & Cannon, B. Oxidative phosphorylation and compartmentation of fatty acid metabolism in brown fat mitochondria. Eur. J. Bio. 11, 183–192 (1969).
Williamson, J. R. Control of energy metabolism in hamster brown adipose tissue. J. Biol. Chem. 245, 2043–2050 (1970).
Prusiner, S. B., Cannon, B., Ching, T. M. & Lindberg, O. Oxidative metabolism in cells isolated from brown adipose tissue. 2. Catecholamine regulated respiratory control. Eur. J. Biochem. 7, 51–57 (1968).
LaNoue, K. F., Koch, C. D. & Meditz, R. B. Mechanism of action of norepinephrine in hamster brown adipocytes. J. Biol. Chem. 257, 13740–13748 (1982).
Nedergaard, J. & Lindberg, O. Norepinephrine-stimulated fatty-acid release and oxygen consumption in isolated hamster brown-fat cells. Influence of buffers, albumin, insulin and mitochondrial inhibitors. Eur. J. Biochem. 95, 139–145 (1979).
Mills, E. L. et al. Accumulation of succinate controls activation of adipose tissue thermogenesis. Nature 560, 102–106 (2018).
Yoneshiro, T. et al. BCAA catabolism in brown fat controls energy homeostasis through SLC25A44. Nature 572, 614–619 (2019).
Schreiber, R. et al. Cold-induced thermogenesis depends on ATGL-mediated lipolysis in cardiac muscle, but not brown adipose tissue. Cell Metab. 26, 753–763 (2017).
Shin, H. et al. Lipolysis in brown adipocytes is not essential for cold-induced thermogenesis in mice. Cell Metab. 26, 764–777 (2017).
Bal, N. C. et al. Sarcolipin is a newly identified regulator of muscle-based thermogenesis in mammals. Nat. Med. 18, 1575–1579 (2012).
MacPherson, R. E. et al. Sarcolipin knockout mice fed a high-fat diet exhibit altered indices of adipose tissue inflammation and remodeling. Obesity 24, 1499–1505 (2016).
Song, A. et al. Low- and high-thermogenic brown adipocyte subpopulations coexist in murine adipose tissue. J. Clin. Invest. 130, 247–257 (2020).
Goldrath, A. W. & Bevan, M. J. Selecting and maintaining a diverse T-cell repertoire. Nature 402, 255–262 (1999).
Davis, M. M. & Bjorkman, P. J. T-cell antigen receptor genes and T-cell recognition. Nature 334, 395–402 (1988).
Hogquist, K. A., Baldwin, T. A. & Jameson, S. C. Central tolerance: learning self-control in the thymus. Nat. Rev. Immunol. 5, 772–782 (2005).
Zhang, Y., Li, H., Wang, X., Gao, X. & Liu, X. Regulation of T cell development and activation by creatine kinase B. PLoS One 4, e5000 (2009).
Davies, L. C., Jenkins, S. J., Allen, J. E. & Taylor, P. R. Tissue-resident macrophages. Nat. Immunol. 14, 986–995 (2013).
Gordon, S. & Martinez, F. O. Alternative activation of macrophages: mechanism and functions. Immunity 32, 593–604 (2010).
DeChatelet, L. R., McCall, C. E. & Shirley, P. S. Creatine phosphokinase activity in rabbit alveolar macrophages. Infect. Immun. 7, 29–34 (1973).
Loike, J. D., Kozler, V. F. & Silverstein, S. C. Increased ATP and creatine phosphate turnover in phagocytosing mouse peritoneal macrophages. J. Biol. Chem. 254, 9558–9564 (1979).
Loike, J. D., Kozler, V. F. & Silverstein, S. C. Creatine kinase expression and creatine phosphate accumulation are developmentally regulated during differentiation of mouse and human monocytes. J. Exp. Med. 159, 746–757 (1984).
Ji, L. et al. Slc6a8-mediated creatine uptake and accumulation reprogram macrophage polarization via regulating cytokine responses. Immunity 51, 272–284 (2019).
Guminska, M., Ptak, W. & Zembala, M. Macrophage metabolism during phagocytosis and digestion of normal and IgG antibody-coated sheep erythrocytes. Enzyme 19, 24–37 (1975).
Kuiper, J. W. et al. Creatine kinase-mediated ATP supply fuels actin-based events in phagocytosis. PLoS Biol. 6, e51 (2008).
Kelly, P. N. The cancer immunotherapy revolution. Science 359, 1344–1345 (2018).
Di Biase, S. et al. Creatine uptake regulates CD8 T cell antitumor immunity. J. Exp. Med. 216, 2869–2882 (2019).
Turer, E. et al. Creatine maintains intestinal homeostasis and protects against colitis. Proc. Natl. Acad. Sci. USA 114, E1273–E1281 (2017).
Gupta, G. P. & Massague, J. Cancer metastasis: building a framework. Cell 127, 679–695 (2006).
Vander Heiden, M. G. & DeBerardinis, R. J. Understanding the intersections between metabolism and cancer biology. Cell 168, 657–669 (2017).
Dupuy, F. et al. PDK1-dependent metabolic reprogramming dictates metastatic potential in breast cancer. Cell Metab. 22, 577–589 (2015).
Fenouille, N. et al. The creatine kinase pathway is a metabolic vulnerability in EVI1-positive acute myeloid leukemia. Nat. Med. 23, 301–313 (2017).
Kurmi, K. et al. Tyrosine phosphorylation of mitochondrial creatine kinase 1 enhances a druggable tumor energy shuttle pathway. Cell Metab. 28, 833–847 (2018).
Lillie, J. W. et al. Cyclocreatine (1-carboxymethyl-2-iminoimidazolidine) inhibits growth of a broad spectrum of cancer cells derived from solid tumors. Cancer Res. 53, 3172–3178 (1993).
Miller, E. E., Evans, A. E. & Cohn, M. Inhibition of rate of tumor growth by creatine and cyclocreatine. Proc. Natl Acad. Sci. USA 90, 3304–3308 (1993).
Annesley, T. M. & Walker, J. B. Cyclocreatine phosphate as a substitute for creatine phosphate in vertebrate tissues. Energistic considerations. Biochem. Biophys. Res. Commun. 74, 185–190 (1977).
Kristensen, C. A., Askenasy, N., Jain, R. K. & Koretsky, A. P. Creatine and cyclocreatine treatment of human colon adenocarcinoma xenografts: 31P and 1H magnetic resonance spectroscopic studies. Br. J. Cancer 79, 278–285 (1999).
Gaidzik, V. I. et al. RUNX1 mutations in acute myeloid leukemia: results from a comprehensive genetic and clinical analysis from the AML study group. J. Clin. Oncol. 29, 1364–1372 (2011).
Feld, R. D. & Witte, D. L. Presence of creatine kinase BB isoenzyme in some patients with prostatic carcinoma. Clin. Chem. 23, 1930–1932 (1977).
Gazdar, A. F. et al. Levels of creatine kinase and its BB isoenzyme in lung cancer specimens and cultures. Cancer Res. 41, 2773–2777 (1981).
Li, X. H. et al. Knockdown of creatine kinase B inhibits ovarian cancer progression by decreasing glycolysis. Int. J. Biochem. Cell Biol. 45, 979–986 (2013).
Glass, C., Wilson, M., Gonzalez, R., Zhang, Y. & Perkins, A. S. The role of EVI1 in myeloid malignancies. Blood Cells Mol. Dis. 53, 67–76 (2014).
Benajiba, L. et al. Creatine kinase pathway inhibition alters GSK3 and WNT signaling in EVI1-positive AML. Leukemia 33, 800–804 (2019).
Saks, V. A., Dzhaliashvili, I. V., Konorev, E. A. & Strumia, E. Molecular and cellular aspects of the cardioprotective mechanism of phosphocreatine. Biokhimiia 57, 1763–1784 (1992).
Orth, M. F. et al. Functional genomics identifies AMPD2 as a new prognostic marker for undifferentiated pleomorphic sarcoma. Int. J. Cancer 144, 859–867 (2019).
Wong, M. et al. AMPD3 is associated with the malignant characteristics of gastrointestinal stromal tumors. Oncol. Lett. 13, 1281–1287 (2017).
Tokarska-Schlattner, M. et al. Phosphocreatine interacts with phospholipids, affects membrane properties and exerts membrane-protective effects. PLoS One 7, e43178 (2012).
Zucchi, R. et al. Protection of isolated rat heart from oxidative stress by exogenous creatine phosphate. J. Mol. Cell. Cardiol. 21, 67–73 (1989).
Johannsen, S. et al. Screening test for malignant hyperthermia in patients with persistent hyperCKemia: a pilot study. Muscle Nerve 47, 677–681 (2013).
Loo, J. M. et al. Extracellular metabolic energetics can promote cancer progression. Cell 160, 393–406 (2015).
Ackerman, S. E., Blackburn, O. A., Marchildon, F. & Cohen, P. Insights into the link between obesity and cancer. Curr. Obes. Rep. 6, 195–203 (2017).
Olson, O. C., Quail, D. F. & Joyce, J. A. Obesity and the tumor microenvironment. Science 358, 1130–1131 (2017).
Park, J. & Scherer, P. E. Endotrophin - a novel factor linking obesity with aggressive tumor growth. Oncotarget 3, 1487–1488 (2012).
Khandekar, M. J., Cohen, P. & Spiegelman, B. M. Molecular mechanisms of cancer development in obesity. Nat. Rev. Cancer 11, 886–895 (2011).
Hamady, Z. Z. et al. Fatty liver disease as a predictor of local recurrence following resection of colorectal liver metastases. Br. J. Surg. 100, 820–826 (2013).
VanSaun, M. N., Lee, I. K., Washington, M. K., Matrisian, L. & Gorden, D. L. High fat diet induced hepatic steatosis establishes a permissive microenvironment for colorectal metastases and promotes primary dysplasia in a murine model. Am. J. Pathol. 175, 355–364 (2009).
Ohashi, K. et al. NOD-like receptor C4 inflammasome regulates the growth of colon cancer liver metastasis in NAFLD. Hepatology 70, 1582–1599 (2019).
Wu, Y. et al. Insulin-like growth factor-I regulates the liver microenvironment in obese mice and promotes liver metastasis. Cancer Res. 70, 57–67 (2010).
Smith, B. K. et al. Treatment of nonalcoholic fatty liver disease: role of AMPK. Am. J. Physiol. Endocrinol. Metab. 311, E730–E740 (2016).
Komatsu, M. et al. NNMT activation can contribute to the development of fatty liver disease by modulating the NAD+ metabolism. Sci Rep. 8, 8637 (2018).
Lowe, M. T., Kim, E. H., Faull, R. L., Christie, D. L. & Waldvogel, H. J. Dissociated expression of mitochondrial and cytosolic creatine kinases in the human brain: a new perspective on the role of creatine in brain energy metabolism. J. Cereb. Blood Flow Metab. 33, 1295–1306 (2013).
Kuiper, J. W. et al. Local ATP generation by brain-type creatine kinase (CK-B) facilitates cell motility. PLoS One 4, e5030 (2009).
Nevo, A. C. & Rikmenspoel, R. Diffusion of ATP in sperm flagella. J. Theor. Biol. 26, 11–18 (1970).
Tombes, R. M., Brokaw, C. J. & Shapiro, B. M. Creatine kinase-dependent energy transport in sea urchin spermatozoa. Flagellar wave attenuation and theoretical analysis of high energy phosphate diffusion. Biophys. J. 52, 75–86 (1987).
Papalazarou, V. et al. The creatine–phosphagen system is mechanoresponsive in pancreatic adenocarcinoma and fuels invasion and metastasis. Nat. Metab. 2, 62–80 (2020).
Ellington, W. R. Evolution and physiological roles of phosphagen systems. Annu. Rev. Physiol. 63, 289–325 (2001).
Schwarz, T. L. Mitochondrial trafficking in neurons. Cold Spring Harb. Perspect. Biol. 5, a011304 (2013).
Kaldis, P. et al. Identification of two distinctly localized mitochondrial creatine kinase isoenzymes in spermatozoa. J. Cell Sci. 109, 2079–2088 (1996).
Skulachev, V. P. Mitochondrial filaments and clusters as intracellular power-transmitting cables. Trends Biochem. Sci. 26, 23–29 (2001).
Steeghs, K. et al. Cytoarchitectural and metabolic adaptations in muscles with mitochondrial and cytosolic creatine kinase deficiencies. Mol. Cell. Biochem. 184, 183–194 (1998).
van Deursen, J. et al. Skeletal muscles of mice deficient in muscle creatine kinase lack burst activity. Cell 74, 621–631 (1993).
van Deursen, J. et al. Creatine kinase (CK) in skeletal muscle energy metabolism: a study of mouse mutants with graded reduction in muscle CK expression. Proc. Natl Acad. Sci. USA 91, 9091–9095 (1994).
Steeghs, K. et al. Use of gene targeting for compromising energy homeostasis in neuro-muscular tissues: the role of sarcomeric mitochondrial creatine kinase. J. Neurosci. Methods 71, 29–41 (1997).
Steeghs, K. et al. Mouse ubiquitous mitochondrial creatine kinase: gene organization and consequences from inactivation in mouse embryonic stem cells. DNA Cell Biol. 14, 539–553 (1995).
Ligon, L. A. & Steward, O. Role of microtubules and actin filaments in the movement of mitochondria in the axons and dendrites of cultured hippocampal neurons. J. Comp. Neurol. 427, 351–361 (2000).
Campello, S. et al. Orchestration of lymphocyte chemotaxis by mitochondrial dynamics. J. Exp. Med. 203, 2879–2886 (2006).
Mills, K. M., Brocardo, M. G. & Henderson, B. R. APC binds the miro/milton motor complex to stimulate transport of mitochondria to the plasma membrane. Mol. Biol. Cell 27, 466–482 (2016).
Ferrante, A. W. Jr. The immune cells in adipose tissue. Diabetes Obes. Metab. 15 (Suppl. 3), 34–38 (2013).
Lumeng, C. N., Bodzin, J. L. & Saltiel, A. R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Invest. 117, 175–184 (2007).
Lumeng, C. N., Deyoung, S. M., Bodzin, J. L. & Saltiel, A. R. Increased inflammatory properties of adipose tissue macrophages recruited during diet-induced obesity. Diabetes 56, 16–23 (2007).
Guilherme, A., Virbasius, J. V., Puri, V. & Czech, M. P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 9, 367–377 (2008).
Kanneganti, T. D. & Dixit, V. D. Immunological complications of obesity. Nat. Immunol. 13, 707–712 (2012).
Acknowledgements
The authors apologize for being unable to cite papers that have contributed to the progress of this field owing to space limitations. The authors acknowledge support by the Canadian Institutes of Health Research (CIHR; grant PJT-159529) and Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant (to L.K.).
Author information
Authors and Affiliations
Contributions
Both authors contributed equally to all aspects of the article.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information
Nature Reviews Endocrinology thanks Y. Li, C. Scheele and M. Skelton for their contribution to the peer review of this work.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Glossary
- Creatine kinase–phosphocreatine circuit
-
Also known as the phosphocreatine shuttle, this system mediates the stoichiometric (1:1) transphosphorylation of phosphate from mitochondrial or glycolytic ATP to phosphocreatine, which is then used by creatine kinase to maintain high local ATP:ADP ratios.
- UCP1
-
A mitochondrial inner membrane protein that dissipates the proton gradient across the lipid bilayer, effectively decreasing the proton-motive force and minimizing ATP synthesis; the energy dissipated across the mitochondrial inner membrane results in a considerable increase in the rate of respiration, substrate oxidation and release of heat.
- Proton-motive force
-
The potential energy stored as a combination of the electrical and concentration (electrochemical) gradient across the mitochondrial inner membrane due to the extrusion of protons into the intermembrane space by the electron transport chain.
- Congenic background
-
An inbred strain of mouse where the control and experimental animals only differ from one another by a small genetic region (typically a single gene).
- Thermoneutrality
-
The ambient temperature where the metabolic rate is at a minimum, when temperature regulation is achieved by non-evaporative physical processes alone.
- Creatine-dependent thermogenesis
-
The phosphorylation of creatine by creatine kinase and subsequent dephosphorylation of phosphocreatine (or downstream phosphometabolite) that regenerates creatine and dissipates the high-energy phosphate to generate heat; also known as futile creatine cycling.
- P:O ratio
-
The number of moles of ADP phosphorylated to ATP for every two electrons that reduce oxygen to water.
- Thymocyte selection
-
During T cell differentiation, thymocytes can undergo expansion, differentiation (positive selection) or cell death (negative selection).
Rights and permissions
About this article

Cite this article
Kazak, L., Cohen, P. Creatine metabolism: energy homeostasis, immunity and cancer biology. Nat Rev Endocrinol 16, 421–436 (2020). https://doi.org/10.1038/s41574-020-0365-5
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41574-020-0365-5
This article is cited by
-
Metabolic heterogeneity in cancer
Nature Metabolism (2024)
-
Preoperative low plasma creatine kinase levels predict worse survival outcomes in bladder cancer after radical cystectomy
International Urology and Nephrology (2024)
-
Anti-adipogenesis effect of indole-3-acrylic acid on human preadipocytes and HFD-induced zebrafish
Acta Diabetologica (2024)
-
Association between serum creatinine and type 2 diabetes in the Chinese population: a retrospective cohort study
Scientific Reports (2023)
-
Depletion of creatine phosphagen energetics with a covalent creatine kinase inhibitor
Nature Chemical Biology (2023)
