
Abstract
Urinary tract infections are among the most common human bacterial infections and place a significant burden on healthcare systems due to associated morbidity, cost and antibiotic use. Despite being a facultative anaerobe, uropathogenic Escherichia coli, the primary cause of urinary tract infections, requires aerobic respiration to establish infection in the bladder. Here, by combining bacterial genetics with cell culture and murine models of infection, we demonstrate that the widely conserved respiratory quinol oxidase cytochrome bd is required for intracellular infection of urothelial cells. Through a series of genetic, biochemical and functional assays, we show that intracellular oxygen scavenging by cytochrome bd alters mitochondrial physiology by reducing the efficiency of mitochondrial respiration, stabilizing the hypoxia-inducible transcription factor HIF-1 and promoting a shift towards aerobic glycolysis. This bacterially induced rewiring of host metabolism antagonizes apoptosis, thereby protecting intracellular bacteria from urothelial cell exfoliation and preserving their replicative niche. These results reveal the metabolic basis for intracellular bacterial pathogenesis during urinary tract infection and identify subversion of mitochondrial metabolism as a bacterial strategy to facilitate persistence within the urinary tract.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Data availability
All data required for evaluation of these results are present in the text, figures or supplemental material. Transcriptional profiling data have been deposited in the Gene Expression Omnibus database (accession GSE188981) and are also contained in Supplementary Table 2a. Data from the following publicly available databases were used in this study: NCBI Protein Database (FASTA sequence: NP_415261.2) and the RCSB Protein Data Bank (PDB) (PDB IDs 6rko and 6rx4). Source data are provided with this paper.
References
Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 7, 653–660 (2010).
Klein, R. D. & Hultgren, S. J. Urinary tract infections: microbial pathogenesis, host–pathogen interactions and new treatment strategies. Nat. Rev. Microbiol. 18, 211–226 (2020).
Flores-Mireles, A. L., Walker, J. N., Caparon, M. & Hultgren, S. J. Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 13, 269–284 (2015).
Martinez, J. J., Mulvey, M. A., Schilling, J. D., Pinkner, J. S. & Hultgren, S. J. Type 1 pilus-mediated bacterial invasion of bladder epithelial cells. EMBO J. 19, 2803–2812 (2000).
Mulvey, M. A., Schilling, J. D. & Hultgren, S. J. Establishment of a persistent Escherichia coli reservoir during the acute phase of a bladder infection. Infect. Immun. 69, 4572–4579 (2001).
Anderson, G. G. et al. Intracellular bacterial biofilm-like pods in urinary tract infections. Science 301, 105–107 (2003).
Reigstad, C. S., Hultgren, S. J. & Gordon, J. I. Functional genomic studies of uropathogenic Escherichia coli and host urothelial cells when intracellular bacterial communities are assembled. J. Biol. Chem. 282, 21259–21267 (2007).
Conover, M. S. et al. Metabolic requirements of Escherichia coli in intracellular bacterial communities during urinary tract infection pathogenesis. mBio 7, e00104–e00116 (2016).
Hagan, E. C., Lloyd, A. L., Rasko, D. A., Faerber, G. J. & Mobley, H. L. Escherichia coli global gene expression in urine from women with urinary tract infection. PLoS Pathog. 6, e1001187 (2010).
Floyd, K. A. et al. The ubiI (VisC) aerobic ubiquinone synthase is required for expression of type 1 pili, biofilm formation, and pathogenesis in uropathogenic Escherichia coli. J. Bacteriol. 198, 2662–2672 (2016).
Eberly, A. R. et al. Biofilm formation by uropathogenic Escherichia coli Is favored under oxygen conditions that mimic the bladder environment. Int. J. Mol. Sci. https://doi.org/10.3390/ijms18102077 (2017).
Beebout, C. J. et al. Respiratory heterogeneity shapes biofilm formation and host colonization in uropathogenic Escherichia coli. MBio https://doi.org/10.1128/mBio.02400-18 (2019).
Beebout, C. J., Sominsky, L. A., Eberly, A. R., Van Horn, G. T. & Hadjifrangiskou, M. Cytochrome bd promotes Escherichia coli biofilm antibiotic tolerance by regulating accumulation of noxious chemicals. NPJ Biofilms Microbiomes 7, 35 (2021).
Borisov, V. B. & Verkhovsky, M. I. Oxygen as acceptor. EcoSal Plus 6, https://doi.org/10.1128/ecosalplus.ESP-0012-2015 (2015).
Shepherd, M. et al. The cytochrome bd-I respiratory oxidase augments survival of multidrug-resistant Escherichia coli during infection. Sci. Rep. 6, 35285 (2016).
Borisov, V. B., Gennis, R. B., Hemp, J. & Verkhovsky, M. I. The cytochrome bd respiratory oxygen reductases. Biochim. Biophys. Acta 1807, 1398–1413 (2011).
Mason, M. G. et al. Cytochrome bd confers nitric oxide resistance to Escherichia coli. Nat. Chem. Biol. 5, 94–96 (2009).
Giuffrè, A., Borisov, V. B., Arese, M., Sarti, P. & Forte, E. Cytochrome bd oxidase and bacterial tolerance to oxidative and nitrosative stress. Biochim. Biophys. Acta 1837, 1178–1187 (2014).
Hannan, T. J. et al. Host–pathogen checkpoints and population bottlenecks in persistent and intracellular uropathogenic Escherichia coli bladder infection. FEMS Microbiol. Rev. 36, 616–648 (2012).
Mulvey, M. A. et al. Induction and evasion of host defenses by type 1-piliated uropathogenic Escherichia coli. Science 282, 1494–1497 (1998).
Haugan, M. S., Charbon, G., Frimodt-Møller, N. & Løbner-Olesen, A. Chromosome replication as a measure of bacterial growth rate during Escherichia coli infection in the mouse peritonitis model. Sci. Rep. 8, 14961 (2018).
Rivera-Chavez, F. et al. Depletion of butyrate-producing Clostridia from the gut microbiota drives an aerobic luminal expansion of Salmonella. Cell Host Microbe 19, 443–454 (2016).
Mascolo, L. & Bald, D. Cytochrome bd in Mycobacterium tuberculosis: a respiratory chain protein involved in the defense against antibacterials. Prog. Biophys. Mol. Biol. 152, 55–63 (2020).
Cai, Y. et al. Host immunity increases Mycobacterium tuberculosis reliance on cytochrome bd oxidase. PLoS Pathog. 17, e1008911 (2021).
Mogi, T. et al. Probing the ubiquinol-binding site in cytochrome bd by site-directed mutagenesis. Biochemistry 45, 7924–7930 (2006).
Safarian, S. et al. Active site rearrangement and structural divergence in prokaryotic respiratory oxidases. Science 366, 100–104 (2019).
Lacerda Mariano, L. & Ingersoll, M. A. The immune response to infection in the bladder. Nat. Rev. Urol. 17, 439–458 (2020).
Friedrich, T. et al. Two binding sites of inhibitors in NADH: ubiquinone oxidoreductase (complex I). Relationship of one site with the ubiquinone-binding site of bacterial glucose:ubiquinone oxidoreductase. Eur. J. Biochem. 219, 691–698 (1994).
Degli Esposti, M. Inhibitors of NADH–ubiquinone reductase: an overview. Biochim. Biophys. Acta 1364, 222–235 (1998).
Tiku, V., Tan, M. W. & Dikic, I. Mitochondrial functions in infection and immunity. Trends Cell Biol. 30, 263–275 (2020).
Marchi, S., Morroni, G., Pinton, P. & Galluzzi, L. Control of host mitochondria by bacterial pathogens. Trends Microbiol. https://doi.org/10.1016/j.tim.2021.09.010 (2021).
Marsboom, G. et al. Dynamin-related protein 1-mediated mitochondrial mitotic fission permits hyperproliferation of vascular smooth muscle cells and offers a novel therapeutic target in pulmonary hypertension. Circ. Res. 110, 1484–1497 (2012).
Krab, K., Kempe, H. & Wikström, M. Explaining the enigmatic KM for oxygen in cytochrome c oxidase: a kinetic model. Biochim. Biophys. Acta 1807, 348–358 (2011).
Semenza, G. L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 3, 721–732 (2003).
Rathmell, J. C. et al. Akt-directed glucose metabolism can prevent Bax conformation change and promote growth factor-independent survival. Mol. Cell. Biol. 23, 7315–7328 (2003).
Klumpp, D. J. et al. Uropathogenic Escherichia coli induces extrinsic and intrinsic cascades to initiate urothelial apoptosis. Infect. Immun. 74, 5106–5113 (2006).
Klumpp, D. J. et al. Uropathogenic Escherichia coli potentiates type 1 pilus-induced apoptosis by suppressing NF-κB. Infect. Immun. 69, 6689–6695 (2001).
Lin, A. E. et al. Role of hypoxia inducible factor-1α (HIF-1α) in innate defense against uropathogenic Escherichia coli infection. PLoS Pathog. 11, e1004818 (2015).
Alteri, C. J., Smith, S. N. & Mobley, H. L. Fitness of Escherichia coli during urinary tract infection requires gluconeogenesis and the TCA cycle. PLoS Pathog. 5, e1000448 (2009).
Alteri, C. J. & Mobley, H. L. Escherichia coli physiology and metabolism dictates adaptation to diverse host microenvironments. Curr. Opin. Microbiol. 15, 3–9 (2012).
Murphy, K. C. & Campellone, K. G. Lambda Red-mediated recombinogenic engineering of enterohemorrhagic and enteropathogenic. E. coli. BMC Mol. Biol. 4, 1–12 (2003).
Hung, C. S., Dodson, K. W. & Hultgren, S. J. A murine model of urinary tract infection. Nat. Protoc. 4, 1230–1243 (2009).
Hartmann, R. et al. Quantitative image analysis of microbial communities with BiofilmQ. Nat. Microbiol. 6, 151–156 (2021).
Pfaffl, M. W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 29, e45 (2001).
Leaver-Fay, A. et al. ROSETTA3: an object-oriented software suite for the simulation and design of macromolecules. Methods Enzymol. 487, 545–574 (2011).
Song, Y. et al. High-resolution comparative modeling with RosettaCM. Structure 21, 1735–1742 (2013).
Alford, R. F. et al. An integrated framework advancing membrane protein modeling and design. PLoS Comput. Biol. 11, e1004398 (2015).
Koehler Leman, J., Mueller, B. K. & Gray, J. J. Expanding the toolkit for membrane protein modeling in Rosetta. Bioinformatics 33, 754–756 (2017).
Ashkenazy, H. et al. ConSurf 2016: an improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 44, W344–W350 (2016).
Landau, M. et al. ConSurf 2005: the projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Res. 33, W299–W302 (2005).
Glaser, F. et al. ConSurf: identification of functional regions in proteins by surface-mapping of phylogenetic information. Bioinformatics 19, 163–164 (2003).
Park, H. et al. Simultaneous optimization of biomolecular energy functions on features from small molecules and macromolecules. J. Chem. Theory Comput. 12, 6201–6212 (2016).
Frenz, B. et al. Prediction of protein mutational free energy: benchmark and sampling improvements increase classification accuracy. Front. Bioeng. Biotechnol. 8, 558247 (2020).
Conway, P., Tyka, M. D., DiMaio, F., Konerding, D. E. & Baker, D. Relaxation of backbone bond geometry improves protein energy landscape modeling. Protein Sci. 23, 47–55 (2014).
Khatib, F. et al. Algorithm discovery by protein folding game players. Proc. Natl Acad. Sci. USA 108, 18949–18953 (2011).
Nivon, L. G., Moretti, R. & Baker, D. A Pareto-optimal refinement method for protein design scaffolds. PLoS ONE 8, e59004 (2013).
Tyka, M. D. et al. Alternate states of proteins revealed by detailed energy landscape mapping. J. Mol. Biol. 405, 607–618 (2011).
Dwyer, D. J. et al. Antibiotics induce redox-related physiological alterations as part of their lethality. Proc. Natl Acad. Sci. USA 111, E2100–E2109 (2014).
Rasmussen, M. L. et al. MCL-1 inhibition by selective BH3 mimetics disrupts mitochondrial dynamics causing loss of viability and functionality of human cardiomyocytes. iScience 23, 101015 (2020).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Acknowledgements
We thank K. Voss and H. Woods for technical assistance and helpful advice. This work was supported by National Institutes of Health (NIH) grants F30AI150077 (C.J.B.), T32GM007347 (C.J.B. and B.I.R.), F99NS125829 (G.L.R.), F30CA247202 (B.I.R.), F32CA250258 (A.M.B.), T32AI112541 (G.H.M.), T32GM007569 (J.R.B.), R01AI127793 (W.J.C.), R01AI101171 (W.J.C.), R01DK105550 (J.C.R.), R35GM128915 (V.G.), RF1MH123971 (V.G.), R01AI107052 (M.H.), P20DK123967 (M.H.) and the Howard Hughes Medical Institute Gilliam Fellowship (G.L.R.). Transmission electron microscopy was performed by W. Beatty at the Molecular Microbiology Imaging Facility (Washington University in St. Louis). Confocal laser scanning and structured illumination microscopy were performed at the Vanderbilt Cell Imaging Shared Resource, which is supported by NIH grant DK20593. NanoString analysis was performed at the Vanderbilt Technologies for Advanced Genomics core facility, which is supported by NIH grants UL1RR024975, P30CA68485, P30EY08126 and G20RR030956. Access to the Vanderbilt Advanced Computing Center for Research and Education was supported in part by NIH grants S10RR031634 and S10OD023680. Some images were created using BioRender.com.
Author information
Authors and Affiliations
Contributions
C.J.B. conceived the study, performed most experiments and composed the manuscript. G.L.R. acquired and analysed mitochondrial imaging data. B.I.R. aided in the design, acquisition and interpretation of flow cytometry data. A.M.B. performed structural modelling experiments and HIF-1α immunoblots. G.H.M. performed computational analysis of imaging data. J.R.B. performed mouse infection experiments. W.J.C., W.K.R., J.C.R. and V.G. contributed essential resources and aided in the design and interpretation of experiments. M.H. conceived the study and oversaw all aspects of its execution. All authors contributed to the generation, analysis or interpretation of the data and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflicts of interest at the time of submission of this manuscript.
Peer review
Peer review information
Nature Microbiology thanks Mark Schembri and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Cytochrome bd does not influence type 1 pilus dependent adherence or invasion of urothelial cells.
a, Anti-FimA immunoblot performed on normalized samples boiled in acidified SDS to depolymerize type 1 pili. b, Anti-FimA immunoblot performed on normalized samples with polymerized type 1 pili. Each band represents FimA polymers of a different size. c, Bacterial hemagglutination titer with or without the FimH inhibitor mannose; mean ± SEM; one-way ANOVA with Dunnett’s test for multiple comparisons. d, Total, adherent, and intracellular bacterial titers in infected urothelial cells; geometric mean ± 95% CI; Kruskal-Wallis test with Dunn’s test for multiple comparisons. All experiments were performed with a minimum of three biological replicates. Each point represents a biological replicate. Exact p-values are provided in the figure, with bold values indicating statistical significance (p < 0.05).
Extended Data Fig. 2 Loss of cytochrome bd increases intracellular bacterial sensitivity to nitrofurantoin.
Percent survival of intracellular UPEC treated with ciprofloxacin or nitrofurantoin as compared to vehicle treated controls. Data is representative of six biological replicates per group. Each point represents a biological replicate; mean ± SEM; two-tailed unpaired t test. Exact p-values are provided in the figure, with bold values indicating statistical significance (p < 0.05).
Extended Data Fig. 3 Loss of cytochrome bd does not impair flagellar biosynthesis.
a, Representative images of wild-type and ∆cydAB transformed with a transcriptional reporter for FlhDC, the master regulator of flagellar expression. b, GFP intensity normalized to cell area; median ± 95% CI; two-tailed unpaired t test. c, Representative transmission electron microscopy images of bacterial cells. Arrows indicate flagella. Images are representative of three biological replicates. Exact p-values are provided in the figure, with bold values indicating statistical significance (p < 0.05).
Extended Data Fig. 4 Urothelial cells upregulated NOS2 in response to intracellular infection.
Normalized counts of NOS2 (inducible nitric oxide synthase, iNOS) transcript in intracellularly infected urothelial cells. Data is representative of at least three biological replicates per group; mean ± SEM; one-way ANOVA with Tukey’s test for multiple comparisons. Exact p-values are provided in the figure, with bold values indicating statistical significance (p < 0.05).
Extended Data Fig. 5 UPEC is not sensitive to rotenone or antimycin A.
OCR readings of wild-type UPEC treated with vehicle (open circles) or treated with oligomycin, FCCP, and rotenone/antimycin A (closed circles) presented as percent OCR of time = 0; mean ± SEM. Data was fit to a one phase decay model (R2 > 0.9 for both groups) and statistically analyzed by comparing k (p = 0.6402). Data is representative of five biological replicates, each with at least three technical replicates.
Extended Data Fig. 6 Expression of respiratory oxidases in intracellular bacterial populations.
a, Abundance of respiratory oxidase transcripts in the inoculum used for infections and intracellular bacterial populations compared to gyrB. Dotted line indicates gyrB abundance; mean ± SEM; two-tailed unpaired t test. b, Relative abundance of respiratory oxidase transcript in the inoculum and intracellular populations. c, Representative peptide nucleic acid in situ hybridization (PNA-FISH) image of IBCs. All experiments were performed with a minimum of three biological replicates. Each point represents a biological replicate. Exact p-values are provided in the figure, with bold values indicating statistical significance (p < 0.05).
Extended Data Fig. 7 Intracellular infection modulates urothelial cell transcriptional programs.
a, Volcano plot depicting changes in transcript abundance between wild-type and mock infected urothelial cells. Transcripts involved in glycolysis, glucose uptake, and hypoxia are denoted by color. b, Volcano plot depicting changes in the abundance of metabolic transcripts depicted in Fig. 5e between wild-type and mock infected urothelial cells. Data is representative of at least three biological replicates per group. Transcript normalization, differential expression calculations, and statistical comparisons were performed on the nSolver Advanced Analysis platform using the Benjamini-Yekutieli method.
Extended Data Fig. 8 Cytochrome bd has minimal impact on urothelial cell immune response to intracellular infection.
Directed pathway expression score for pathways not directly involved in central metabolism in wild-type and ∆cydAB infected compared to mock infected urothelial cells; mean ± SEM; two-tailed unpaired t test. Data is representative of at least three biological replicates per group. Exact p-values are provided in the figure, with bold values indicating statistical significance (p < 0.05).
Extended Data Fig. 9 Flow cytometry gating strategy.
Single cells were selected by gating on FSC-A and FSC-H. Debris and bacteria were subsequently excluded by gating out the FSC-Alow, SSC-Alow population. The remaining population was analyzed for UPEC (GFP) and annexin V (Pacific Blue) or caspase-3 activity (Pacific Blue). Because we are interested in quantifying cell death, dead cells were not specifically excluded from analyses. Each dot represents one cell. Relative population density is represented by a color spectrum (red indicates regions of high cell density, blue indicates regions of low cell density).
Extended Data Fig. 10 Proposed model.
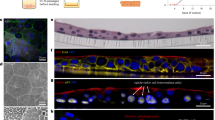
Schematic depicting the proposed model of how intracellular infection modulates urothelial cell metabolism and survival. left, During bladder infection UPEC induces a strong inflammatory response that triggers urothelial cell apoptosis and exfoliation. Urothelial cell exfoliation exposes underlying tissue layers to infection and promotes bacterial persistence in the bladder. right, By consuming oxygen and activating HIF-1 signaling, intracellular bacterial aerobic respiration alters urothelial cell metabolism and antagonizes apoptosis, allowing UPEC to complete its intracellular infection cascade and evade exfoliation.
Supplementary information
Source data
Source Data Fig. 1
Source data.
Source Data Fig. 2
Source data.
Source Data Fig. 3
Source data.
Source Data Fig. 4
Source data.
Source Data Fig. 5
Source data.
Source Data Fig. 5
Unprocessed immunoblots for Fig. 5.
Source Data Fig. 6
Source data.
Source Data Extended Data Fig. 1
Source data.
Source Data Extended Data Fig. 1
Unprocessed immunoblots for Extended Data Fig. 1.
Source Data Extended Data Fig. 2
Source data.
Source Data Extended Data Fig. 3
Source data.
Source Data Extended Data Fig. 4
Source data.
Source Data Extended Data Fig. 5
Source data.
Source Data Extended Data Fig. 6
Source data.
Source Data Extended Data Fig. 7
Source data.
Source Data Extended Data Fig. 8
Source data.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Beebout, C.J., Robertson, G.L., Reinfeld, B.I. et al. Uropathogenic Escherichia coli subverts mitochondrial metabolism to enable intracellular bacterial pathogenesis in urinary tract infection. Nat Microbiol 7, 1348–1360 (2022). https://doi.org/10.1038/s41564-022-01205-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-022-01205-w
This article is cited by
-
UPEC-mediated alteration of host mitochondrial function — a survival strategy
Nature Reviews Urology (2022)
